dominancia hemisfÉrica
DESCRIPTION
dominancia hemisfericaTRANSCRIPT

1
CAP 14 DE RAPIN DESTREZA MANUAL: UNA PERSPECTIVA BIOLÓGICA. INTRODUCCIÓN La tendencia hacia la destreza manual diestra ha sido foco de gran interés desde el comienzo de la NPS. Como el lenguaje, otra de las funciones más lateralizadas de nuestro cerebro, se piensa que la cantidad de humanos diestros representa algo único en nuestra especie. Por lo tanto el estudio de los orígenes y la significación de la destreza manual puede proveer una idea fundamental dentro de lo innato en el humano. Igualmente importante es el desarrollo en entender el origen de la variación individual de la destreza manual. Además 2 temas están muy relacionados. Una posible significación adaptativa de la destreza manual diestra o su correlato fundamental debe estar considerada en el contexto de la variabilidad fenotípica en los actos de selección natural. En este capítulo tendemos a proveer una explicación de la destreza diestra desde una perspectiva biológica, un enfoque que creemos es el más apto para ayudarnos a entender qué hace el cerebro humano. Una perspectiva biológica puede justificarse por las observaciones en que la variación de destreza manual está probablemente influenciada por factores genéticos, que el fenómeno de ser diestro es bastante más invariable a lo largo de las culturas y generaciones y que la asimetría manual aparece después del nacimiento. Debería destacarse que la propuesta biológica no excluye el análisis sistemático de las influencias ambientales, en la misma forma que el análisis biológico del desarrollo del lenguaje necesita especificar la naturaleza y el tiempo de importantes influencias ambientales. Hay 3 importantes aspectos del esquema biológico que son propuestos aquí. El 1ro que proponemos es de una perspectiva ontogénica. El desarrollo neural es un campo de juego en el cual interactúan lo innato y lo adquirido. Una perspectiva del desarrollo de dinámicas temporales de las variaciones fenotípicas pueden también restringir nuestras teorías de las influencias causales sobre la variación. 2do intentamos entender los mecanismos por la cual la variación individual en la preferencia de una mano y habilidad son inmediatas. Esto es, ¿cuál es el sustrato neural para las diferencias en destreza manual y cómo hacen para emerger? Finalmente investigamos las argumentaciones básicas de evolución genética sobre destreza manual. : Esto trae consigo 2 clases de preguntas, acerca de los problemas de filogenia y los factores de selección conservando la diversidad genética actual. La literatura científica actual sobre destreza manual es inmensa, reflejando un interés duradero y de fácil medida. Son necesarias unas pocas palabras sobre la evaluación de la destreza manual. Hay 2 importantes expresiones de destreza manual para considerar, ej la mano preferida usada para varias tareas, y la diferencia relativa en la habilidad manual para obtener tareas. Las correlaciones entre mediciones de la asimetría en ejecución y preferencia son generalmente significativas, aunque la magnitud de la interrelación varía a lo largo de las tareas y está disminuida cuando los zurdos y diestros son examinados separadamente. Los inventaros de preferencia son los más comunes para intentar evaluar la destreza manual. Es conocido generalmente que la preferencia de escribir con una mano está influenciada más por presión social que por otras actividades. Hay diferentes tipos de tareas usadas para la evaluación relacionada con la mano hábil con importantes diferencias en la complejidad de las tareas. Las mediciones de ejecución están distribuidas con muchos menos sesgos que las mediciones de preferencia (y pueden estar normalmente distribuidas) debido a que los individuos que prefieren consistentemente su mano derecha varían considerablemente con la ejecución de la mano. Por lo tanto las medidas de ejecución hacen un trabajo mejor en los diferentes extremos desde una manualidad derecha moderada. La selección de una medida de ejecución versus una de preferencia depende de asuntos prácticos y teóricos. La fácil administración de los inventarios de preferencia hace que sea la medición más común en estudios a gran escala. Ellos son más apropiados para teorías de categorización fenotípica, y menos para teorías que enfatizan continuamente la distribución fenotípica. En general, los inventarios de preferencia son menos satisfactorios que las mediciones de ejecución en distinguir la dirección del grado de destreza, un tema potencialmente importante en análisis genético. LO INNATO Y LA ONTOGENIA DEL FENOTIPO 3 tipos diferentes de asimetría han sido descriptas. La ASIMETRÍA DIRECCIONAL existe a nivel de la población debido a un “diseño” orgánico de asimetría. Un ej de asimetría direccional es el del diseño asimétrico del

2
corazón humano. La ANTISIMETRÍA se refiere a la situación donde los individuos tienen diferentes diseños para asimetría, con alguna tendencia del lado izquierdo de rasgos más amplios que en el derecho, en otros lo opuesto. La ANTISIMETRÍA puede resultar en una distribución bimodal. La ASIMETRÍA FLUCTUANTE (AF) se refiere a la desviación de la simetría de un rasgo, y no debida al diseño desarrollado para la asimetría. La AF refleja la carencia de un control preciso sobre el desarrollo de un rasgo en el sentido de potenciales perturbaciones como mutaciones o agentes patógenos, resultando en una salida a partir del diseño del desarrollo para simetría. Por ej. Escuchar del lado derecho e izquierdo tiende a ser simétrico en los humanos. El factor que algunos individuos tengan una escucha más amplia en su lado derecho y algunos en sus oídos izquierdos es debido a la expresión imprecisa del diseño desarrollado, no a causa de algún diseño genético individual para escucha asimétrica. En contraste, el factor que mayor cantidad de gente tiende a estar capacitada con la mano derecha que con la izquierda, una asimetría direccional, es probablemente un resultado de diseño, no de error desarrollado. Palmer condujo un análisis filogenético en el sentido de asimetrías direccionales desarrolladas o evolucionadas y encontró que la asimetría direccional tiende desarrollarse desde cambios en el estado larval de una clase simétrica ancestral. Mientras no se excluya un rol de las influencias ambientales tardías estos patrones sugieren que, más frecuentemente que no, son importantes factores citogénicos internos para el desarrollo de la asimetría funcional. Consistente con el análisis de Palmer, muchas evidencias directas e indirectas indican que las asimetrías asociadas con la destreza manual surgen tempranamente en el desarrollo. Las evidencias directas sugieren que la asimetría motora puede ser observada tempranamente en el desarrollo fetal. Las evidencias indirectas vinculan la variación en destreza manual con variaciones en otros rasgos físicos determinados tempranamente en el desarrollo. Sin embargo, como pocos estudios proveen una perspectiva longitudinal limitan las conclusiones con respecto a la estabilidad temporal de expresiones de la destreza manual. Los estudios de ultrasonido de fetos humanos revelan la asimetría marcada de destreza manual. En un estudio de 274 fetos fueron observados succionando sus dedos pulgares y el 92% demostró succión de su pulgar derecho. Aquellos entre 12 y 15 semanas de edad gestacional el 87% succionaban su pulgar derecho, demostrando la emergencia temprana de la preferencia lateral. En un estudio de 72 fetos observados a las 10 semanas de edad gestacional mostraban mayor frecuencia de movimientos en su brazo derecho que en el izquierdo, mientras que el 12% mostraba mayor frecuencia de movimientos en su brazo izquierdo y el otro 12% un uso de igual frecuencia de ambos brazos. También se destacó una disminución en la frecuencia de movimientos de brazos tanto del derecho como del izquierdo en las semanas 21 y 27 de gestación. No se sabe si éstas asimetrías tempranas predicen la subsiguiente destreza manual en ñ o adultos jóvenes, la succión del pulgar derecho está asociada con el giro cefálico del lado derecho justo después del nacimiento el cual predice la preferencia manual hacia los 18 meses. Los índices de crecimiento fetal parecen estar relacionados con la destreza manual del adulto. Una forma indirecta de investigar el índice de crecimiento fetal es a través de la medición de los DERMATOGLIFOS.La génesis de las características dermatoglíficas ocurre en la 6ta semana de gestación y el patrón final se establece en la semana 19. Muchos estudios han encontrado que los diestros y zurdos muestran diferentes patrones de características dermatoglíficas, idea que la magnitud de los efectos son bastantes más bajas. En uno de estos grandes estudios, Rife informó que “la asimetría bilateral individual es la más grande si bien la asimetría grupal es menor entre zurdos que en diestros”. Esto es, que la mano derecha e izquierda de un zurdo fueron muy diferentes entre ellas que las 2 manos de un diestro, pero las asimetrías direccionales fueron evidentes en diestros. Como las variaciones en los patrones dérmicos reflejan variaciones en los índices de crecimiento. El informe de Rife sugiere una mayor variación lateral en índices de crecimiento en zurdos que en diestros. Coren destacó que las características del patrón dermatoglífico en las impresiones digitales de los zurdos parecía un patrón simplificado de los individuos con Down. Como el síndrome de Down es asociado con un índice de crecimiento prenatal mas lento, las observaciones de Coren dieron origen a la posibilidad que los individuos zurdos in útero deben también estar vinculados con índices de crecimiento lento. Informes de un gran n|° de ANORMALIDADES FÍSICAS MENORES (MPAs) también pueden indicar una asociación de destreza manual con variaciones en los índices tempranos de crecimiento. Las MPAs están más comúnmente evaluadas con la escala de HALVERSON Y WALDROP, quienes evalúan varias características de la cara, cabeza, manos y cuerpo que reflejan una perturbación o enlentecimiento del desarrollo prenatal, que ocurre típicamente en el 1er y principios del 2do trimestre. Un ej es el hipertelorismo (mayor distancia entre ambos ojos). En un momento particular del desarrollo prenatal los ojos migran hacia al línea media; si el desarrollo en ese momento está enlentecido, los ojos permanecen muy distanciados. En cerebros humanos, la asimetría del planum temporale se establece alrededor de las 15 semanas de edad gestacional y una asimetría atípica del planum es observada frecuentemente en adultos zurdos. Estas vinculaciones entre la destreza

3
manual y las características físicas determinadas durante la gestación llaman la atención sobre la importancia de las influencias prenatales en el fenotipo de la destreza manual en el adulto. El desarrollo de la destreza manual en los ñ y chicos jóvenes ha estado sujeta a una gran relación con la investigación. Muchos estudios proveen evidencias de algún tipo de inclinación dirigida desde la infancia temprana, pero la emergencia de una preferencia manual relativamente estable a los 3 años de edad o si es precedida por patrones algunas veces fluctuantes y complejos del desarrollo. Michel describe en aproximadamente, a lo largo del 1er año de vida, 50% de los ñ la preferencia en alcanzar algo con la mano derecha , con 25% de ñ con la mano izquierda o sin preferencia. Un leve aumento de la preferencia manual derecha para manipular es observado durante el mismo período de tiempo. En contraste, la manipulación bimanual de los objetos en el cual la mano preferida “explora” y la mano no preferida “sostiene”, es típicamente vista la preferencia no lateral hasta el final del 1er año. El 46% de los ñ mostraron una preferencia estable a lo a largo del tiempo de la mano derecha para alcanzar y manipular objetos a los 7, 9, 11 y 13 meses de edad, mientras que el 18% mostró una preferencia izquierda estable y el 36% no exhibió preferencia anual estable. Si bien la inclinación de la mano derecha es claramente demostrada para alcanzar, estos estudios demostraron menor lateralidad notable que los estudios prenatales de Hepper, en el cual la proporción de destreza manual derecha se aproxima más a los adultos. Un reciente estudio de Corbetta demuestra los cambios en el desarrollode destreza manual en ñ en forma individual revelando las notables fluctuaciones en asimetría. Estudiaron 4 ñ semanalmente durante el 1er año de vida con técnicas cinemáticas sofisticadas, dando mediciones de preferencia en movimientos de alcanzar y no alcanzar. Se destacó la notable variabilidad temporal, si bien global, donde cada ñ mostró algún grado de preferencia derecha. Luego, a los 3 ½ de edad, se encontró una preferencia manual derecha bastante consistente para el uso de objetos. Los autores hablan a favor de una interpretación de “sistemas dinámicos” en estos informes, manteniendo que “ en el desarrollo cambia la lateralidad, pudiendo estar influenciada por cambios en la postura como ñ aprenden a sentarse, gatear y caminar”. La emergencia de estas otras habilidades motoras fuerzan la reorganización de habilidades lateralizadas más de una vez en el 1er año de vida. El grado de superioridad observado en la mano derecha en las tareas manuales como movimientos de separar parecen ser relativamente estables después de los 3 años. Por lo tanto, a pesar del uso de una mano preferida por años, los adultos y ñ grandes muestran el mismo grado de asimetría de ejecución. La mano preferida, sin embargo, parece mostrar un curso de desarrollo diferente. Harris revió diferentes estudios demostrando que la incidencia de destreza manual “ambigua” disminuía durante la infancia, en asociación con un aumento de la preferencia diestra. A pesar de este aparente origen prenatal temprano, la manera compleja en la cual el fenotipo de la destreza manual se desarrolla es en efecto probablemente el mejor rasgo de un proceso dinámico. En este sentido puede ser distinguido desde otros rasgos físicos influenciados por procesos tempranos del desarrollo , tales como los dermatoglifos y las MPAs. En contraste éstos “restos fósiles de eventos prenatales”, la destreza manual es un fenotipo evolucionado, al menos durante los 1ros de la infancia. Nuestro desafío es entender en qué momentos la genética y las influencias ambientales afectan aspectos específicos de estos procesos dinámicos. Por lo tanto hablamos del fenotipo de la destreza manual como teniendo 2 dimensiones correlacionadas, ej asimetría en la ejecución y en la preferencia. Hay muchas evidencias, sin embargo, que en adultos el fenotipo puede ser más complejo. Peters demostró que diferentes patrones de preferencia surgen en zurdos consistentes (con una preferencia marcada y estable de la mano izquierda en las tareas) versus los zurdos inconsistentes (que prefieren su mano izquierda para algunas tareas y la derecha para otras). Mientras ambos grupos escriben con su mano izquierda y demuestran gran habilidad en su mano izquierda en el Test de Purdue Pegboard, el zurdo inconsistente es más sólido en su mano derecha, prefiere arrojar con su mano derecha (y no es sorprendente, arrojan mejor con ella). Por lo tanto, asimetrías para habilidades que requieren un control de la motricidad fina parecen estar disociadas de aquellas que implican movimientos de fuerza y del brazo entero. Los variados estudios cruzados en adultos han demostrado un aumento de la preferencia de la mano derecha con el avance de la edad. Esta tendencia puede reflejar en parte un efecto cohorte más que un efecto de la edad, debido en los recientes años a una presión social disminuida contra los zurdos. Lo que no es claro, sin embargo, es si bien éste es el único involucrado. Mc Manus ha sugerido que la frecuencia genética actual puede estar cambiando, tal que los alelos que favorecen la destreza manual izquierda estando siendo más comunes. Raramente, algunos individuos cambian de una destreza izquierda hacia una derecha con el avance de la edad, si bien este efecto no explica la incidencia reducida de zurdos con la edad. Ellis intento discriminar los temas de preferencia manual que son más aptos para estar sujetos a la

4
presión social (mano que escribe) desde aquellos menores ( ej arrojar) y encontró que a nivel de la eliminación de la anterior, el aumento edad relacionada en la preferencia manual derecha fueron a pesar de todo observado. MECANISMOS CEREBRALES SUBYACENTES A LA ASIMETRÍA PREFERENCIA Y HABILIDAD MANUAL. En esta sección hablamos de evidencias con respecto al sustrato neural de la destreza manual. Numerosas asimetrías anatómicas están correlacionadas con variaciones en la destreza manual, si bien solo algunos de estos probablemente representan sus sustratos anatómicos. Algunas asimetrías vinculadas con la destreza manual, en virtud de su localización anatómica , están improbablemente relacionadas directamente a las habilidades motoras. Más notable acerca de estas asimetrías es el planum temporal y la petalia frontal y occipital. Beaton recientemente hizo un análisis crítico de estudios relacionando la variación de asimetrías a la habilidad manual. Él concluyó “que parece que de la asimetría de la destreza manual y del plano temporal están relacionadas en alguna forma, pero la exacta naturaleza de esta relación es oscura”. Luego, los resultados a través de estudios “el origen que posibilita que alguna diferencia en la magnitud de la asimetría del planum está relacionada no mucho con la direccionalidad como con el grado de destreza manual.” Le May llamó la atención de la tendencia de los seres humanos para tener una mayor amplitud y longitud en la corteza anterior derecha y una mayor amplitud y longitud en la corteza posterior izquierda (petalia). Estos patrones parecen estar atenuados o revertidos en los zurdos. Bear encontró que los zurdos tienen una asimetría posterior atípica pero anterior normal, mientras otros estudios no observaron diferencias en la destreza manual. El mismo patrón de asimetría destacada para la petalia cortical ha sido identificado en el cerebelo, y éstos parecen estar más atenuados en los zurdos. Sin embargo, mientras las proyecciones cerebelosas homolaterales, en contraste a la corteza, el hecho que las mismas asimetrías son destacadas en diferentes niveles del neuroeje plantea la cuestión acerca de si las asimetrías cerebelares son parte del sustrato anatómico de la asimetría en habilidad manual o preferencia. Quizás, una gran incidencia del plegamiento cortical y cerebelar atípico en zurdos refleja el hecho que los zurdos son más aptos para demostrar anormalidades mínimas neurales análogas a la gran incidencia demostrada en los MPAs. Diferencias entre los zurdos y diestros en la anatomía del cuerpo calloso probablemente no está relacionada en forma directa con las habilidades motoras. Un estudio reciente encontró una mayor área callosa en los hombres zurdos con supuesta dominancia de lenguaje en el hemisferio derecho que en los hombres zurdos o diestros con una supuesta dominancia de lenguaje en el hemisferio izquierdo. Variaciones en la corteza sensoriomotriz y sus conexiones son probablemente de gran relevancia en la variación individual en la destreza manual. Regiones diferentes pueden contribuir a distintos aspectos del fenotipo. La destreza manual es evidente en varios aspectos de la habilidad motora, incluyendo el control motor fino de los movimientos independientes de los dedos, control para manipulear, tomar, arrojar o embocar objetos. Cada una de estas funciones está controlada por aspectos diferentes del sistema motor, con movimientos de los dedos más distales controlados primariamente por los haces corticoespinales originados en el área 4 de Brodman. Sin embargo los movimientos que involucran el brazo y el tronco reciben control indirecto cortical a través de conexiones occipito-temporales y frontales a los núcleos subcorticales de los sistemas ventromedial y lateral. Por lo tanto el grado por el cual las asimetrías corticales subyacen a la destreza manual dependen de la función específica tratada. En base a estudios de lesiones, Jakobson argumentó que movimientos de alcanzar y tomar guiados visualmente requieren información que debe experimentar un procesamiento dentro de la amplia corteza dorsal de las proyecciones visuales. La integración inicial tiene lugar en la corteza parietal posterior y luego en las áreas corticales frontales. Las asimetrías identificadas en el sistema motor relevante a la destreza manual probablemente representa solo parte de regiones diversas involucradas en los movimientos de la mano y brazo. Las funciones asimétricas relacionadas a la destreza manual han sido documentadas tanto para las áreas motoras subcorticales como las vías de control motor distal y espinal. El trayecto cortico-espinal es el haz más largo de fibras descendentes desde el cerebro. A nivel medular la mayor parte de las fibras cruzan la línea media y descienden en la columna ventral contralateral del cordón espinal. Alrededor del 10-15 % de estas fibras son homolaterales. Aunque la asimetría notable de estos está documentada no tiene relación con la destreza manual. Sin embargo, asimetrías morfológicas pueden existir

5
en el cordón espinal. En segmentos del cordón espinal que inervan brazos y mano, una gran motoneurona fue observada en el lado derecho, quizás representando un precursor a nivel celular de la destreza manual. Los estudios de flujo sanguíneo funcional han examinado la activación contralateral y homolateral durante el control de la motricidad fina. Por ej movimientos repetitivos de los dedos, el giro pre y post central la corteza motora lateral, el área cortical suplementaria, activaciones ocurridas 1riamnete en el hemisferio contralateral al movimiento. Grafton mostró una mayor activación de la corteza occipital dorsal que la activación temporal o ventral durante la acción de “señalar o asir”. El opérculo parietal izquierdo se activa durante la acción de asir pero no en la de señalar con la mano contralateral. Estos resultados demuestran que vastas redes de regiones cerebrales están involucradas en un grupo de tareas por las cuales son observadas la preferencia y habilidad manual. Los estudios en neuroimagen funcional son más consistentes que los estudios anatómicos con respecto a la posible asimetría relacionada a la destreza manual. Los estudios de RMNf comparan los zurdos con los diestros en tareas secuenciales de tocarse el pulgar con los otros dedos. La activación de la corteza motora derecha fue vista en movimientos de los dedos de la mano izquierda tanto en diestros y zurdos. La activación de la corteza motora izquierda con movimientos de los dedos homolaterales y contralaterales, si bien el grado de activación homolateral fue mayor en diestros. Otros recientes estudios de RMNf también demostraron la mayor activación en el córtex 1rio contralateral de la mano dominante que la mano no dominante. Luego, el grado de asimetría en la activación de la corteza motora fue correlacionada con el grado de preferencia manual (determinado por el Inventario de Edimburgo) en un grupo de 7 diestros y 6 zurdos. La asimetría en tamaño de las representaciones motoras izquierdas y derecha en sujetos diestros y zurdos estaba muy relacionada con el grado de destreza manual. ESTABLECIMIENTO EMBRIÓNICO DE LA ASIMETRÍA Los sustratos anatómicos de la destreza manual que surgen desde la distribución en redes con componentes asimétricos más notablemente involucra la corteza parieto-frontal. Las asimetrías motoras surgen en un punto del desarrollo antes que el cerebro puede controlar la actividad motora. De hecho, la corteza motora humana está en un rudimentario estadío del desarrollo a los 5 meses de edad gestacional. Por lo tanto el control neural periférico es mucho más probable ser el responsable para asimetrías motoras que las asimetrías subcortical y cortical. La actividad motora prenatal representa probablemente movimientos “autogénicos” o autogeneradores involucrando tanto neuronas excitatorias como inhibitorias. Las conductas autogénicas constituyen una característica primaria y fundamental de los estadios tempranos de la ontogenia. Muchos de los complejos patrones de acción motora especie-típica que caracterizan el repertorio conductual de los animales adulto (nadar, volar, locomoción, etc.) muy probablemente representan la activación del circuito neuronal cuya larga extensión tiene sus orígenes en la sustrato neurobiológico básico impuesto en el embrión y en quien la manifestación funcional se refleja en la conducta prenatal espontánea. Las fibras de los ganglios basales y talámicas desde la cápsula interna penetran en la corteza desarrollada en el 2do o 3er mes de edad gestacional y el desarrollo de la lámina cortical normal parece depender de la disponibilidad de los inputs aferentes a la corteza motora. La importancia de la actividad aferente del desarrollo característico del sistema visual es bien conocida. Por ej las ondas rítmicas de la actividad de barrido a través de la retina hacen surgir un input ordenado al núcleo geniculado lateral; las ondas desde las 2 retinas no están relacionadas, Basados en le principio que “neuronas que disparan juntas, se contactan juntas” este tipo particular de actividad ayuda a asegurar tanto la organización retinotópica y la separación neural de los inputs corticales y talámicos desde cado ojo. En una forma similar, los movimientos prenatales de las extremidades superiores representan una “actividad ordenada”, a diferencia del ruido, y el input asimétrico al tálamo debería tener consecuencias en el desarrollo. Quizás la actividad asimétrica de las extremidades superiores en los embriones conducen a la asimetría del desarrollo cortical y talámica, contribuyendo últimamente a la asimetría descripta en la sección previa. ¿Qué produce esta actividad asimétrica periférica? Los mejores candidatos hasta el presente son moléculas expresadas asimétricamente que regulan las proliferación, adhesión y migración celular en el desarrollo embriónico temprano. Si bien es desconocida la manera en la cual la definición del eje derecha-izquierda está determinado en el desarrollo embrionico temprano, una cascada de estudios recientes proveen evidencias que muchas de las diferentes moléculas están expresadas asimétricamente, en diferentes lugares embrionicos y en diferentes estadios del desarrollo. INFLUENCIAS PRENATALES EN LA VARIACIÓN INDIVIDUAL EN LA DESTREZA MANUAL

6
Un gran número de factores afectan potencialmente el desarrollo del SNC. Esto incluye infecciones, stress y variaciones hormonales. De éstas las variaciones hormonales han sido claramente el tema más tratado en la literatura de la destreza manual. Geshwind sugiere que los niveles altos de testosterona prenatal enlentece el crecimiento del hemisferio izquierdo en relación al derecho, llevando a, entre muchas otras cosas, a una mayor incidencia de zurdos. Galaburda ofrece una versión revisada de esta hipótesis ej a mayor testosterona lleva a menor reducción neuronal en el Hemisferio derecho, y por lo tanto a un tamaño relativamente más grande en el planum temporale derecho. Hay varias formas de estudiar la interrelación entre testosterona y destreza manual. En humanos los niveles hormonales del 2do trimestre en el líquido amniótico han sido evaluados y relacionados con la destreza manual examinada a la edad de 7 años. Entre chicas con mayores niveles de testosterona predicen más destreza manual derecha, mientras que esta relación no fue vista en varones. Otra estrategia de investigación involucra el análisis de la destreza manual en gemelos dicigóticos del mismo sexo o de sexo opuesto. Las mujeres con un varón gemelo están expuestas a altos niveles de testosterona que mujeres con gemela mujer; similarmente los varones con varón gemelar están expuestos a altos niveles de testosterona que aquellos con una gemela mujer y por lo tanto, cada uno debería exhibir una elevada incidencia de la destreza manual izquierda. Otro estudio reciente no observó tales efectos. 2 estudios han encontrado que las mujeres expuestas al dietilestilbestrol en útero, una hormona masculinizante, demuestra una incidencia aumentada de zurdos en adultos. Obviamente, es dificultoso hacer conclusiones firmes con respecto a los niveles de testosterona y la destreza manual. INFLUENCIAS PERINATALE SY POSTNATALES EN LA VARIACIÓN INDIVIDUAL EN LA DESTREZA MANUAL. La noción que el trauma perinatal causa variación en la destreza manual tiene una larga historia . No cabe duda que el daño en el hemisferio izquierdo franco puede llevar a algo que ha sido llamado “ destreza manual izquierda patológica”, A causa que muchos ñ están predispuestos a “ la destreza manual derecha más que a la izquierda ” la incidencia de destreza manual (DM) izquierda patológica será mayor que la de la DM patológica derecha. La discusión de esto es: 1ro ¿Cuál la incidencia de la DM izquierda patológica? 2do ¿hay alguna relación DM y tipos stress al nacer menos serios que el daño en un hemisferio cerebral izquierdo? Y si lo es, ¿es la interrelación causal o correlativa? Satz establece que la DM izquierda patológica es acompañada por injuria del hemisferio cerebral izquierdo, dificultades motoras de la mano derecha, lateralización atípica del habla, e hipoplasia de pie derecho. Estos criterios surgen de poblaciones clínicas específicas, sin embargo, no permite una estimación de la incidencia en la población con DM izquierda patológica. Bishop argumentó que si la DM patológica contribuye más a la DM izquierda que a la derecha, luego la proporción de habilidad motora deficiente en la mano no preferida deberá ser mayor en zurdos que en diestros. Ella analizó informes de 12000 ñ del estudio del Desarrollo Infantil Nacional en 2 tareas motoras: 1) un test de marcación cuadrada usando implementos de escritura 2) el tiempo para transferir 20 fósforos de una caja a otra. El patrón esperado basado en la hipótesis de DM izquierda patológica, ej mayor incidencia de una ejecución muy pobre con la mano no preferida en los zurdos, fue observado sólo en el test de la caja de fósforos. Para este test Bishop estimó que 1de cada 20 de los zurdos debería ser considerado patológico. Sin embargo, esto probablemente es una sobreestimación de la verdadera incidencia, como no hay razón para esperar la DM patológica surja sólo en el test de la caja de fósforos y no en el otro test. En algún caso, la vasta mayoría de los zurdos no parecen sufrir una disfunción en el hemisferio izquierdo temprano, y por lo tanto, algún otro mecanismo debe estar operando la producir una DM izquierda. ¿Hay una relación entre mayor stress al nacer y la DM izquierda? La mejor respuesta surge de un metanálisis de Searleman. El no encontró evidencias que el parto esté relacionado con la DM, si bien el parto es un marcador relevante para una gran variedad de factores estresantes al nacer. La edad materna avanzada, los partos múltiples, el parto rápido, el parto lento, y dificultades respiratorias fueron también no relacionados en varones y mujeres. Entre los varones, la incompatibilidad RH, el bajo peso al nacer, la prematurez, el parto por cesárea, parto en podálca fueron escasamente relacionadas a la DM, explicando al menos un 1 % de la variancia. Los factores de stress no relacionados al parto fueron predictivos en las mujeres. Hay una correlación positiva escasa entre stress en el parto y DM izquierda. Aún continúa demostrarse, sin embargo, que alguna de tales relaciones son causales, como es posible que fetos con alguna forma de anormalidad

7
subclínica o leve antes del nacimiento son más proclives a la complicación perinatal y la revisión de las evidencias predichas revela diferencias entre zurdos y diestros durante el desarrollo fetal. Ha tenido dificultad en identificarse las influencias ambientales específicas en DM durante la infancia. Posiblemente, la DM está influenciada por instrucciones directas o imitación desde los padres o hermanos. Una posibilidad contrastante es que los factores ambientales no-biológicos postnatales son no relevantes para la variación individual en la DM. En estudios genéticos conductuales, las influencias no genéticas se conceptualizan diviéndose en 2 amplias clases. Aquellos factores sociales y ambientales que son más compartidos por miembros de una familia llamados “factores compartidos” e influencias únicas a un miembro de una familia llamados “factores únicos”. Los puntajes de estudios de estudios en las 2 décadas pasadas de todas las variables psicológicas han revelado que los factores únicos son más importantes que los compartidos. Cuando los efectos compartidos son observados, ellos frecuentemente disminuyen a medida que el ñ crece. En el área de la inteligencia, por ej, los estudios de adopción han encontrado que los efectos ambientales compartidos están asociados en 4-5% de la variancia en el puntaje de CI en jóvenes no biológicamente relacionados con sus hermanos, pero esto disminuye de 0 % hacia los 18 años. La forma más estricta para estimar la importancia relativa de los factores compartidos o únicos ambientales es a través de estudios de adopción. Los padres adoptivos no compartían la carga genética de sus hijos adoptivos. Por lo tanto, la semejanza entre ellos debería reflejar probablemente los efectos compartidos, ej ñ enseñados a usar una determinada mano para una tarea dada, o simplemente imitando a sus padres. El grado al cual los ñ adoptados se semejaban a sus padres biológicos proporcionó la medida de la influencia genética. Un importante estudio de adopción de Carter brindó algún esclarecimiento en este tema. Notablemente, la DM en la descendencia fue relacionada a sus padres biológicos y a no a sus padres adoptivos. Si la imitación o el entrenamiento del uso de la mano izquierda fue causalmente relacionada al uso de la mano izquierda en ñ, uno debería esperar que los padres adoptivos quienes eran zurdos deberían tener más ñs adoptados quienes fueron zurdos. Comparamos la incidencia de la DM derecha en ñ adoptados con padres zurdos (86%) versus aquellos con ninguno (87%) en estudio de Carter. Este informe marca un caso preciso: que las influencias de los padres en la casa no contribuyen a la variación individual en la DM. Por supuesto es posible que los factores únicos ambientales contribuyen a las variaciones en DM. Sin embargo la identificación de factores “únicos” es más compleja que los factores compartidos, e importantes estadísticas confluyen a influencias social unica e influencia genética ni direccional como discutimos con mayor detalle más adelante. Hay evidencias en efecto que injurias severas para un brazo o mano pueden cambiar la DM en una proporción pequeña de individuos, pero esta clase de efecto parece improbable a ayudarnos a entender el desarrollo de una DM atípica en la vasta mayoría de individuales con ausencia de trauma. Con respecto a esto es importante acordarse de que las impresionantes evidencias de una estabilidad de la habilidad manual relativa en la infancia y la adultez a lo largo de la vida, de los informes surgen cuestiones serias acerca de la posible importancia de los determinantes ambientales “únicos” postnatales. FACTORES GENETICOS EN LA DESTREZA MANUAL Las teorías con respecto a la herencia de la DM se retrotraen a la 1ra parte de este siglo. Aún no existe un consenso con respecto al componente genético, o si es lo es, exactamente en qué aspectos del fenotipo va a estar el foco del modelo genético. En la próximas 2 secciones son tratadas evidencias de las influencias genéticas en la preferencia manual y la habilidad manual relativa. Luego, son tratados nuevos datos que sugirieren la importancia de genes específicos o cromosomas. Finalmente, modelos genéticos de DM son analizados. PREFERENCIA MANUAL El estudio de adopción de Carter sugiere que los factores genéticos son importantes para explicar el parecido familiar en DM: Los descendientes adoptados se parecen a sus padres biológicos, no a los padres adoptivos. Parecería ser una situación muy simple a aplicar en las técnicas de investigación genética conductual para dar una estimación de la importancia relativa de lo genético como también de las fuentes de variación ambientales “únicas” o “compartidas”. Sin embargo, no hay acuerdo de la magnitud de las influencias genéticas surgidas hasta el momento y es importante entender por qué. Un problema importante es que el fenotipo de la DM es edad-dependiente y multifacético. ¿Queremos nosotros un modelo de preferencia prenatal asimétrica, o el fenotipo influenciado socialmente de la mano que escribe, las continuas asimetrías de los sustratos corticales putativos de la habilidad manual asimétrica o la asimetría de la habilidad en tareas de movimiento “peg”? ¿Queremos el modelo de asimetrías motriz fina , o

8
asimetrías de la motricidad gruesa o de arrojar cosas?¿ Queremos un modelo de asimetría direccional o del grado de desviación desde la media de la población, qué parece ser heredado en especies no humanas . La respuesta obvia es que nosotros deberíamos tener un análisis genético conductual en general de tales aspectos de la DM, pero la vasta mayoría de los análisis examinaron sólo la variable dicotómica de la mano preferida para la escritura. Este informe básico es óptimo para testear una teoría genética de la DM que postula 2 categorías discretas de DM (Mc Manus), adecuadas para testear una teoría genética que postula 3 diferentes genotipos, e inadecuada para testear una teoría genética que postula el grado de atipicidad, en ambas dirección desde la media poblacional, es hereditaria. Otra dificultad inherente en los estudios genéticos de la DM es la cuestión del uso útil de modelos de gemelos, la llave maestra de la genética conductual contemporánea. Hay temas variados surgidos con respecto a los gemelos, si bien, como es típico en este campo, el consenso ha demostrado ser fugaz. Algunos han sugerido una incidencia elevada en la DM izquierda en gemelos (Annet) o en los padres de gemelos. Otros han señalado que algunos gemelos monocigotas son entre ellos “imágenes en espejo”, hacia la dirección de un patrón en espiral, un factor que complica el análisis de lateralidad. Algunos gemelos monocigotas (MN) son monocoriónicos, compartiendo la misma membrana fetal (30%), mientras otros son dicoriónicos, los cuales ocurre por la partición del huevo que tiene lugar 72 horas después de la fertilización. Un estudio reciente no encontró evidencias que el estado del corión influencie en la DM izquierda o concuerde entre gemelos MZ, pero puede tener un poder limitado para detectar tales efectos, En algún caso, los datos surgidos de estudios en gemelos no parecen ser consistentes con los datos surgidos de estudios en familias. En el análisis genético conductual de otros rasgos, la combinación de las influencias de la casa y genéticas inherentes en los estudios familiares les rendiría a ellos mucha menor preferencia como diseños para un análisis genético que estudios de gemelos. Si uno asume, sin embargo, que el ambiente familiar contribuye algo a la DM (como sugiere Carter en estudio sobre adopción) los estudios de gemelos tienen sus propios problemas únicos en la DM el cual puede o no ser el caso), los estudios familiares puede suministrar el mejor dato disponible. Una cosa nos es segura: la preferencia manual transcurre en familias. En un reciente metanálisis de Mc Manus revisó la incidencia de DM izquierda en 72000 individuos. Donde ambos padres fueron diestros, el 9,5% de sus descendientes fueron zurdos. Donde un padre fue zurdo y el otro fue diestro el 19,5% de su descendencia fue zurda, Si la madre fue zurda el 21,9% de la descendencia fue zurda, en oposición al 16,8% si el padre fue zurdo (el llamado efecto maternal). Finalmente, si ambos padres eran zurdos, el 26,1% de su descendencia fue zurda. Estos datos también pueden ser expresados como una incidencia. El promedio de incidencia, el n° de ñ nacidos zurdos de padres en los cuales al menos un padre fue zurdo dividido por el n° de ñ nacidos diestros de tales padres, es dividido por el promedio de otra incidencia, el n° de ñ zurdos nacidos de padres diestros dividido por el n° de ñ nacidos diestros de tales padres. El promedio de tal incidencia es de 2.08 para el metanálisis de Mc Manus y representa una expresión cuantitativa de la mayor tendencia de los ñ zurdos de nacer de un grupo de padres con al menos uno zurdo que de aquellos con los 2 padres diestros. Estos patrones familiares deben ser explicados por alguna teoría, ambiental o genética, que proponga explicar la variación individual de la DM. Un importante estudio ha examinado el parecido de la preferencia manual en familias de India y Canadá (Mc Manus). Este autor obtuvo datos en mano preferente usada para 10 tareas tanto en ñ como en sus padres. Ellos también intentaron evaluar la herencia de la motricidad gruesa de la mano (sin tomar en cuenta la dirección) y el extremo de DM, variables importantes para determinar el esquema genético descripto más adelante. En Canadá, la incidencia de tener un ñ zurdo cuyo uno de sus padres era zurdo fue aproximadamente el doble con respecto de aquellos donde ambos padres eran diestros, mientras que en India la desigualdad fue del triple. La herencia de la preferencia manual gruesa, omitiendo la dirección de la preferencia, fue también significativa en ambos países, y la incidencia fue mayor que para la de dirección de preferencia. El efecto también fue mucho más grande en India que en Canadá. Datos similares fueron dados por Coren , quien encontró escasas, si bien significativas , correlaciones a nivel no direccional de asimetría preferente de la mano, ojo, pie, y oído entre miembros de una familia. Ellos no encontraron correlaciones significativas para la dirección de preferencia. El mismo tipo de efecto también ha sido demostrado por Collins en ratones. En un análisis relacionado, Bryden también examinó la herencia de extremos, en los cuales es relevante para un modelo de desarrollo inestable de DM genética. El puntaje de la preferencia manual fue dividido en cuartos, con 2 mitades llamada moderada y las otras 2 llamadas extremas. Fue computada la incidencia de (a) 2 padres, al menos uno de ellos extremo, teniendo un ñ con DM extrema, a b) 2 padres sin la DM extrema teniendo un ñ de DM extrema, y encontraron que era significativa en el ejemplo de India pero no en el Canadá.

9
2 recientes estudios resumieron la investigación llevada a cabo antes 1990 en preferencia manual de gemelos (Mc Manus). Agregando datos de otros estudios, ellos sugieren que la proporción global de concordancia para preferencia manual es casi idéntica a los gemelos MZ y DZ. En contraste con algunos estudios pero consistentes con otros la DM izquierda no fue más común en gemelos que en la población general. Los cálculos de incidencia indican una gran asociación de zurdos en varones gemelos MZ que en varones gemelos DZ; para las mujeres, fue notada una tendencia en la misma dirección. La DM materna fue asociada con DM tanto descendencia masculina como femenina, pero la DM por parte del padre influenció sólo a la DM de los hijos varones, surgiendo la posibilidad de un efecto cromosómico Y. La DM izquierda fue más común en el 1er gemelo que en le 2do, y entre el 1ro, la incidencia de DM izquierda aumentó en el 2do gemelo cuando fue más pesado que el 1ro. Los autores sugieren que la DM izquierda” está asociada con disturbios en el crecimiento intrauterino por la cual una gran disparidad en el peso al nacer de ambos gemelos es una indicación y que el 1er gemelo que nace podría ser más vulnerable cuando su peso es menor que el del 2do gemelo”. Los estudios dan una oportunidad para comparar la herencia entre gemelos versus estudios familiares, Uno puede calcular la incidencia limitada a gemelos MZ y DZ y sus padres. Reanalizando estos datos originales la incidencia es 1.31, notablemente menor que el valor de 2.08 de Mc Manus o el de 2.29 de Carter. Los factores determinantes de la DM en gemelos puede por lo tanto ser algo diferente que aquellos realizados en no gemelos. Por lo tanto, uno probablemente debería ser cauteloso en generalizar conclusiones surgidas desde estudios en gemelos a la población que no lo es. HABILIDAD MANUAL RELATIVA Pocos estudios han examinado la herencia de la habilidad manual relativa. ANNET no observó parecido al menos en la velocidad de movimiento “peg” derecho entre todas de los miembros de una familia (ej padre-hijo. Madre-hija) excepto para una débil relación entre madre-hija. Una tarea computarizada de movimientos “peg” permitiendo medir en forma separada el “tiempo de transporte” (el tiempo empleado en mover una clavija al próximo agujero) y el “tiempo de búsqueda” (el tiempo empleado en buscar para agarrar la próxima clavija) fue usado en un gran estudio llevado cabo en ñ y sus padres. El tiempo de transporte estaba más fuertemente relacionado con la preferencia manual y mostró un mayor grado de asimetría. La DM materna predijo una simetría reducida estricta en ambas tareas en ñ, mientras la asimetría sólo se redujo en el tiempo de búsqueda y fue predicha por la DM paterna. Carlier examinó hermanos con el test dot-filling y el test tapping computarizado y luego fueron analizados 2 mediciones del mismo, la diferencia en velocidad entre las 2 manos y la regularidad de tocarse en forma repetitiva el dedo pulgar con el índice (el promedio de la desviación Standard en el intervalo intertap entre la mano preferida y la no preferida.). El parecido entre hermanos fue encontrado sólo para la tarea de dot-filling. Investigamos la posibilidad de hasta que punto un individuo puede heredar, el grado en el cual un individuo difiere de la población en la asimetría en la habilidad típica. En esencia pensamos que esto refleja la versión humana de la hipótesis Collins que el grado pero no la dirección de la asimetría es hereditaria, la sóla diferencia está en que la media de la población humana es desviada desde lateralidad cero hacia mayores habilidades diestras. Usando el test de “pegboard” de Annet determinamos para cada sujeto hasta el que punto al ellos se desviaron del ejemplo promedio, Predijimos que individuos con DM extrema, ya sea habilidad manual diestra, o relativamente mayor que el promedio de habilidad manual diestra, debería ser más apto para tener padres que ellos informaron ser zurdos, como la preferencia manual es más atípica que en diestros. Fue observada una relación en forma de curva, de modo que individuos ya sea en el final de la distribución de la habilidad manual relativa (izquierda menos derecha) fueron más aptos para tener padres zurdos, consistente con nuestra hipótesis de hasta que punto el grado de atipicidad es hereditaria. Annet no observó esta relación en un análisis similar, sin embargo, los datos esperan información adicional. La mejor forma de testear la idea es explorar relaciones atípicas en habilidad manual tanto en los padres como en los ñ, más que depender de los informes de preferencia manual de los padres. Comparando estudios genéticos de preferencia manual y ejecución, 2 observaciones deberían tenerse en cuenta: la 1ra como fue notado en estudios de influencia genética en la preferencia, los estudios familiares proveen algo más que un soporte al factor genético que los estudios en gemelos. 2do hay evidencias menos notables de lo heredado de la habilidad manual relativa que en la preferencia manual. Una razón posible es que la medición de preferencia, se responde través de un n° de tareas compuestas y por lo tanto proveen una clase de medición de DM, mientras que la medición de la habilidad manual relativa lo hace desde una tarea única.

10
Quizás, una medición compuesta de habilidad manual relativa a través de un n° de tareas debería tener una herencia más similar que las mediciones de preferencia manual. Esto rendiría estudios de herencia de la DM más probablemente que estudios de la herencia de la inteligencia, lo cual depende de evaluaciones de habilidades compuestas. INTENTO PARA IDENTIFICAR INFLUENCIAS GENETICAS ESPECÍFICAS Está bastante bien establecida la contribución genética de la variación en la DM. El próximo paso es identificar las influencias genéticas específicas ej alelos individuales o cromosomas. O clases de factores genéticos. Yeo ha intentado vincular variaciones en la DM a una serie de factores genéticos. El hipotetizó que variaciones de la media poblacional a la habilidad manual relativa en humanos (asimetría direccional) refleja los mismos factores que la variación determinada alrededor de la media poblacional (0) a otros asimetrías caracterizadas por asimetría fluctuante (AF). Ha habido mucha investigación en diversas especies, como los determinantes de la magnitud de la AF y han sido encontradas tanto influencias genéticas como ambientales. Los factores genéticos incluyen una carga genética, complejos genéticos alterados o coadaptados mediante recombinaciones sexuales, y posiblemente, homocigóticos poligénicos. Los factores ambientales incluyen stress fisiológico materno, toxinas, y agentes patógenos. Los factores maternos son más aptos para influenciar en la estabilidad del desarrollo ej la habilidad de un organismo a ejecutar completamente su programa ontogénico a desarrollar, en genes susceptibles. Las 2 expresiones del fenotipo más importantes del desarrollo inestable son la presencia de características fenotipo-desviadas morfológicamente como las MPAs (características atípicas morfológicamente) y la Asimetría Fluctuante. En 3 estudios de índice de desarrollo estable, que constan de FA y MPAs de variadas características corporales, fue relacionado para medir la preferencia manual y la habilidad manual relativa. El total del puntaje de la MPAs y la FA fueron hechos con score “z” y combinados para proveer un índice total de desarrollo inestable. El grupo de sujetos con preferencia manual zurda tuvieron una mayor inestabilidad en el desarrollo que aquellos con una preferencia manual diestra. La relación entre la habilidad manual relativa y la inestabilidad en el desarrollo se muestra en la figura 4. 2 características son de mención. Una desviación de la superioridad de la habilidad manual diestra moderada, en esa dirección, un aumento del desarrollo inestable. 2do el valor de la ejecución manual relativa asociada con inestabilidad mínima del desarrollo está más próximo del valor medio de la habilidad manual relativa. Las implicaciones teóricas de esta observación son discutidas en la próxima sección. Sin embargo, la asociación de marcadores del desarrollo inestable con la DM sugieren de la importancia de una relación entre factores genéticos y ambientales conocidos del desarrollo inestable en la variación individual en la DM. Una medición extrema de características físicas (alteración en la morfología de los pies) fueron relacionadas con la DM atípica. Tales atipicidades podrían reflejar una carga en la mutación global u homocigótica poligénica. La preferencia manual izquierda ha sido vinculada con alelos particulares en el sistema antígeno leucocitario humano (HLA). Este estudio sugirió que la DM izquierda estaba asociada con enfermedades autoinmunes de la tiroides (Enfermedad de Graves y la tiroiditis de Hashimoto), enfermedades autoinmunes del tubo digestivo (enfermedad celíaca y la enfermedad de Crohn) y el Lupus eritematoso sistémico. Cada uno de estas enfermedades han sido vinculadas con el mismo grupo de alelos HLA en el cromosoma 6: A1, B8 y DR3. Estos alelos están en una unión en desequilibrio y por lo tanto tienden a agregarse dentro individuos. Los individuos con preferencia manual izquierda fueron más aptos para tener los alelos B8 y el DR3. Es incierta la manera en la cual estos alelos, u otros que están unidos en forma desequilibrida, están relacionados a DM. Una clave puede ser que el alelo B8 y el haplotipo A1/B8 es poco predictivo en la descendencia que el alelo individual. Especulamos que estos alelos pueden afectar adversamente (o pueden estar vinculados con otros alelos que afectan) ya sea la respuesta inmune de la madre o del ñ siendo una amenaza patógena al desarrollo fetal y por lo tanto influencia en el desarrollo estable del feto, produciendo últimamente DM atípica. De acuerdo a esto, se informó que las infecciones maternas aumentan la AF en la descendencia. El alelo DR3 parece estar vinculado a la gran incidencia de aborto espontáneo. Otro estudio reciente señaló el rol de los genes en el cromosoma sexual. Estudios en gemelos describieron la posibilidad originada de un gen o genes en el cromosoma Y relacionado a la preferencia manual. Un estudio de un grupo de 5 informes proveyó información en al menos 15000 pares de hermanos también surgió un locus cromosómico sexual. Acerca del 10% de pares de hermanos del mismo sexo fueron más concordantes a la DM que los pares de hermanos de sexo opuesto. Esto es consistente con la posibilidad de un gen o genes en los cromosomas sexuales relacionados con la preferencia manual.

11
Corballis expresó escepticismo acerca de la existencia de alelos diferentes para la DM en el cromosoma Y .El demostró mediante procesos de modelamiento que los polimorfismos en el cromosoma Y fueron muy improbables para mantener una incidencia estable en la población de varones dadas muy pequeñas diferencias entre los sexos en la consecuentes adaptaciones asociadas con su gen. Quizás los genotipos XY y XX pueden ocasionar alguna diferencia en el crecimiento dinámico prenatal, afectando la DM en formas consistentemente diferentes. En general Rice ha mostrado que algunos “trabajan mejor” cuando son masculinos y otros cuando son femeninos. En esta forma, estos genotipos pueden no sólo conducir a leves diferencias sexuales en la incidencia de la DM, sino también modelan diferentemente los procesos del desarrollo conducentes a la DM. Si el stress del desarrollo contribuye a la DM izquierda podría explicar la diferencia en las incidencias concordantes a través de los pares de hermanos de sexo opuesto y del mismo sexo. Esta posibilidad representa más de un “cromosoma entero” o influencia genética que los efectos de un simple cromosoma. Otro estudio reciente investigó la posibilidad de un locus del cromosoma X para ambas DM y psicosis, reflejando la teoría de Crow que un solo gen puede relacionarse a ambos, DM izquierda y la tendencia a la psicosis. En un grupo de familias en las cuales pares de hermanos fueron afectados con psicosis, un locus cercano al centrómero fue más apto para ser compartido por hermanos varones afectados que hermanos varones no afectados, pero no en hermanas mujeres. En un 2da familia ejemplo en la cual un par de hermanos varones fueron zurdos, un locus en una región similar del cromosoma X fue asociado con un grado DM izquierda, pero no en la preferencia manual zurda versus diestra. Estos resultados son intrigantes. ¿Por qué es el efecto psicótico sólo encontrado en los pares de hermanos varones y la observación con respecto a la habilidad manual puede generalizarse a todos los tipos de hermanos? Basados en informes de un nuevo grupo de familias, han sugerido la transmisión vinculada al cromosoma X para la preferencia manual. Se destacó que “la madres zurdas producen significativamente más hijos zurdos que hijas zurdas, que los padres zurdos producen significativamente más hijas zurdas que hijos zurdos, y que los padres zurdos no producen más hijos zurdos que los padres diestros. El cromosoma X debería predecir una mayor incidencia de DM zurda en varones que mujeres, pero la incidencia de DM izquierda en varones es sólo el 2% mayor que la incidencia para las mujeres. Para explicar esta leve diferencia en sexos McKeever hipotetizó que las mujeres puede ser más vulnerables a causas no genéticas de la DM izquierda, tales como un desarrollo inestable o stress al nacer. Para nuestro conocimiento, esto no es un soporte empírico para la gran incidencia de desarrollo inestable en mujeres, y el metanálisis de Searleman no encontró evidencias que el stress perinatal afectaba la DM en mujeres. Son necesarias más investigaciones antes que uno pueda concluir que han sido identificadas las influencias genéticas específicas sobre la DM. Los factores propuestos incluyen la mutación, homocigótico poligénico, los antígenos específicos HLA, genes en los cromosomas sexuales. Por supuesto, estas alternativas no se excluyen mutuamente. TEORIAS GENETICAS DE LA DM Una variedad de modelos han sido propuestos para explicar la naturaleza de los factores genéticos produciendo variación en la DM humana. El 1er tema confrontado por alguna teoría genética de la DM es la definición del fenotipo en particular, lo innato de la característica conductual y si bien es una variable distribuida continuamente o dicotómica. Hay muchas características relacionadas a las características de la DM, incluyendo la preferencia, habilidad motora fina y gruesa. Además, la DM es relacionada levemente con la gran cantidad de asimetrías no manuales, desde la dominancia del pie y del oído a la variación al menos de alguna habilidad cognitiva mostrando lateralización cerebral a nivel de la población. Como se destacó más arriba, está relacionado con una amplia variedad de asimetrías neuroanatómicas. ¿Cuáles son las fronteras apropiadas para una teoría de la DM? No hay dudas que la variación fenotípica en la mano usada para escribir puede ser expresada como una característica dicotómica, zurda o diestra, y eso probablemente caracteriza la mejor ejecución en muchos test de habilidad con papel y lápiz, tales como el test de Tapley y Bryden. Pero todas las otras manifestaciones de destreza manual y todas las otras relaciones de asimetrías de habilidad cognitiva y anatómica están tratadas continuamente. Un resúmen de los componentes centrales de las teorías genéticas de la DM está en la tabla 1 RESUMEN DE LS MAYORES DIFERENCIAS ENTRE LAS TEORIAS DE LAS VARIACIONES EN LA DESTREZA MANUAL
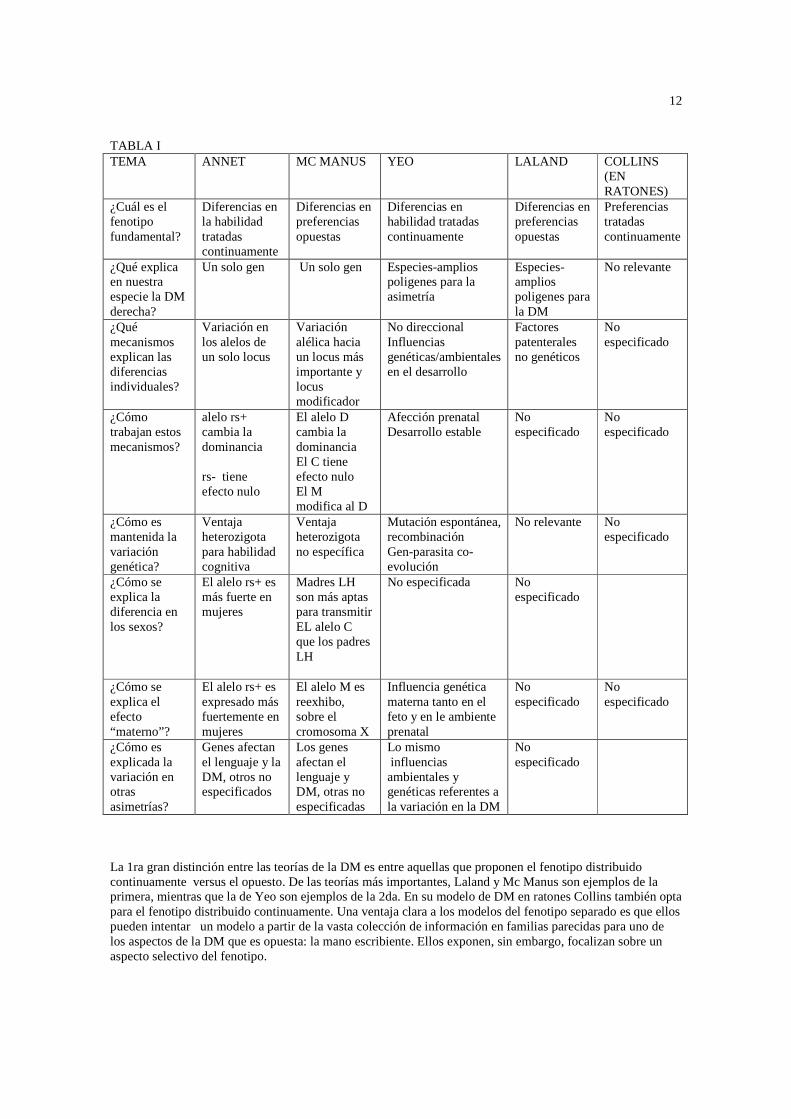
12
TABLA I TEMA ANNET MC MANUS YEO LALAND COLLINS
(EN RATONES)
¿Cuál es el fenotipo fundamental?
Diferencias en la habilidad tratadas continuamente
Diferencias en preferencias opuestas
Diferencias en habilidad tratadas continuamente
Diferencias en preferencias opuestas
Preferencias tratadas continuamente
¿Qué explica en nuestra especie la DM derecha?
Un solo gen Un solo gen Especies-amplios poligenes para la asimetría
Especies-amplios poligenes para la DM
No relevante
¿Qué mecanismos explican las diferencias individuales?
Variación en los alelos de un solo locus
Variación alélica hacia un locus más importante y locus modificador
No direccional Influencias genéticas/ambientales en el desarrollo
Factores patenterales no genéticos
No especificado
¿Cómo trabajan estos mecanismos?
alelo rs+ cambia la dominancia rs- tiene efecto nulo
El alelo D cambia la dominancia El C tiene efecto nulo El M modifica al D
Afección prenatal Desarrollo estable
No especificado
No especificado
¿Cómo es mantenida la variación genética?
Ventaja heterozigota para habilidad cognitiva
Ventaja heterozigota no específica
Mutación espontánea, recombinación Gen-parasita co-evolución
No relevante No especificado
¿Cómo se explica la diferencia en los sexos?
El alelo rs+ es más fuerte en mujeres
Madres LH son más aptas para transmitir EL alelo C que los padres LH
No especificada No especificado
¿Cómo se explica el efecto “materno”?
El alelo rs+ es expresado más fuertemente en mujeres
El alelo M es reexhibo, sobre el cromosoma X
Influencia genética materna tanto en el feto y en le ambiente prenatal
No especificado
No especificado
¿Cómo es explicada la variación en otras asimetrías?
Genes afectan el lenguaje y la DM, otros no especificados
Los genes afectan el lenguaje y DM, otras no especificadas
Lo mismo influencias ambientales y genéticas referentes a la variación en la DM
No especificado
La 1ra gran distinción entre las teorías de la DM es entre aquellas que proponen el fenotipo distribuido continuamente versus el opuesto. De las teorías más importantes, Laland y Mc Manus son ejemplos de la primera, mientras que la de Yeo son ejemplos de la 2da. En su modelo de DM en ratones Collins también opta para el fenotipo distribuido continuamente. Una ventaja clara a los modelos del fenotipo separado es que ellos pueden intentar un modelo a partir de la vasta colección de información en familias parecidas para uno de los aspectos de la DM que es opuesta: la mano escribiente. Ellos exponen, sin embargo, focalizan sobre un aspecto selectivo del fenotipo.

13
La 2da mayor distinción entre las teorías es si el fenotipo está mejor representado por la habilidad o la preferencia. Esto es un aspecto del fenotipo que puede inevitablemente llevar a otro. En ese sentido la elección puede no ser decisiva. La incidencia de la asimetría en preferencia y habilidad se muestran totalmente diferentes, si bien, como la preferencia tiene una forma de J mientras que la la 2da es cercana a lo normal. Por lo tanto las asimetrías en habilidad ofrecen la posibilidad de lograr una mayor discriminación entre los fenotipos. Por ej, la distribución de asimetría en habilidad puede distinguirse entre la vasta proporción de la población quien está descripta a sí misma como diestra, mientras los inventarios de preferencia están limitados con respecto a esto. Sin embargo, hay obviamente limites sobre algunas actividades pueden medirse en un individuo, y algunos estudios de preferencia manual tienen beneficios prácticos reales. Siguiendo a Annet, nuestra inclinación ha sido para mediciones en la habilidad, mientras que para Laland y Mc Manus preferien enfocar sobre preferencia, como lo hace Collins. 2 diferentes fenómenos diferentes deben ser explicados:1) La gran tendencia de la especie hacia la DM derecha, y la 2da es la variación individual. Annet y Mc Manus sugieren que un único gen hace a los humanos diferentes en términos de dominancia de lenguaje y DM; variaciones en los alelos de ese locus único explica todas las variaciones genéticas en la población. Laland y Yeo sugieren que muchos genes, compartidos entre todos los miembros de las especies, contribuyen a obtener una asimetría humana diferente de otros animales, pero estas teorías difieren en cómo explican la variación individual. El modelo de desviación derecha de Annet y el modelo de Mc Manus han sido revisados en detalle. TEORIA DE LA DESVIACIÓN DERECHA DE ANNET. El mejor modelo genético conocido de DM es el modelo de desviación derecha de Annet. Annet propone que determinados alelos, el alelo desviación derecha (o rs+), tiende a enlentecer el desarrollo in útero del hemisferio derecho, inclinandose el crecimiento cerebral, como un aumento probablemente, a la dominancia del hemisferio izquierdo para el lenguaje y la DM derecha. Otro alelo, el alelo de desviación derecha negativo (rs-) , no tiene tal predisposición. Annet ha argumentado siempre un elemento “al azar” en el desarrollo de la DM, tal como factores aleatorios actúan para influenciar el fenotipo. Sin embargo, el gen rs+ explica el 50% de la variación fenotípica. En individuos con 2 copia del alelo rs-, los factores aleatorios junto con las influencias ambientales producen la variación fenotípica. Los efectos genéticos son sumados (si bien en versiones tempranas de su modelo el alelo rs+ fue dominante). Estimaciones de la frecuencia de alelos fueron extraídas de la incidencia de individuos afásicos luego de la lesión en el hemisferio derecho ( rs+= 0.57 rs-= 0.43) y pueden ser usados para generar frecuencias de fenotipo(rs-/rs-= 018 rs+/rs-=0.49 ; rs+/rs+= 0.32). Annet supone que el alelo rs+ está expresado más fuertemente en la mujer que en el hombre. Esto le permitió explicar la mayor evidencia de DM izquierda en el hombre. Esto también ayuda a explicar el efecto materno (si bien Mc Manus señalaron que este detalle se del modelo de Annet predice un menor efecto maternal que el observado). Annet asume que el alelo rs+ es expresado más fuertemente en una persona que en gemelos. Dados estos supuestos, el modelo de Annet predice satisfactoriamente patrones de parecido familiar sin tomar en cuenta el sexo. Otra característica importante del modelo de Annet es que sostiene especificar la selección eficaz manteniendo la frecuencia del gen hacia un porcentaje estable. La hipótesis es que los 3 genotipos diferentes estarían asociados con diferentes patrones de habilidades cognitivas. El genotipo rs+ se dice tener poca consistencia relativa en razonamiento matemático y espacial, debido al excesivo impedimento del hemisferio derecho en desarrollo. El fenotipo rs- se dice que tiene poca consistencia en el procesamiento fonológico debido a la carencia de dominancia cerebral para el lenguaje. El genotipo heterozigota (rs+, rs-) no sufre ninguna de estas desventajas y es por lo tanto favorecido por la selección natural. Las ventajas de alelos heterozigotas mantienen la variabilidad genética a lo largo de generaciones porque este genotipo no puede reproducirse exacto. Una cierta proporción de la descendencia de 2 heterozigotas con ventajas, son inevitablemente genotipos rs,+rs+ y rs-,rs-desventajados, El supuesto empírico de estas asociaciones predictivas de patrones de habilidad cognitiva con DM es asociado a mejor. TEORÍA DE MC MANUS Mc Manus ofrece un modelo genético que, como hizo la propuesta de Annet en un locus autonómico con 2 posibles alelos que trabajan en forma sumatoria, los alelos D (para diestros) y el C (al azar). El genotipo DD

14
siempre produce DM derecha, mientras que el tipo CC resulta en asimetría fluctuante, con 50% de zurdos y el 50% de diestros. El genotipo DC produce 25% de zurdos y 75% de diestros. La frecuencia genética fueron cambiadas para optimizar el ajuste al modelo de informes de DM en familias. Los informes de gemelos están acondicionados para presumir que en el genotipo CC y DC el azar opera independientemente en cada par. Hay 3 grandes diferencias entre el modelo de Annet y el de Mc Manus La 1ra como de señaló más arriba, Mc Manus propone 2 fenotipos fundamentales, personas con preferencia manual derecha y personas con preferencia manual zurda. 2do factores aleatorios que operan sólo sobre los genotipos DC y CC, mientras que para Annet ellos afectan a cada de los 3 genotipos. 3ro Mc Manus propone un locus modificado sobre el cromosoma X . El dominante, el alelo M no tiene efecto sobre la lateralidad, mientras el alelo m recesivo permite la función del alelo D lo mismo que en el alelo C. Mientras los hombres tengan un solo cromosoma X el alelo m es más apto para expresarse, explicando la mayor incidencia de zurdos en varones como así también el efecto maternal. Alguna manera de ventaja heterozigota se presume para explicar el equilibrio estático de la frecuencia en los genes, pero la naturaleza de esta ventaja no está aún especificada. Ambos modelos proveen una adaptación razonablemente a los informes familiares. Kear ha ofrecido un modelo similar que también explica muchos informes familiares. Pudiendo adoptarse con precisión la información familiar, sin embargo, no significa que e modelo sea correcto( una adaptación precisa de modelo a informes indica que el modelo sólo es consistente con el informe. Salvo un gen específico es un hecho encontrar algo cualitativo ad hoc a esos modelos y sus supuestos. ¿Es biológicamente razonable que sólo un gen diferencia al hombre de otras especies en términos de lateralidad del lenguaje y DM? Sabemos de un fenotipo conductual complejo e el el hombre normal está determinado por 1 sólo gen, pero si los modelos del gen único son correctos, ellos son ciertamente únicos. Debe también responderse cómo estos modelos explican la variación en otras asimetrías que no sean la dominancia del lenguaje y DM. Las diferentes asimetrías cognitivas no se correlacionan, esencialmente ya que son asimetrías neuroanatómicas diferentes. Patrones como apretar con las manos o cruzarse de brazos y la dominancia ocular, aparecen en familias, sugiriendo un componente genético, y a lo mejor relacionando levemente con la DM (Mc Manus). Parece poco probable que el modelo del gen único explique los factores genéticos de otros aspectos de la lateralidad, que parece estar caracterizado por componente genético único en los humanos. TEORIA DEL DESARROLLO INESTABLE Proponemos que la variación de DM en el hombre es el resultado de un DESARROLLO INESTABLE (D I). Nuestro esquema dio lugar al dilema planteado más arriba. Si como parece ser el caso, la DM (destreza manual) tiene un componente genético, ¿cómo puede conservarse en una incidencia firme cuando parece estar en desventaja? Este esquema de DI comienza en un intento para entender la significación y causas de la AF desde una perspectiva biológica. La DI se refiere a la inhabilidad de los organismos para expresar completa y fielmente el plan especie-específico del desarrollo. Leves desviaciones de los diseños causadas por efectos fortuitos como mutaciones, agentes patógenos o toxinas, resultan en una expresión menos perfecta del plan ontogénico. Los individuos de una especie difieren en su exposición a perturbaciones perjudiciales y/o habilidad para resistir sus efectos. El modelo de la DI de la DM propone que 1) la DM derecha típica de la población en humanos se explica por un diseño de desarrollo universal culminando en una DM derecha moderada. Y 2) las desviaciones de la DM derecha modal resultan de un desarrollo inestable durante etapas tempranas del período prenatal. Sugerimos que los efectos genéticos en la DM no resultan de variaciones en los genes que codifican la lateralidad, como lo sustentan Annet y Mc Manus. Mejor dicho los efectos genéticos resultan de 3grupos de factores: 1) Amplios genomas característicos tales como complejos genéticos adaptados o perjudiciales y homocigotos poligénicos pueden influenciar susceptiblemente al desarrollo inestable 2) Las mutaciones pueden alterar el proceso de desarrollo 3) agentes patógenos pueden desorganizar el desarrollo y, genes que afectan susceptiblemente a agentes patógenos pueden también afectar el desarrollo inestable. En términos de preferencia la DM Izq es atípica y por lo tanto debería estar asociada con un mayor desarrollo inestable. En términos de habilidad manual la atipicidad en cada dirección de la media poblacional debería estar asociada con DM. Si bien las influencias genéticas subyacen a la DM, la relación fenotipo- genotipo es compleja y debería ser entendida en términos epigéneticos. El Control Epigenetico se refiere a los múltiples

15
factores genéticos y no genéticos que regulan o influencian la actividad durante el desarrollo. Según Klingerberg en su discusión del desarrollo biológico de la asimetría fluctuante establece que “la variación hereditaria de la AF es debida a una expresión modulada genéticamente de una expresión aleatoria que es en sí misma independiente del genotipo. Hay por lo tanto una dicotomía o separación no bien definida entre variación ambiental y genética”.Ellos notaron que relación no lineal entre influencias ambientales y genéticas y el fenotipo están omnipresentes en el desarrollo. Molenaar y colegas se refieren a estas influencias como la 3ra fuente de las diferencias en el desarrollo. A lo largo de los procesos genéticos y ambientales y sugieren que la variación en estos procesos epigenéticos puede ser considerado de origen caótico. En modelos biométricos Standard, esa fuente de variación es conseguida bajo el término de variación única o ambiental dentro de la familia. 2 estudios recientes al menos sugieren que el DI está asociado a un genoma menos regulado por las perturbaciones ambientales. En un ejemplo de gemelos MZ Y DZ de 7 años de edad usamos métodos para adaptarles modelos genéticos a una variedad de fenotipos físicos y conductuales. Los ñ que demostraron gran asimetría entre las manos en el test de WALLIM mostraron menor nivel hereditario para muchas mediciones físicas y cognitivas. Los niños diestros y zurdos extremos parecen contribuir de igual forma a este efecto. De esta forma, los factores ambientales y epigenéticos parecen ser relativamente más importantes en fenotipos con gran asimetría en habilidad manual. Esto es consistente con la hipótesis que plantea que una asimetría atípica en la habilidad manual refleja DI; el plan genético para el desarrollo es expresado en forma menos precisa, resultando en una mayor determinación epigenética y ambiental. En otros estudios, examinamos diferencias individuales como una función del DI en respuesta a la ingesta de cafeína. En tareas de memoria verbal la cafeína típicamente tiene un leve impacto negativo. Encontramos que los individuos con mayor DI (MPAs, FA) tuvieron un mayor impacto en tareas de aprendizaje verbal, luego de la ingesta de cafeína. Esto nuevamente es consistente con la noción de que los marcadores de DI denotan un individuo menos regulado por perturbaciones ambientales. También podría ayudar el explicar la observación que sostiene que los zurdos tienden a ser relativamente más sensibles a las drogas. El modelo de DI puede ser extendido a otras asimetrías cognitivas y neuroanatómicas. Suponiendo que esas varias asimetrías direccionales, como la habilidad manual, son parte de la naturaleza humana, y de esta manera, no hay diferencias individuales en el código genético para este tipo de asimetrías. Diferimos sólo en cómo fielmente el plan humano de todas estas literalidades es expresado, el cual esta determinado en gran parte por el desarrollo inestable. En un reciente estudio que demostró que el desarrollo inestable está relacionado a la variación en la lateralización de un grupo de habilidades cognitivas. La lateralización atípica fue predicha por los mismos marcadores de desarrollo inestable usados en nuestros estudios de destreza manual, pero no lo fue la dirección de la lateralización. Por ej. Un mayor DI fue asociado con ambos dominancia hemisférica izquierda para afectar el judging facial ( una tarea para la cual hay una inclinación en la población de dominancia hemisférica derecha) y mayor que la dominancia hemisférica derecha común. Algo de interés fue la observación que el componente de las anormalidades físicas menores de nuestra medición de DI estuvo más relacionada a DM, mientras que el componente de AF estuvo más relacionado con lateralización cognitiva. Como las AFMs son fijadas antes del nacimiento y el componente de la AF puede cambiar a lo largo de la vida, esto sugiere que el desarrollo de influencias en la destreza manual es ontogénicamente previo a aquellas de lateralización cognitiva. Si el desarrollo inestable está relacionado con asimetrías manual y cognitiva atípicas, uno debería esperar que estén relacionadas con las asimetrías neuroanatómicas. En un reciente estudio examinamos la relación entre AF corporal y una medida compuesta de asimetría cerebral atípica (asimetría del planum temporal atípica , asimetría del volumen de sustancia gris en un hemisferio cerebral atípico, asimetría del volumen d sustancia blanca cerebral atípica y asimetría de volumen del hemisferio cerebral atípico). Los mismos factores genéticos y ambientales que subyacen a AF de las características corporales producen variaciones de los patrones típicos de especies de asimetría cerebral. En un subgrupo de individuos, examinamos la relación entre mediciones combinadas de AF corporales y cerebrales (mientras éstos parecen reflejar el mismo desarrollo inestable subyacente) y la asimetría del tamaño de la región somatosensorial de la mano siendo esta correlación significativa (=0.63), demostrando que una mayor asimetría fluctuante es predictiva del sustrato de destreza manual humana. El modelo de Desarrollo Inestable provee una explicación correcta del por qué la destreza manual no diestra está asociada con muchos desórdenes del neurodesarollo (autismo, esquizofrenia, dislexia, retraso mental). Cada uno de estos desórdenes muestra también una elevada incidencia de los marcadores de desarrollo inestable. Por lo tanto, puede ser compartido, a través estos desórdenes que no son una anormalidad

16
específica, sino que una susceptibilidad general del desarrollo. La destrezas manuales atípicas, AF y las AFM por sí mismas representan manifestaciones benignas del desarrollo inestable. Su presencia sin embargo, denota un individuo que puede ser relativamente vulnerable a factores específicos y diferentes subyacentes a cada desorden del neurodesarrollo. En forma similar, el modelo de DI puede ayudar a explicar la gran incidencia de destreza manual derecha en hombres y mujeres homosexuales. También el modelo de DI provee una explicación simple de la mayor influencia de la destreza manual paterna y materna en la destreza manual de la descendencia. La destreza manual de la descendencia está influenciada no sólo por el grado de DI conferidos por los genes maternos y paternos, sino también por el desarrollo inestable de la madre a medida que se desarrolla el feto. El DI de la madre asociada con su propia destreza manual, tiene un efecto en el desarrollo del feto que el DI del padre no lo tiene. Como destacamos, una razón de por qué desarrollamos el modelo de la DI es porque las evidencias muestran que la destreza manual izquierda está asociada con fertilidad más reducida, más aún la destreza manual izquierda parece ser sostenida casi al mismo nivel durante el tiempo. La selección tiende a remover alelos perjudiciales, los cuales conducen a la cuestión de cómo la DM puede ser sostenida en la población El modelo de DI provee una explicación consistente con la evolución biológica. A pesar de la selección contra los alelos perjudiciales, la variación de los rasgos relacionados a la capacidad puede ser sostenida al menos de 4 formas: 1) mutaciones ocurridas en el ADN, muchas de las cuales son perjudiciales. Mientras la selección tiende a remover estas mutaciones, esta remoción no es efectiva para todas las mutaciones de una única generación. En un momento dado, los humanos probablemente poseen, en promedio, decenas de cientos de mutaciones perjudiciales que eventualmente son removidas por selección, con una variación sustancial del promedio debido al azar. 2) El ambiente selectivo no permanece constante. Los cambios rápidos en el ambiente selectivo pueden sostener variaciones genéticas en la capacidad. Las fuerzas claves selectivas a organismos de larga vida son patógenas, y ellas mismas evolucionan y por lo tanto cambian. El huésped patógeno coevolucionado puede sostener una variabilidad genética en la habilidad huésped para resistir a patógenos, incluyendo la capacidad de resistencia a enfermedades conferidas por los alelos HLA. Recientemente, ha sido sugerido que toxinas creadas por plantas que comemos pueden también coevolucionar con sus habilidades para resistir sus efectos y por lo tanto sostener la variación genética en nuestra resistencia de tales efectos. 3) otros procesos coevolucionados pueden sostener la variación genética en la capacidad. Por ej un conflicto entre los sexos o conflicto materno-fetal puede resultar en cambios continuos en la presión selectiva y por lo tanto sostener la capacidad. Los genes sexualmente antagónicos (en los cuales el alelo dado no puede trabajar igualmente mejor en hombres y mujeres) es un fenómeno relacionado. En períodos de tiempo, los alelos que no trabajan extremadamente bien en un sexo o el otro pueden ser sostenidos en la población, llevando a una variación sostenida en capacidad dentro de los sexos. 4) algunas características genómicas que no resultan genuinas pueden a ayudar a resistir perturbaciones del desarrollo. Los heterozigotas pueden aumentar la habilidad de un organismo para adaptarse a una variedad de desafíos para las capacidades mientras los heterozigotas a un único locus puede esperarse que tengan efectos generalmente pequeños en la capacidad, los efectos agregados de heterocigotos a través de un genoma puede ser significativos; un individuo que es heterozigota en algún locus no puede pasar información de rasgos a su descendencia. La variación genética en el DI puede ser sustancial Este esquema de DI ha generado numerosa observaciones que no pueden ser explicadas por la teorías de Annet y Mc Manus incluyendo las evidencias de la herencia del grado de DM, la asociación con los alelos HLA y la asociación con DM atípica con variaciones en el crecimiento y desarrollo. Es un modelo que permite ver cómo la variación genética es sostenida a pesar de la probable capacidad reducida de los zurdos. Nuestra observación que los diestros extremos (en términos de habilidad manual) tienen más padres zurdos (en términos de preferencia manual) fue un paso en el camino, pero Annett no replicó esta relación. Dado el hecho que la determinación de la DM en gemelos puede estar bajo menor control genético que en los hermanos no gemelos, la mejor clase de informes debería provenir de familias dando test múltiples de habilidad manual relativa. MODELO GENETICO CULTURAL Laland proveyó un nuevo modelo de DM humana. Ellos primero ofrecen un análisis cuantitativo de cómo los factores de selección pueden dirigir las frecuencias genéticas en un sistema genético basados en el modelo genético original de Mc Manus en que se asume un locus no modificador. Laland propone que los alelos D y C que actúan como se los describió más arriba, pero también propone que la tendencia cultural (especialmente de los padres) puede deformar la distribución genética del grado de variación. Ellos suponen que previa a la introducción del alelo D en el pool genético, los factores ambientales culturales y genéticos se suponen estar

17
en equilibrio. Los DD y CD tienen diferentes capacidades ventajosas sobre el genotipo CC y ellos afirman “nuestros parámetros reflejan nuestro acierto para explorar, si bien debería ser posible para explicar la variación de la DM sin tener en cuenta el beneficio heterozigota”. Sus análisis sugieren que por debajo de estos supuestos debe llegarse a un estado de equilibrio y que hacia este equilibrio, todas las variaciones genéticas han sido pérdidas, y ese estado final no es claramente consistente con cada uno de los modelos genéticos principales de DM. Sus análisis no realizan el test de aptitud de los modelos de Annett y Mc Manus para predecir las frecuencias genéticas, y no incluyen los beneficios heterozigotas. Sin embargo sugieren que cuando uno elimina la ventaja heterozigota, de la cual no existen evidencias sólidas, los modelos no permiten mantener la diversidad genética produciendo variación en la dirección de DM. Dada esta conclusión, Laland desarrolla su propio modelo, un modelo de “gene-culture” o modelo genético-cultural”. Ellos estiman la probabilidad que la descendencia de padres diestros llegan a ser diestros ( un efecto cultural) desde probablemente un análisis máximo de informes de estudios de antiguos de familias en la proporción de de ñ diestros y zurdos que habían nacido de diferentes combinaciones de padres (diestros - diestro, diestro- zurdo, zurdo-zurdo). Su modelo por lo tanto incluye un efecto genético universal así como algún tipo de influencia principal padre-hijo a un fenotipo similar, presumiblemente reflejando una imitación o entrenamiento. El análisis formal de este modelo indica un buen acceso a 16 de las 17 familias analizadas. Más aún, el modelo genera muy pocas expectativas a coeficientes entre gemelos y pares de hermanos, y es consistente con los porcentajes concordantes observados a través de 13 de los 14 estudios. Laland argumenta que es probable que todos los humanos tengan el mismo genotipo de DM. Al respecto es consistente con uno de los principios fundamentales del modelo de DI en que hay una variación no individual entre los humanos en el plan genético a la lateralidad. COMPARACIONES CRÍTICAS Nos permitimos considerar el criterio por el cual las teorías de DM deberían ser juzgadas. En la tabla 2 se resúmen aquellas consideradas por Laland, Mc Manus y nosotros. Criterios sugeridos para juzgar las teorías de la DM 1. Es apta para explicar los diferentes promedios de DM izquierda si la descendencia es de padres diestro-diestro, diestro-zurdo y zurdo-zurdo. 2. Es apta para explicar la observación que los ñ adoptados se parecen a sus padres biológicos, y no a sus padres adoptivos. 3. Es apta para explicar los parecidos mínimos entre pares de hermanos y gemelos homocigotos. 4. Es apta para explicar las evidencias de que hasta qué grado la DM puede ser heredada. 5. Es apta para explicar la mayor concordancia de DM entre la madre y su descendiente que la DM entre el padre y su descendiente 6. Es apta para explicar las diferencias de sexo en la DM. 7. Es apta para explicar el hecho que todas las culturas humanas son diestras 8. Es apta para explicar el hecho que no hay variación en la DM a través de la cultura. 9. Es apta para explicar los procesos que subyacen en la DM humana. 10. Es apta para explicar para conocer otras relaciones de la DM con :otras lateralizaciones funcionales y anatómicas, marcadores de la variaciones en el crecimiento temprano. 11. Es apta para explicar la evidencia de fertilidad reducida en los zurdos 12. Es apta para hacer todo lo de arriba con supuestos mínimos ad hoc, generalizaciones y máximas mezquindades El mayor logro de Annett y Mc Manus es su capacida de patrones de modelos efectivos de parecido familiar de preferencia manual. Ellos no pueden explicar por evidencias que el grado de DM puede ser heredado, ni tampoco explican los recientes informes vinculados a alelos de HLA en la DM izquierda o un gen cromósomico ligado al X el grado de DM. Más aún ellos no pueden explicar la relaciones de DM demostrada

18
en estudios que sostienen en el esquema del DI. Más aún ellos no proveen explicaciones convincentes de cómo la variabilidad genética es mantenida. El beneficio heterozigota es invocado, pero no demostrado. Si, como estas teorías postulan, un gen único tiene un efecto principal en la DM, la gran vinculación de estudios debería detectarlos. El modelo genético cultural de Laland también en efecto modelo de informes familiar, y lo hace sin invocar un gen hipotético, único a los humanos, que confiere variación direccional en la asimetría cerebral. Pero tiene 2 importantes defectos. La sugerencia que el parecido familiar es toda de origen socio-cultural, y una hipótesis reñida con los informes de ñ adoptados y las observaciones que u grado de DM es heredada. Esto también no es capaz de explicar muchas de las asociaciones de DM vistas arriba. El modelo de DI comparte los beneficios del modelo de genética-cultural, pero es capaz de explicar la riqueza de relaciones demostradas de DM. Creemos que el modelo de DI es también mas mezquino , en 2 diferentes formas. La 1ra factores no determinantes de las diferencias individuales en asimetría están invocadas fuera de aquellas descriptas en otras especies. Los mismos factores que conducen a la AF en otras especies conducen hasta un punto a la variación de la asimetría cerebral y de la mano atípica en humanos. Qué es diferente en nuestra especie , y qué es compartido por todos los miembros de nuestra especie, es un plan único de desarrollo neural que culmina en algunas de nuestras asimetrías. 2) Este es un enfoque que puede ser extendido a otras asimetrías, y que no requiere de supuestos separados, determinantes genéticos múltiples de diferencias individuales en otros sistemas asimétricos (ej juntar las manos ) . UNA PERSPECTIVA EVOLUTIVA EN DM En el comienzo de este capítulo sugerimos que un entendimiento de la DM debería proveer alguna información sobre la naturaleza humana, la naturaleza de nuestro cerebro y sus habilidades únicas. En esta sección esperamos satisfacer esa promesa con un análisis de las importantes implicaciones tanto del modelo de DI y el modelo genético-cultural. Cada una sugiere que todos los humanos comparten un mismo plan genético básico que, entre otras cosas, culmina en le grado de especie común de DM derecha. Si en efecto el plan es universal, luego pude servir mejor a una función adaptativa. Esto es, puede haber sido suficientemente adaptativo en ambientes humanos ancestrales que seleccionan efectivamente eliminando diseños de otras fenotipos. ¿En qué momento de la evolución de los primates apareció la 1ra DM? ¿Es el diseño para la DM o para alguna propiedad más fundamental del sistema nervioso? ¿Cuál es exactamente la naturaleza de los beneficios provistos por el diseño subyacente a la destreza manual?. CONSIDERACIONES FILOGENETICAS E HISTORICAS Una clave importante de cuándo apareció por 1ra vez la DM en el cerebro d primates ha surgido de recientes estudios de primates no humanos. Nos enfocamos en los chimpancés, nuestra relación relativamente estrecha con los primates, con quienes compartimos el 98% aproximadamente de nuestro material genético. La lateralidad no es evidente en tareas simples de alcanzar o en el uso de herramientas . Sin embargo, la lateralidad diestra consistente ha sido descripta en tareas manuales más complejas y la tarea de comer con las 2 manos. Por lo tanto, la tendencia direccional hacia la DM puede estar presente en los chimpancés, pero no demuestran ningún grado de asimetría de población ni tampoco ninguna clase de preferencia de tareas cruzadas vista en humanos. Con respecto a esto es interesante destacar 2 recientes estudios que documentaron que hay una asimetría hacia a la izquierda del planum temporale en chimpancés , equivalente en magnitud a lo observado en humanos. Tradicionalmente, el mayor tamaño del planum izquierdo se pensó que representa parte del sustrato del hemisferio izquierdo dominante para el lenguaje. Adicionalmente, asimetrías en el área de Broca análogas a aquellas vista en humanos han sido documentadas estar presentes en 3 grandes especies de simios.. La emergencia de estas asimetrías en nuestra estrecha relación sugiere una presión relevante selectiva estaban no relacionado al lenguaje. Por lo tanto las asimetrías anatómicas corticales notables probablemente surgen de un ancestro común al menos 5 millones de años atrás, previo al desarrollo tanto de la DM y del lenguaje. Eso es potencialmente una clave importante del origen de nuestras asimetrías, una de las cuales es la DM que es un ejemplo funcional, sugiriendo que ellos pueden servir a la intención cognitiva general, quizás relacionada a las necesidades organizativas de un gran cerebro. Annett sugiere que el alelo rs+ conduce tanto a una asimetría del planum y a una asimetría en el sustrato de la DM, esto ahora parece ser bastante improbable. El aumento de la lateralidad vista en humanos debería, por supuesto, no haber sido seleccionada en forma directa. Un reciente estudio comparativo demostró que el incremento importante en el tamaño cerebral visto

19
en el mono(23cm3) y en los humanos(1300cm3) ha sido acompañado por una disminución en el tamaño relativo tanto del cuerpo calloso y de la comisura anterior. Esto es, los cerebros de primates están caracterizados por una menor integración hemisférica funcional. La misma tendencia se notó en las especies cetáceas, sugiriendo una menor conectividad, y una mayor independencia hemisférica, puede ser un diseño característico consistentemente asociado con un mayor volumen cerebral. Rinlling sugiere que” la función de lateralidad puede ser una propiedad surgida acompañando un agrandamiento en la evolución de los primates”. La asimetría anatómica aumentada también acompaña un aumento en el volumen dentro de la línea de los primates. Hopkins analizaron la RMN de 45 primates. La asimetría hacia la izquierda d la petalia (especialmente occipital) predice un mayor tamaño cerebral total y un tamaño menor del cuerpo calloso. Más, especies de primates con preferencia manual derecha tiene un área callosa menor (en relación al volumen cerebral). El reanálisis de estos datos también sugieren una asociación entre tamaño cerebral y asimetría cerebral en primates. Le May publica informes sobre importantes tipos de asimetría en diversas especies de primates, si bien el n° de cerebros examinados de cada especie fue bastante pequeña. Ella proveyó información en la presencia de asimetrías, tanto hacia la derecha como hacia la izquierda, a estas características: la longitud de los hemisferios cerebrales, la altura de la Cisura de Silvio, la petalia occipital y el ancho del polo occipital. Calculamos el promedio de asimetrías observadas por individuo de cada especie, sin tener en cuenta la dirección de la asimetría. El dato del volumen cerebral fue obtenido de Rilling. Calculamos la raíz cuadrada del volumen cerebral y relacionamos ese valor con la medida global de asimetría. Una relación fuerte fue observada en las especies r = 0,91, p menor 0,001, como se muestra en la figura 5. Por lo tanto ,parece ser que en primates en el aumento del volumen cerebral está asociado con una mayor asimetría y menor conectividad cortical. Como los cerebros más grandes son aptos a ser más inteligentes, la asimetría puede ser un diseño de características vinculadas con la inteligencia, Consistente con nuevos hallazgos de asimetrías anatómicas en otros primates, y la sugerencia que la asimetría puede representar un diseño característico de cerebros grandes, la DM parece haber surgido bastante en la línea hominide, junto con Orebro grandes. Se examinaron las herramientas de piedra del homo habilis y observaron que la forma en que obtenían dichas herramientas indicaban que probablemente lo hacían individuos diestros. Corballis citó otros ejemplos sugiriendo un surgimiento temprano de la DM diestra, incluyendo estudios que indicaban grandes evidencias del desgasten el lado derecho de las herramientas de piedra. Mientras estos estudios sugieren una aparición temprana de la DM derecha, ellos son insuficientes para establecer si la incidencia de la DM derecha fue similar a la línea actual. Las teorías genéticas de Annett y Mc Manus establecen que la DM derecha surge secundariamente a la dominancia del hemisferio izquierdo para el lenguaje y por lo tanto, el orden de aparición de estos 2 rasgos en nuestros antecesores es de gran interés. Ha sido muy debatido de cuando exactamente surgió el 1er lenguaje humano, pero parece probablemente que el lenguaje moderno es de origen más reciente que la DM: Bickerton argumentó que el “protolenguaje”, difiere significativamente de nuestra propia complejidad gramatical y fonémica, más probablemente surge del homo erectus, aproximadamente hace 16 millones de años. El lenguaje moderno es mucho más que una comunicación social, proveyendo a un desarrollo de una cultura rica en ideas y conocimientos, así como sirviendo de vehículo para simbolismos e pensamientos abstractos. Por lo tanto muchas escuelas argumentan que el surgimiento del lenguaje real debería ser coincidente con el surgimiento de muchos productos conferidos por el lenguaje. Como mucho uno puede inferir desde la diversidad y complejidad de herramientas de piedra , que fue un cambio no significativo desde el tiempo del h. erectus(hace 14 millones de años) hasta aproximadamente 100000años atrás. Las herramientas de piedra del h erectus fueron remarcadamente estables durante ese período de tiempo. Esto sugiere la ausencia del lenguaje moderno con su aptitud para registrar el desarrollo comunicativo histórico y por lo tanto facilitar la mejora tecnológica. White señaló el desarrollo importante en el p surgió período paleolítico que coincidiría con el surgimiento funcional del lenguaje moderno: expresiones artísticas en el arte de las cavernas y los adornos corporales, un aumento en el desarrollo de nuevas herramientas y el n° de materiales usados para fabricar herramientas.,un aumento de las comunidades y la variación regional cultural. Por lo tanto parece que la emergencia de las asimetrías cerebrales anatómicas y las de la DM anteceden la lenguaje humano actual. Si es así, hay importantes implicaciones a teorías de los orígenes de la DM. Los beneficios asociados con un cerebro asimétrico, del cual la DM es una manifestación obvia, podría no serlo para el lenguaje actual, la idea que uno puede argumentar que la aparición temprana de la asimetría está asociada con el desarrollo de un protolenguaje. La evidencia revisada arriba, sin embargo, nos persuade que el patrón humano de asimetría sirve a una propuesta más general que la del lenguaje. Creemos que ninguna DM derecha y tampoco ningún tipo de habilidad motriz fue el fenotipo selecto activamente sobre el eón de la prehistoria humana. Más, la DM derecha es el producto de presión selectiva a otros fenotipos. Sugerimos que

20
el patrón humano del desarrollo de la asimetría cerebral representa un diseño facilitando el desarrollo de la eficiencia computacional o inteligencia. DISEÑO CEREBRAL ADAPTATIVO Y ASIMETRIA Asumimos que la función cerebral es masivamente paralela en arquitectura, y que, a lo mejor, sólo un “protomapa” impreciso existe para especificar la localización cortical de una función. Las influencias genéticas en el desarrollo cortical están probablemente para codificar clases de estrategias de “autoorganización”, más que el destino de células específicas o un grupo celular. Pase lo que pase estas estrategias pueden estar, y necesitan consiguientemente un diseño que es bastante eficiente y flexible para trabajar efectivamente e tipos de ambientes en los cuales los humanos deben ser esperados a desarrollar( ej un ambiente lingüístico inglés o español). Algunas claves como el de características del diseño permitiendo la eficiencia puede ser recogidos de los principios de computadoras funcionando en paralelo. Mark notó que la modularidad (en débil sentido) es un principio de diseño eficiente, tanto para cerebros como para computadoras permitiendo una depuración eficiente. Nelson describe 2 fuentes fundamentales de ineficiencia computacional tanto en computadoras en paralelo y cerebros. Una carga en desequilibrio se refiere a 2 cómo uniformemente la carga de trabajo computacional es distribuida entre los procesadores disponibles. Mientras la velocidad de la computación paralela está limitada por la velocidad del procesador más lento, excesivamente cargando incluso a un simple procesador puede dramáticamente disminuir la perfomance global. El 2do factor la comunicación elevada es “el costo de información comunicativa entre los procesadores… 1riamente asociados con el costo del tiempo para intercambiar información entre los espacios físicos de los procesadores obtenida por conecciones entre los procesadores. Poniendo las cualidades de la modularidad y el costo elevado comunicacional y la carga en desequilibrio, surgen 2 principios diferentes de diseño cortical eficiente: los módulos podrían estar distribuidos equitativamente a lo largo de los 2 hemisferios (limitando la carga en desequilibro) y “) módulos que interactúan frecuentemente podría desarrollarse en una proximidad física (limitando la elevada comunicación). Este diseño puede no estar solidamente conectado y, con otros aspectos del desarrollo cortical es probablemente alcanzado a través de una estrategia de desarrollo autoorganizativo. Sugerimos que el plan de neurodesarrollo para implementar estos principios culmina en el patrón modal de la lateralidad humana. El mecanismo para implementar este plan se postula que es una secuencia alternante de promedios de crecimiento hemisférico asimétrico. El mejor informe para tal patrón “dialéctico” del desarrollo proviene del estudio EEG en niños, la idea es también evidenciar el promedio de crecimiento prenatal asimétrico del giro cortical y el crecimiento dendrítico asimétrico. Como mucho, la duración del comienzo del crecimiento es aproximadamente igual para cada hemisferio, ese plan de desarrollo se distribuye equitativamente sobre los 2 hemisferios. Esto disminuye la carga en desequilibrio. Aquella porción de la corteza que se desarrolla más rápidamente en un momento determinado estará influenciada al máximo por inputs ambientales y conductas prevalentes formando patrones de actividad cortical. Debido a que las habilidades en la vida son aprendidas durante la infancia son un tanto específicas de la edad (lenguaje versus relaciones con la edad), aquellas módulos relacionados a un grupo de habilidades dadas están probablemente desarrolladas en proximidad física. Eso minimiza la excesiva comunicación. Thatcher destaca que sólo porciones particulares de cada hemisferio pueden mostrar un comienzo de crecimiento durante una fase dada (derecho más rápido o izquierdo más rápido). Por lo tanto el tiempo durante el cual una asimetría dad es establecida difiere de acuerdo a la trayectoria desarrollada del sustrato cortical específico. La dirección de una asimetría funcional dada está determinada por la fase (ej aquel hemisferio que se desarrolla más rápidamente) Asumimos que las asimetrías direccional amplia en especies reflejan los posibles eventos de nuestra historia filogenético distante. Esto es, los beneficios no computacionales a asimetrías direccionales comunes en la especie, tales como dominancia del hemisferio izquierdo para el lenguaje, es comparado como una imagen en espejo. Esa perspectiva sugiere que el neurodesarrollo universal propuesto, el cual culmina en la típica lateralidad cerebral, fue seleccionado como para asegurar una eficiencia computacional. Otra estrategia puede ser más deseable en especies en las cuales la adaptación postnatal a circunstancias ambientales particulares es menos importante en nosotros mismos. 2 siguientes predicciones:1) la mayor lateralidad de a la función a través de las especies podría estar asociada con una mayor eficiencia computacional. Esta hipótesis es difícil de ser testeada directamente en gran parte porque se está lejos de cómo exactamente la eficiencia computacional podría estar operando en especies cruzadas en comparación.

21
2) variaciones de asimetrías cerebrales comunes podrían predecir una eficiencia computacional más reducida dentro de los miembros de especies de Orebro grande. Para hacerlo más simple, consideramos a la “inteligencia” como una autoridad de eficiencia computacional. La posesión de estos patrones humanos de crecimiento deberían ser más importante a inteligencia que variaciones direccional específicas en la lateralidad. Un individuo con asimetría cognitiva y manual exactamente lo opuesto de la norma de la población no obstante logra un diseño cerebral limitando la sobrecarga comunicacional y la carga en desequilibrio debido a un crecimiento asimétrico de desarrollo. En tales casos un retraso temprano en el crecimiento probablemente conduce a una “fase inversa” en que el patrón dialéctico, pero los módulos aún se desarrollan por igual a través de los hemisferios y los módulos funcionalmente relacionados están para estar en el mismo hemisferio. En cambio, en el individuo sin ese patrón de crecimiento especial, ej una ausencia en alguna asimetría direccional, quien puede sufrir de una insuficiencia cognitiva. En términos de DM, es el individuo “ambiguo”, sin una diferencia en la preferencia o habilidad a través de las tareas, podrían sufrir dificultades cognitivas .En relación a esto es importante notar que los variados trastornos del neurodesarrollo frecuentemente se vinculan con variaciones en la destreza manual y son más característicos los individuos con DM “ambigua” que la DM izquierda per se. Esto es verdadero para el retraso mental, la esquizofrenia y dislexia demostrando que un CI relativamente más bajo fue encontrado en el de “hemisferio indeciso”o habilidades manuales iguales en la derecha e izquierda, en individuos normales. Si el DI conduce a una lateralidad atípica, podría también llevar a un menor funcionamiento cognitivo eficiente. Sustentando esta hipótesis, una mayor asimetría fluctuante corporal, la cual hemos vinculado con asimetrías cebarles atípicas, predice una baja inteligencia fluida. Basados en un modelo matemático que detalla la fuerte relación entre la asimetría fluctuante y los rasgos subyacentes de desarrollo inestable, del 17 al 50% de la variancia heredada en la inteligencia está relacionada con el DI. Uno debería sospechar ue las mediciones de asimetría cerebral atípica podría mostrar por igual una fuerte relación con la ineligencia que las mediciones de l asimetría fluctuante corporal. CONCLUSIONES La DM y las asimetrías cerebrales funcionales y anatómicas representan atributos humanos que nos distinguen cualitativamente de otros primates. Una perspectiva biológica de la DM puede ayudarnos a esclarecer aspectos fundamentales de la naturaleza humana y su historia evolutiva. Hay todavía mucho que aprender y el fundamento empírico está lleno de vacíos e inconsistencias Harris enfatiza que el progreso en el entendimiento de la destreza manual debería depender de un análisis adicional del fenotipo, una muestra de gran tamaño, técnicas de neuroimagen y una aguda observación a las causa de los cambios del desarrollo. Además , en los próximos los desarrollos genéticos y psicológicos seguramente tendrán un impacto en este campo.