dinámica molecular de proteínas modelado y simulación ... · dinámica molecular – unsam –...
TRANSCRIPT

Dinámica Molecular de ProteínasDinámica Molecular de ProteínasModelado y Simulación ComputacionalModelado y Simulación Computacional
Profesores: Eliana K. Asciutto & Ignacio J. General2do cuatrimestre 2018
Escuela de Ciencia y TecnologíaUNSAM

Dinámica Molecular de ProteínasDinámica Molecular de ProteínasModelado y Simulación ComputacionalModelado y Simulación Computacional
Introducción al modelado de proteínasIntroducción al modelado de proteínas

3
Aminoácidos
Dinámica Molecular – UNSAM – 2018
Cα C
O
O
N
H
H
H
R
Cα C
O
N
H
H
H
R
Cα C
O
O
N
H H
R
Enlace peptídico
Cadena lateral
Esqueleto
H
Grupo amino Grupo carboxilo
P + P PP + H2O
Proteínas --> polímeros de 20 aminoácidos naturales
Terminal N o amino
Terminal C o carboxilo

4
Aminoácidos
Dinámica Molecular – UNSAM – 2018
C
O
N
H
El enlace peptídico es un enlace parcialmente doble (híbrido de resonancia)
C y N están sp2 hibridados:
C N
H
..
..
..O
......
+1
-1
C N
O
H
C N
O
HVisto de arriba de costado
sp2
pz
Unión σUnión σ
Unión πUnión π

5
Aminoácidos
Dinámica Molecular – UNSAM – 2018
Como consecuencia del (parcial) doble enlace peptídico:
El enlace peptídico es plano y rígidoEl enlace peptídico es plano y rígidoTiene menor longitud C-N (1,33 Tiene menor longitud C-N (1,33 ÅÅ) que una unión simple C-N (1,45 ) que una unión simple C-N (1,45 ÅÅ))
Tiene mayor longitud C-O que un carbonilo típicoTiene mayor longitud C-O que un carbonilo típico
O
H
R
Cα
N
H
H
R
Cα
C

6
By Dan Cojocari · · [CC BY-SA 3.0 (http://creativecommons.org/licenses/by-sa/3.0) or ✉ ✍GFDL (http://www.gnu.org/copyleft/fdl.html)], via Wikimedia Commons
Cargados
Neutros, polares Otros
Hidrofóbicos
(family vw)

7
Aminoácidos - Estructura 3D
Dinámica Molecular – UNSAM – 2018
C
N
Cα
Cφφ: CNC
αC
ψ: NCαCN
ω: CαCNC
α
Ángulos diedros del esqueleto:
NN
Cα
C
ψ
N
Cα
Cα
C
ω
χ1: NC
αC
βC
γ
χ2: C
αC
βC
γC
δ
...
Ángulos diedros de las cadenas laterales:
Cα C
O
O
N
Cβ
Cγ
χ1
φ
casi siempre ~ 1800 (trans),excepto en Pro ~ 0 (cis)

8
Aminoácidos - Estructura 3D
Dinámica Molecular – UNSAM – 2018
Ángulos diedros del esqueleto:
Biochemistry. 7Th edition ©2012 W. H. Freeman and Company
N
Cα C
α
C
ω=180°
NC
α Cα
Cω
ω=180°
ω
Trans es usualmente más estable: repulsión estérica entre cadenas lateralesEl ΔE(trans-cis) es mucho mayor para Pro ==> tarda mucho en pasar a trans
Una proteína con Pro en cis retarda mucho su plegamiento, exhibiendo dos velocidades de plegamiento

9
Aminoácidos – Protonación
Dinámica Molecular – UNSAM – 2018
[H+]agua
= 10-7 → pHagua
= 7
pH=−log10 [H+] Mide la concentración de H+ en solución
jugo de limón
jabón
sangre
1
5
9
13
café
ácido estomacal
destapa caños
pH
ácidoácido
básicobásico

10
Aminoácidos – Protonación
Dinámica Molecular – UNSAM – 2018
HA⇔H++ A−
Reacción de deprotonación
K=[H+
][ A−]
[HA ]
pK a=−log K
Constante de equilibrio
Definición de pKa
Si pKa= pH: −log
[H +][ A−
]
[HA ]=−log [H+
] ⇒ [ A−]=[HA ]
Si pKa= pH+1:−log
[H+][ A−
]
[HA ]=−log [H+
]+log10=−log[H+
]
10
⇒ [ A−]=
[HA ]
10o [HA ]=10 [H+
]
Si pKa= pH-1:
⋯ ⇒ [HA ]=[H +]/10
o [H+]=[HA ]

11
Aminoácidos – Protonación
Dinámica Molecular – UNSAM – 2018
Entonces:
pka = pH → la mitad del aa está deprotonadopka = pH → la mitad del aa está deprotonado
pka > pH → el aa está mayormente protonadopka > pH → el aa está mayormente protonado
pka < pH → el aa está mayormente deprotonadopka < pH → el aa está mayormente deprotonado
[HA ]=10 [H +]
[HA ]=[H+]/10
[H +]=[HA ]

12
Aminoácidos – Protonación
Dinámica Molecular – UNSAM – 2018
Ejemplo) Histidina
Cα C
O
ON
H
H
H
HC
C CN H
CCNH
+
H+
Cα C
O
ON
H
H
H
C
C CN H
CCNH
+
H+
–
Cα C
O
ON
H
H
H
C
C CN
CCNH
H+
Cα C
O
ON
H
H
H
C
C CN
CCNH
––
A BC D
pkaamino = 9.2
pKacarboxilo = 1.8
pKalateral = 6.0
¿Qué estructura tendrá?
pH < 1.8: A1.8 < pH < 6.0: B6.0 < pH < 9.2: C9.2 < pH : D
A medida que el pH aumenta (el solvente “quiere” mas [H+]), HIS dona sus H.

13
Aminoácidos – Protonación
Dinámica Molecular – UNSAM – 2018
Punto isoeléctrico (PI):Punto isoeléctrico (PI):
Es el pH al cual la molécula tiene carga eléctrica nula.Se puede calcular como el promedio de los pKa alrededor del caso neutral.
Para el HIS: PI = (6.0+9.2)/2 = 7.6
PH > PI molécula con carga negativaPH < PI molécula con carga positiva

14
Aminoácidos – Gráfico de Ramachandran
Dinámica Molecular – UNSAM – 2018
Dado el volumen que ocupan los grupos R, el fenómeno de choques estéricos y la tendencia de las cadenas laterales a interactuar, es de esperar que exista un sesgo en φ y ψ.
By Dcrjsr (Own work) [CC BY 3.0 (http://creativecommons.org/licenses/by/3.0)], via Wikimedia Commons
Todos los aa, excepto Pro y Gly
Pro (muy rígido) Gly (muy flexible)
15
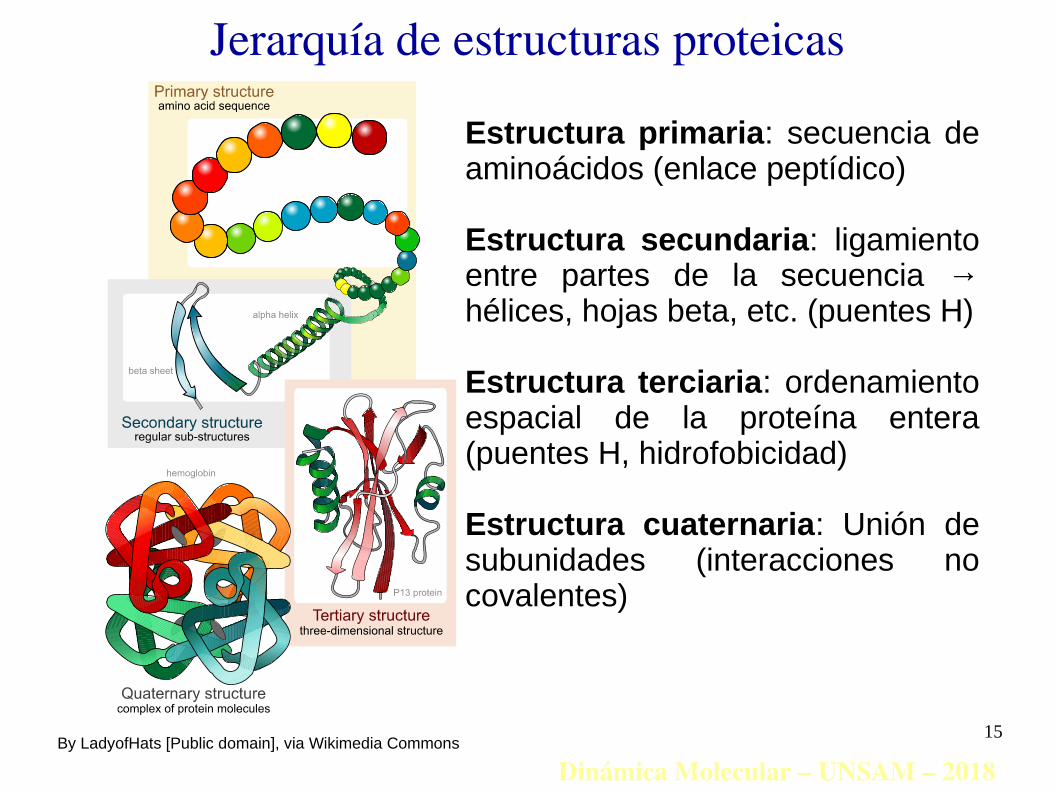
Jerarquía de estructuras proteicas
Dinámica Molecular – UNSAM – 2018 By LadyofHats [Public domain], via Wikimedia Commons
Estructura primaria: secuencia de aminoácidos (enlace peptídico)
Estructura secundaria: ligamiento entre partes de la secuencia → hélices, hojas beta, etc. (puentes H)
Estructura terciaria: ordenamiento espacial de la proteína entera (puentes H, hidrofobicidad)
Estructura cuaternaria: Unión de subunidades (interacciones no covalentes)

16
Jerarquía de estructuras proteicas
Dinámica Molecular – UNSAM – 2018
Hélice alfa: Hélice alfa:
● Puentes H entre O de un aa en la posición i y NH del aa en la posición i+4
● Paso de la hélice ~ 5.4 Å● Pro y Gly la desestabilizan
Puente H

17
Jerarquía de estructuras proteicas
Dinámica Molecular – UNSAM – 2018
Lamina beta:Lamina beta:
● Esqueleto en zig-zag● Cadenas adyacentes (// o anti-//) formando una lamina● Cadenas estabilizadas por puentes H
Puentes H
2 cadenas paralelas
2 cadenas anti-paralelas

18
Jerarquía de estructuras proteicas
Dinámica Molecular – UNSAM – 2017
Giro beta:Giro beta:
● Puentes H entre O de un aa en la posición i y NH del aa en la posición i+3
● Cambia la dirección de la cadena● Conecta hélices/láminas● Pro y Gly lo estabilizan
C
C
N
C
HN C
C
N
C
H
H H
O
HO
H
O
H
ß turn: Type I ß turn: Type II
C
C
N
C C NH
C
C
N
CH
O H
OH
O H
H
H
By Muskid (Own work) [CC BY-SA 3.0 (http://creativecommons.org/licenses/by-sa/3.0)], via Wikimedia Commons
Puente H

19
Jerarquía de estructuras proteicas
Dinámica Molecular – UNSAM – 2017
Alfa, beta:Alfa, beta:
By CNX OpenStax [CC BY 4.0 (http://creativecommons.org/licenses/by/4.0)], via Wikimedia Commons
Giro β
Hélice α
Lamina β

20
Jerarquía de estructuras proteicas
Dinámica Molecular – UNSAM – 2017
Frecuencias relativas de aa en estructuras secundariasFrecuencias relativas de aa en estructuras secundarias
"Biochemistry" 2nd ed. Garrett, R.H. and Grisham, C.M. Saunders College Publishing. 1999.

21
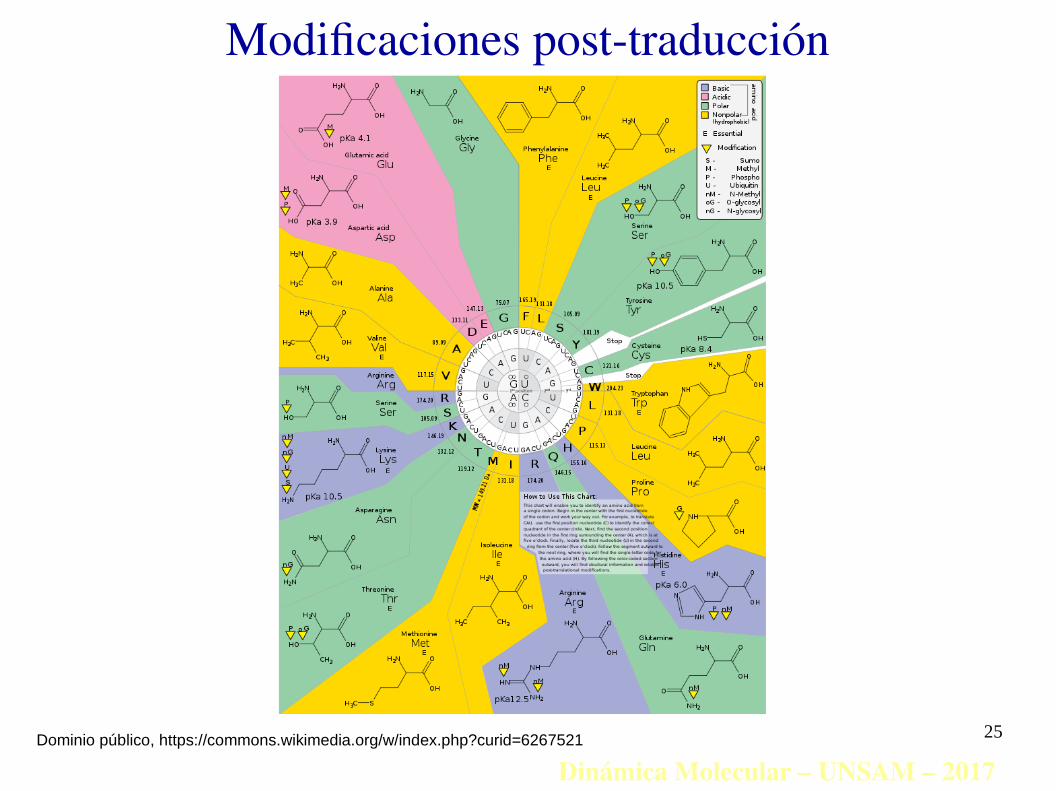
Modificaciones post-traducción
Dinámica Molecular – UNSAM – 2017
Glucosilación:Glucosilación:● Es la reacción por la cual un carbohidrato (azúcar donante) es
adicionado a un grupo funcional de una proteína (aceptor).

22
Modificaciones post-traducción
Dinámica Molecular – UNSAM – 2017
Fosforilación:Fosforilación:● Es el proceso por el cual un grupo fosfato (PO4) es agregado a
una proteína, en forma covalente. ● Los aminoácidos usualmente fosforilados (defosforilados), por
acción de una quinasa (fosfatasa), son SER, THR, TYR y HIS. ● El ejemplo más importante en biología es la fosforilación del
ADP para producir ATP.

23
Modificaciones post-traducción
Dinámica Molecular – UNSAM – 2017
Acetilación:Acetilación:● Es el proceso por el cual un grupo acetilo (CH3CO) es
agregado a una proteína, en forma covalente. ● Las proteínas son típicamente acetiladas en las Lisinas. ● Las enzimas que catalizan estos procesos son:
acetiltransferasa y deacetilasa.

24
Modificaciones post-traducción
Dinámica Molecular – UNSAM – 2017
Enlace (puente) disulfuro:Enlace (puente) disulfuro:● Son enlaces formados entre dos átomos de azufre. ● En las proteínas estos enlaces se forman entre los grupos tiol
de las Cisteinas (no así entre los S de las Metioninas).● La estructura del enlace disulfuro se puede describir por su
ángulo diedro χss entre átomos Cβ–S–S–Cβ, que usualmente se aproxima a ±90°.
● De Jü - Trabajo propio, CC0, https://commons.wikimedia.org/w/index.php?curid=24825672
Ejemplo de enlace SS en una proteína. Importantes para determinar la estructura terciaria de la proteína. *
25
Modificaciones post-traducción
Dinámica Molecular – UNSAM – 2017 Dominio público, https://commons.wikimedia.org/w/index.php?curid=6267521

26
Plegamiento de proteínas
Dinámica Molecular – UNSAM – 2017
Plegamiento de proteínas:Plegamiento de proteínas:● Es el proceso que le da una forma especifica a una serie de aa
sin forma inicial (cadena ==> hélice). Se dice que la proteína llega a su estado nativo, su conformación más estable.
● Dicho proceso puede ser ayudado por moléculas chaperonaschaperonas
Desnaturalización de proteínas:Desnaturalización de proteínas:● Es el proceso por el cual la proteína en estado nativo pierde su
estructura 3D.● Generalmente la estructura es fundamental para la función
==> desnaturalización = perdida de función● Se puede producir por cambios de T, ataque químico
(detergentes, sales, ácidos), ataque mecánico (agitación), etc.

27
¿Por qué se pliegan las proteínas?
Dinámica Molecular – UNSAM – 2017
Recordar que la estabilidad de un sistema físico a presión (volumen) constante viene dado por su energía libre de Gibbs (Helmholtz):
ΔG=Δ H−T Δ S
El sistema tenderá a minimizar su entalpía minimizar su entalpía (energía)(energía) y maximizar su entropía
i.e., aumentar el desordenaumentar el desorden
(Δ F=Δ E−T Δ S)

28
¿Por qué se pliegan las proteínas?
Dinámica Molecular – UNSAM – 2017
+-
Grupos atómicos no-polares→hidrofóbicos.Las aguas no se sienten atraídas al grupo, quedando desorganizadas, > S> S.
Grupos atómicos polares→hidrofílicos (el agua es polar). Las aguas apuntan al grupo y se acercan, quedando más ordenadas, < S< S.
Fuerzas hidrofílicashidrofílicas vs hidrofóbicashidrofóbicas (amor vs miedo al agua)
El agua es una molécula polar (H2+O-). Por lo tanto, las
moléculas polares tenderán a unirse a ella.

29
¿Por qué se pliegan las proteínas?
Dinámica Molecular – UNSAM – 2017
Ej) Sub-unidad de CaMKII
¿Por qué CaMKII no está más extendida?
Comparemos las dos posibilidades (compacta vs extendida). Para ello supongamos una superficie h-fi (azul) y un nucleo h-fo (verde)
núcleo h-fo
Las aguas son atraídas a la superficie h-fi→S
H2O decrece (más orden)
Lo mismo, pero ahora entran más aguas→S
H2O decrece aun más
Las formas globulares (izquierda) permiten maximizar S, escondiendo a los grupos hidrofóbicos de la proteína.
superf. h-fi

30
¿Por qué se pliegan las proteínas?
Dinámica Molecular – UNSAM – 2017
Entonces, es más favorable una proteína h-fo globular que una extendida. Pero, ¿es favorable diluir una proteína (o un grupo h-fo)? ¿O la proteína preferiría no encontrarse en agua?
Datos experimentales* de disolución de cadena h-fo (butanol, pentanol):
*Atkins, Physical Chemistry for Life Sciences.
ΔG0(kJ/mol) ΔH0(kJ/mol) ΔS0(J/mol.K)
CH3CH
2CH
2CH
2OH -10 +9 +65
CH3CH
2CH
2CH
2CH
2OH -13 +8 +72
Las cadenas hidrofóbicas largas se disuelven más fácilmente.
¿Por qué?¿Por qué?
31
¿Por qué se pliegan las proteínas?
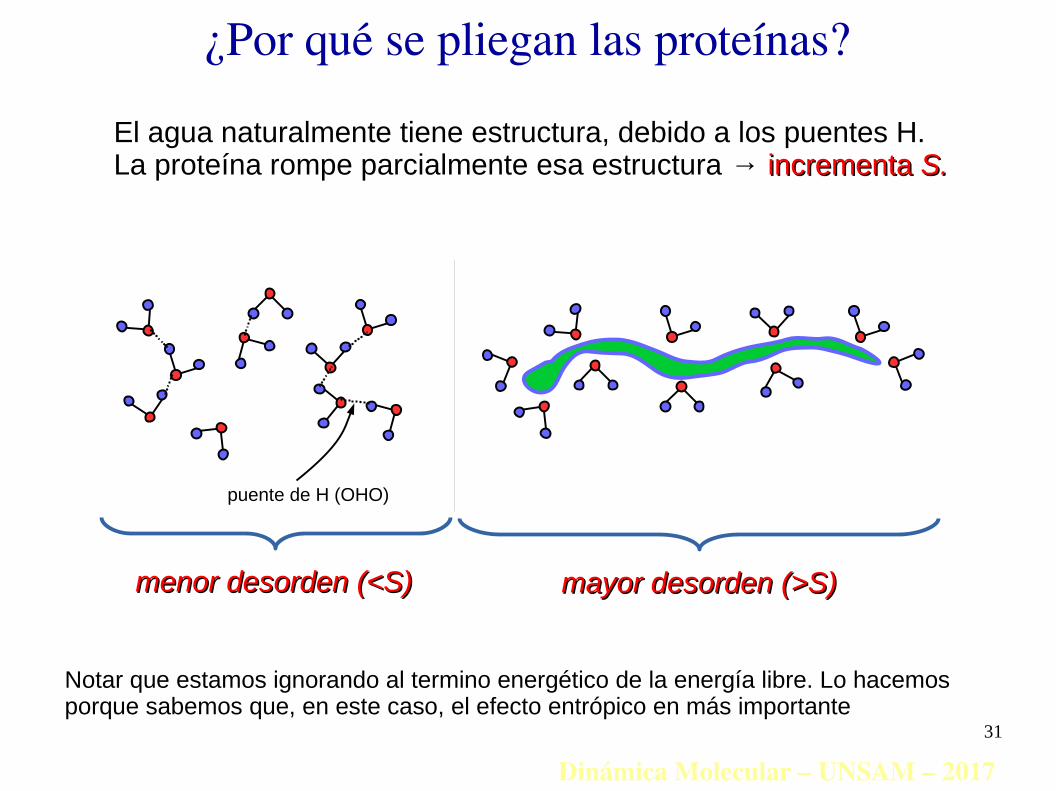
Dinámica Molecular – UNSAM – 2017
puente de H (OHO)
El agua naturalmente tiene estructura, debido a los puentes H.La proteína rompe parcialmente esa estructura → incrementaincrementa S.S.
mayor desorden (>S)mayor desorden (>S)menor desorden (<S)menor desorden (<S)
Notar que estamos ignorando al termino energético de la energía libre. Lo hacemos porque sabemos que, en este caso, el efecto entrópico en más importante

32
¿Por qué se pliegan las proteínas?
Dinámica Molecular – UNSAM – 2017
Usando estos conceptos:
1) Mutar un aminoácido de alguna proteína a glicina, ¿debería favorecer su plegamiento (P) o su desnaturalización (D)?
El grafico de Ramachandran de la glicina muestra que es muy flexible. En una zona interna de una proteína P no va a poder moverse, debido a las restricciones del empaquetado. Pero en una proteína D, si va a poder hacerlo, aumentando su S→→favorece al estado Dfavorece al estado D
2) Incluir enlaces disulfuro en una proteína, ¿debería favorecer su plegamiento (P) o su desnaturalización (D)?
Los enlaces ponen restricciones al movimiento de la proteína, tanto en el caso P como D. Pero el estado P ya esta restringido por el empaquetamiento. Por eso, es el estado D el que pierde mas entropía→→favorece al estado Pfavorece al estado P