contribución al conocimiento de los opiliones de la … · resumen de la tesis presentada para...
TRANSCRIPT

UNIVERSIDAD DE BARCELONA
SECRETARIADO DE PUBLICACIONES.
Dlltflmm'll\1. lEVl HERBERT W. LEVI
INTERCAMBIO CIENTÍFICO Y EXTENSIÓN UNIVERSITARIA
Contribución al conocimiento de los Opiliones de la fauna ibérica.
Estudio de los subórdenes Laniatores y Palpatores (pars.)
RESUMEN DE LA TESIS PRESENTADA PARA ASPIRAR AL GRADO DE DOCTOR EN CIENCIAS
POR
MARIA RAMBLA CASTELLS
BARCELONA - 1973

UNIVERSIDAD DE BARCELONA
Facultad de CIENCIAS (Sección de Biológicas)
Tesis doctoral del Dr. MARIA RAMBLA CASTELLS
TEMA:
CONTRIBUCION AL CONOCIMIENTO DE LOS OPILIONES DE LA FAUNA IBERICA ESTUDIO DE LOS SUBORDENES LANIATORES Y PALPATORES (PARS.)
Presidente:
Vocales:
Director:
TRIBUNAL DE TESIS
Dr. D. SANTIAGO ALCOBE NOGUER Catedrático de Antropología Facultad de Ciencias Universidad de Barcelona
Dr. D. ANTONIO PREVOSTI PELEGRIN Catedrático de Genética Facultad de Ciencias Universidad de Barcelona
Dr D. JAIME GALLEGO BERENGUER Catedrático de Parasitología Facultad de Farmacia Universidad de Barcelona
Dr. D. ARTURO VALLS MEDINA Profesor Agregado de Antropología Facultad de Ciencias Universidad de Barcelona
Dr. D. ENRIQUE GADEA BUISAN Catedrático de Zoología (Inv. no Artrópodos) Facultad de Ciencias Universidad de Barcelona
Realizada la presentación y lectura de la Tesis en fecha 30 de junio de 1972, obtuvo la calificación de SOBRESALIENTE «cum laude>>.
SELLO DE LA FACULTAD Barcelona, 27 de Enero de 1973
El Decano,
FRANCISCO SALES VALLES
Vernet- Dep. LegaL 38.767 ·1973

INTRODUCCION
El tema de esta tesis doctoral, me fue sugerido por mi malogrado maestro el Dr. GARCIA DEL CID, por tratarse precisamente los Opiliones, de un grupo de Arácnidos que no habían tenido nunca investigadores en nuestro país.
Que esta úmple alusión a l Dr. GARCIA DEL CID, sirva de emocionado y respetuoso recuerdo a la memoria del que fue fundador y primer Director del Instituto de Biología Aplicada, siendo mi primer maestro y el que tan acertadamente me inició en los eswdios aracnológicos.
Al Dr. GADEA, actual Director de dicho Instituto y fiel continuador de la labor iniciada por m i antecesor maestro, debo mi más profundo reconocimiento, no sólo por haber asumido la responsabilidad de dirigir esta tesis, sino además, por haber puesto a mi alcance todos los medios necesarios para llevarla a feliz término.
Las actividades aracnológicas en nuestra Patria, resultan muy desoladoras, si comparamos el estado actual de estos estudios en España, con el de los demás países, ya que ú ltimamente los estudios aracnológicos han sufrido un impulso extraordinario hasta el punto, de que la Aracnología se ha independizado de la Entomología y ha adquirido vida propia.
Si tenemos en cuenta además, que por su situación y distinta configuración geográfica, España ofrece una gran riqueza en diversidad de biotopos y una abundante fauna autóctona, no es difíci l imaginar, que cualquier labor aracnológica emprendida podría ser muy fecunda, y esta rama de la Zoología, saldría del letargo que la caracteriza.
Siendo este nuestro deseo, ofrecemos en estas líneas los aspectos más destacados que encierra nuestro trabaj o, con la esperanza de estimular el interés hacia los estudios aracnológicos, a la vez que contribuir con esta pequeña aportación al conocimiento de los Opiliones de la fauna ibérica
Procedencia del material estudiado y técnicas
La rea li zación de este trabajo ha supuesto una larga labor de recolección y estudio de más de 1.300 ejemplares procedentes de 52 localidades distintas. Las regiones más exploradas han sido, el litoral cantábrico, la región catalana y la meseta central.
Siempre que nos ha sido posible hemos procurado realizar prospecciones escalonadas en e l uanscurso del aúo en la misma localidad, al objeto de obtener ejemplares en diferentes fases de desarrollo. Cuando ello no era factible, hemos completado en el Taboratorio, el ciclo «i n vitrO>> de especies recolectadas jóvenes, al objeto de obtener el estado adulto de las mismas.
En cuant0 a las técnicas empleadas para la recolección, traslado de ejemplares vivos, al imentación de larvas, técnicas de disección, etc., debemos señalar que han sido muy diversas, ya que salvo el medio acuático ,estos seres pueblan los biotopos más dispares, y es obvio que e] estudio de especies cavernícolas, frondícolas , humícolas, etc., se traduzca En la utilización de instrumentos y técnicas también diferentes .
Para el transporte de especies nivícolas y de alta montaña en donde las temperaturas son , s!empre muy inferiores a las del laboratorio, hemos usado un tipo de nevera portatll, de los muchos modelos que se encuentran en los comercios.
Para extraer los ejemplares de muestras de tierra o de detritus se han usado los agaratos de BERLÉSE y el método de NüRDMAN. '
En el estudio de los ciclos de desarrollo, el mayor obstáculo fue la formación de colonias de hongos, especialmente en la cría de aquellas especies, que requerían
-3-

un grado de humedad próximo a la saturación, problema que fue resuelto por un lado, con la limpieza y desinfección de los recipientes a base de fungicidas, tales como nipagina, y por otro lado, estableciendo una corriente de aire y perforando las tapas de los recipientes para su ventilación.
Para aquellas especies cuyo ciclo de desarrollo en la naturaleza se realiza en condiciones climatológicas iguales o muy semejantes a las del jardín de nuestra Universidad, se instalaron en el mencionado jardín, unas jaulas acompañadas de un termómetro de máxima y mínima y de un higrómetro; para poder registrar las variaciones ele temperatura y humedad.
Para las especies procedentes de las zonas de alta monlai'ia, o de las regiones áridas del sur de la península, nos hemos valido de un tipo de estufa llamada «Refritherm» de temperatura regulable y provista de un ventilador que produce una corriente de aire muy útil para E-vitar la formación de colonias de hongos.
Las larvas fueron alimentadas con drosofilas procedentes del Departamento de Genética y con fragmentos de un isopodo (Porcelio scaper) muy abundante en el jardín ele la Universidad Estas dos fuentes de material de alimentación nos ofrecen, además de la ventaja de estar ubicadas en la misma Universidad, la de asegurarnos una continuidad en su obtención.
Es preciso, no obstante, hacer constar aquí, que el objeto perseguido no es un estudio ele los factores ecológicos que condicionan la biología de estas especies, sino la obtención de datos biológicos de enfoque sistemático, tales como los cambios morfológicos en las fases postembrionarias y la historia estacional de huevos, ninfas y adultos, para poderlos utilizar en el estudio sistemático de estas formas.
CONSIDERACIONES GENERALES
El Orden Opiliones es uno de los II con representantes vivos de la cbse Arácnidos, la cual con los dos pequeños grupos de artrópodos marinos Merostomas y Pantópodos, forman el subphylum Quehcerados, que con el opuesto Mandibulados, forman el phylum Artrópodos.
Los Opiliones ocupan el tercer puesto según la importancia numérica de especies. Los Araneidos son los más numerosos seguidos de los Acaros .Ambos dominan a los nueve órdenes restantes, no sólo en número de especies ,sino en la abundancia de las mismas. El resto son grupos relativamente pequei'ios, entre los cuales los Opiliones destacan en relación a los demás.
En la línea de evaluación filogenética, los Opiliones quedan emplazados en una posición intermedia, más evolucionados que los Escorpiones y menos que los Araneidos, aunque la misma diversidad de formas dentro del orden, plantea difíciles problemas, que son interpretados de modo contradictorio y que hacen imposible establecer un orden cronológico aceptable.
Esta diversidad de formas y de organización se traduce asimismo, por modos de vida muy diversos, y todo ello viene expresado sistemáticamente por una división del Orden en tres subórdenes: Cifoftalmos, Laniatores y Palpatores.
La proporción de especies en cada uno de estos tres subórdenes, viene representada en la figura 3. (1)
Mundialmente las especies conocidas del suborden Laniatores, doblan casi el número de las de los Palpatores. Por el contrario, si comparamos los dos gráficos de la figura 3, vemos que los Palpatores ocupan casi la totalidad de nuestra fauna de Opiliones.
(1) La numeración de las figuras, corresponde a la del original de esta tesis.
-4-

LANIATORES
a b Figura 3. Proporción de especies en los tres subórdenes:
a) tauna mundial; b) fauna de !a Península Ibérica.
Bosquejo histórico y estado actual del íema
En el siglo XVIII los precusores de lo que más tarde sería la clase Arácnidos describieron las primeras especies, dentro del heterogéneo y primitivo grupo de LINNEO llamado «Insecta», que era !Tlás o menos lo que luego fue el Phylum Artrópodos, siendo emplazadas junto con Pignogónidos, Acaros, Insectos , Mirápodos y hasta Crustáceos.
El primero que empezó a vislumbrar lo que llegaría a ser el Orden, fue HERBST (1799), en su «Natursystems der ungeflügelten», pues reúne en dos fascículos todas las especies separándolas claramente del resto, y LATRETLLE (1798) nos ofrece la primera obra destinada a los Opiliones, titulada «Memoire pour servir de suite a l'Histoire des Insectes connus sous le mon de Faucheurs».
En la etapa final del siglo XIX, es cuando empiezan a sentarse las bases de la taxonomía de los Opiliones como Orden, y su división en subórdenes y familias.
Este primer período se caracteriza por una taxonomía tipológica clásica sin más preocupaciones que la de dar a conocer especies. Posteriormente cuando el concepto tipológico de especie va siendo paulatinamente abandonado y substituido por un concepto más dinámico, se inicia el período del estudio de poblaciones. Pero el grupo aracnológico, no está todavía influido por las nuevas ideas. más bien diríamos que ha sido en esta época, cuando ha proliferado más la descripción de especies y géneros monotípicos, sin tener en cuenta para nada las nuevas orientaciones _
Donde ello se hace más patente es en los estudios del gran aracnólogo alemán RüEWER, el cual desde 1910 nos va ofreciendo una serie de trabajos que culminan en la gran monografía mundial de 1923 titulada «Die Weberknechte der Erde>> con sus sucesivos suplementos hasta 1957. Estos trabajos constituyen la obra de recopilación sistemática más valiosa aparecida hasta nuestros días, y a pesar de que no le negamos todo su valor, por otro lado, es un ejemplo patente del caos producido por la excesiva e injustificada proliferación de géneros y especies.
En 1923 . cataloga 1.700 especies en 500 géneros, de los cuales 300 constan sólo de 1 ó 2 especies. En los suplementos que van apareciendo hasta 1957, describe 62 géneros monotípicos más, y otros 45 que constan sólo de 2 ó 3 especies .
ROEWER, clasificó material de todo el mundo, dejando ,a su fallecimiento en 1963, una colección valiosísima que se halla depositada actualmente en el Museo de Senckenberg, y que constituye el material de base imprescindible para todo trabajo de revisión taxonómica, el cual es a todas luces cada vez más apremiante para poder definir las especies a la luz de Jos actuales conceptos dinámicos de especiación.
-5-

Dificultades en la aplicación del concepto de especie biológica
La aplicación en la sistemática de los conceptos evolutivos, fertilizados sobre una base de interación de la herencia y la selección natural, ha supuesto la mtraducción de un nuevo co:1cepto de especie que se ha convenido en llamar <<especie biológica», en el sentido de que ella constituye un sistema separado de los demás, morfológica, genética, etológica y ecológicamente, por una discontinuidad biológica.
No obstante, el especialista se encuentra a veces con grandes dificultades para la aplicación de este concepto, dificultades que, aunque afectan por igual a diversos grupos, son especialmente pertinentes en los Opiliones, sobre todo en los Laniatores de nuestra fauna, ya que muchas de sus especies son fósiles vivientes, considerados como reliquias, que forman poblaciones reducidas muy difíciles de localizar y recolectar, lo que se traduce en una penuria de material, que imposibilita la aplicación ele métodos de trabajo, requeridos por el actual concepto dinámico ele especie.
Por otro lado, los Opiliones suelen tener un ciclo vital anual, y en algunas especies más amplio todavía, lo que dificulta asimismo el estudio genético de poblaciones, como se viene realizando con otras especies de desarrollo más breve.
Pero aun en el supuesto de que pudieran salvarse estas dificultades , hemos podido percatarnos, que los Opiliones llevan un retraso ele información con relación a otros grupos mejor estudiados, y ello hace que algunas etapas de su conocimiento estén todavía por realizar, cuando las mismas han sido ya superadas en otros gru · pos. Así, la labor primitiva de distinguir y diagnosticar morfológicamente las especies, no ha sido resuelta todavía debido a una incompleta valoración de datos tan elementales como variaciones morfológicas por diferencia de edad, dimorfismo sexual, etc., por lo que, la sistemática de algunas familias aparece poco estable y se pone de manifiesto la necesidad de un trabajo de revisión, que será necesario empezar por la base, y que es lo que abordamos, en parte, en estas líneas .
CONSIDERACIONES TAXONOMICAS
Un capítulo entero está dedicado en el original de esta tesis, a ofrecer los conocimientos básicos de morfología externa de los Opiliones, describiendo un tipo morfológico primero y <>numerando después las variaciones de este tipo, con el análisis de todos los caracteres morfológicos de valor taxonómico. Apoyados por esta base morfológica, presentamos diversas observaciones sobre la taxonomía de los Opiliones, valorando los caracteres morfológicos mencionados y dando cuenta de los errores cometidos en los estudios realizados hasta la fecha.
De todos los errores observados, expondremos aquí los que aparecen con mayor frecuencia y que afectan a ;:nayor número de especies de nuestra fauna:
Lo En muchas ocasiones, se ha relacionado un carácter taxonómico con una categoría taxonómica dada, y en la . práctica esta correlación no se da siempre. Por ejemplo. el número ele artejos de los tarsos, no tiene valor taxonómico en la mayoría de Palpatores, en cambio en los Laniatores sí Aquí ha sido usado para discriminar subfamilias, géneros y especies, pero su uso no siempre ha sido acertado Jo que ha dado origen a una serie de géneros monotípicos, que deberán ser refun: didos al comprobar que, en este caso, este carácter tiene sólo valor específico.
2.0 Se han descrito como especies diferentes el 6 y la 'i' de una misma especie, por no valorar el diformismo sexual que en muchos casos es extraordinario. Estos caracteres sexuales secundarios no aparecen en los 6, hasta después de la última muda, por lo que las fases subadultas de algunas especies, no han sido debidamente interpretadas.
3.0 Se han descrito como especies diferentes los estados ninfales de otras especies, por el desconocimiento de los cambios morfológicos experimentados en el desarrollo postembrionariq. La progresiva variación individual, a través de una serie de estados juveniles o larvarios hasta llegar al estado adulto ha sido desestimada en la taxonomía de los Opiliones y constituye la causa más fr'ecuente de los errores cometidos en la sistemática del grupo. En la literatura abundan ejemplos de es-
-6-

pecies que han stdo descntas con varios nombres, los cuales no son ~ás q~e las formas juveniles de la mtsma especte. Estos errores .han SidO subsanaaos solo ~n parte por Jos Cl utores a ntenores, p ues siguen aceptandose como buenas especies y aun géneros, muchas formas q ue no lo son, y que en e l. c~:~rso de nuestros estuchas hemos podido demostrar que se t rata stmplemente de dtstmtas fases del desarrollo de una misma especie.
4.0 No se han valorado las modificaciones que Jos relieves quitino!>OS experimentan durante este desarrollo, siendo la causa de que en muchas ocasiones, se haya confundido la variabilidad individual con la de crecimiento.
5.0 Hasta fechas relativamente reciemes se ha venido descuidando el estudio de la morfología genita l en casi toda la sistemática de los Opiliones, la cual se ve tristemente afectada por ello. Su estudio reviste un doble interés, pues aparte de su valor en el diagnóstico, nos proporciona el conocimiento de si el ejemplar que estamos estu~iiando es o no adulto, [actor que ha sido muy descuidado por la mayoría de autores.
6.0 Por último, algunos datos biométricos usados para discriminar especies, han sido tomados de partes del cuerpo afectadas de gran plasticidad, por lo que el margen de error es considerable, y e l dato no tiene ningún valor
RELACIONES FAUNISTICAS
La fauna opi lionológica :1ctual de la Península Ibérica, consta cle 138 especies en las que están representados los tres subórdenes. Se conocen cuatro especies del suborden Cifoftalmos, ocho del suborden Laniatores (una de ellas se describe aquí), y el resto 125, pertenecen al suborden Palpatores.
No obstante, y a pesar de que son posibles nuevas aportaciones, probablemente el número total de especies se verá mermado en lugar de incrementarse, pues los estudios del ciclo vital de algunas de ellas nos obligan a fusionarlas, e incluso a poner en sinonimia algunos géneros.
De estas 138 especies, 10 de ellas han sido descritas por nosotros en publicaciones anteriores, siendo todas autóctonas de nuestra fauna. Estas 10 especies son las siguientes: Nemastoma dipentalum, N. ibericus, N. cristinae, Dentizacheus zuluetai, D. minar, D. ibericus, Lacinus carpeta11us, L. magnus, Dicranopalpus pullchellus y D. dispar.
A estos 10 endemismos descritos por nosotros, hay que añadir otros 51 ya conocidos, que hacen un total de 61 especies, lo que representa más del 45 % de la fauna total. Estas cifras creemos serán objeto de variación. tendiendo a un aumento de endemismos, ya que la revisión de las especies del Museo de Zoología de Barcelona y del Museo de Ciencias Naturales de Madrid, y las del Museo de París y Frankfurt, clasificadas por SIMON, RoEwER y KRAUS, nos demuestra que muchas especies han sido identificadas como elementos pirenaicos y centroeuorpeos ya conocidos, cuando en realidad se trata de una fauna autóctona, que será descrita oportunamente. La situación y distinta configuración geológica y geográfica que ofrece nuestra península, se traduce en este número tan elevado de formas endémicas.
De las 72 especies restantes, casi la mitad, que equivale a un 30 %, son pirenaicas y gran número de tóllas habitan tanto en la vertiente francesa como en la española, por Jo que debemos aceptar que, para estas especies, la función de los Pirineos como barrera faunística, es de escasa importancia.
Quedan 37 especies, de las cuales 26 son típicamente mediterráneas, representando un 16% del total, y de las cuales más de la mitad, son comunes a nuestra península y al continente vecino A pesar de ser bien manifiesta la conexión de nuestra fauna con el Norte de Africa, el número de especies comunes, probablemente, se verá disminuido, ya que al revisarlas, hemos visto que, en algunos casos, se trata de especies diferentes.
Por último quedan II especies de difusión holoarctica, en el Oeste y Centro de
-7-

Europa, o sea aproximadamente un 8 % del total, y una de ellas Phalangium opilio es cosmopolita.
Ante la imposibilidad de pres~ntar el estudio de toda la fauna ibérica dentro de esta tesis, a causa de su enorme extensión y la dilación en el tiempo que ello supondría, hemos elegido dos grupos de características totalmente opuestas, con la intención de demostrar que el uso de los diversos métodos de trabajo de que dispone actualmente el sistemático, está supeditado a la clase de material con el que trabaja.
Así, las especies ibéricas del primer grupo, Jos Laniatores, son todas ellas consideradas como reliquias de una fauna tropical muy abundante en el terciario, pero hoy prácticamente extinguida, formando reducidas poblaciones muy difíciles de prospectar y recolectar, y ello nos obliga a estudiarlas aplicando los criterios morfológicos tradicionales, ante la imposibilidad de trabajar c~m otras fuentes de información.
Las del segundo grupo, los Palpatores, son más evolucionados y la. mayoría forman poblaciones de gran número de individuos, de las que hemos obtenido muestras abundantes, que nos han permitido la elaboración de datos experimentales.
ESTUDIO OPILIONOLOGICO
SUBORDEN LANIATORES
Este suborden comprende en Ia actualidad alrededor de 2.500 especies de distribución casi exclusivamente tropical, por lo que la labor más importante descansa sobre autores americanos, siendo de destacar MEI.LO-LEITAO, SOAREZ, RINGUELET, GoooNIGHT, BRIGGS, etc. De todas las familias aceptadas, solamente tres ( Paralolidae, Phalangodidae y Travuniidae), poseen representantes europeos y de estas tres, sólo las dos últimas (Phalangodidae y Travuniidae), están representadas en la Península Ibéri~a. La última es casi exclusiv<>.mente europea, por el contrario Phalangodidae, tiene una gran representación de fauna tropical.
Fam. PHALANGODIDAE. Tarsos del 3.0 y 4.0 par de patas con dos uñas sencillas y separadas o simplemente fus ionadas en la base, pero siempre sin un largo pretarso común.
Fam. TRAVUNIIDAE Tarsos del 3." y 4.0 par de patas con una uña compleja, multirramosa, y siempre con un pretarso común, corto o largo.
PHALANG ODIDAE { TRICOMMATINAE (l." Distitarso de 2 artejos) p HALANGODIDAE ( >) >> 3 )
TRAvu : ;s:~.\c . { PELTO NYCH INAE (l."' Distitarso de 2 artejos) TRAVUNLI1\AE ( » » 3 )
De todos los caracteres usados en la discriminación de las subfamilías y géneros eurooeos, el que ba causado mayor impacto en la sistemática, es el número de artejos de los tarsos y distitarsos en los cuatro pares de patas, número que viene expresado por la llamada fórmula tarsal.
Ejemplo: 5(3):7(4):6:6. Los números entre paréntesis indican los segmentos del 1.0 y 2." distitarso y los sin paréntesis, el número total de artejos de cada tarso.
Esta fórmula, a pesar de su utilidad en sistemática, no se ha usado adecuadamente, valorándose por encima del nivel que le corresponde.
ROEWER (1935), insiste en considerar como un criterio decisivo ¡:;ara la separación de subfamilias, el número de artejos del primer distitarso, por presentar el pnmer par de patas una mayor constancia tn el número de artejos y las variaciones no ser tan acusadas como en los tres pares de patas restantes.
-8-

Si bien esto es cierto, esta constancia ha podido comprobarse que no es tan firme como suponía ROEWER, y este carácter, más variable de lo que se suponía en un principio, está siendo actua lmente objeto de discusión, AvRAM (1968).
Ciñéndonos exclusivamente a las especies europeas, vemos que la separación de las cuatr::J subfamilias existentes no puede sostenerse, pues en algunos ejemplares el número de artejos de los distitarsos es asimétrico y hemos hallado también en la misma especie individuos con 2 y otros con 3 artejos indistintamente, por otra parte, el resto de caracteres estudiados tampoco permite la separación.
Separación de géneros: postura disgregadora de Roewer
Los caracteres que se han usado hasta ahora para discriminarlos son los siguientes: 1.0
, número de artejos de los tarsos; 2.", presencia o ausencia de la prominencia ocular; 3.0 , posición y relieves de la prominencia ocular; 4.", áreas del escudo bien marcados o apenas reconocibles; S_", con o sin surcos transversos en las áreas del escudo; 6.", con o sin dientes o espinas en e l escudo y terguitos libres, y 7.", espiráculos claramente visibles o no.
Hasta hace relativamente pocos años, toda posible variación de uno de estos caracteres, constituía un género potencial, de donde el número de géneros monotípicos en los Laniatores era desmesurado. Posteriormente la experiencia demostró la variabilidad de algunos de estos caracteres y la necesidad de usarlos en combinación, y si bien esto es útil para separar los géneros tropicales y éstos de los europeos, no Jo es para separar estos últimos entre sí, los cuales se reconocen únicamente por el número de artejos de los tarsos, ya que el único criteriu de RoHVER para discriminar estos géneros, fue la tan ponderada fórmula tarsal, llegando a sembrar el wborden de un número extraordinario de género monotípicos.
La lista de géneros europeos que nos da ROEWER es la siguiente:
PHALA NGO UJDAE
1." Scotolemon 2." Ptycosoma 3." Querilhacia 4.0 Scotolemops S." Scotolemoniscus 6." Brigestus 7.0 Lucassa 8." Metascotolemon 9." Bucovina
10." Holoscotolemon
3 (2) : S (3) : 4: 5 3 (2) : 5 (3) : 5 : 5 4 (2) : 6 (3) : 4 : 5 4 (2) : 6 ( 4) : 4 : 4 5 (2) : 6 (3) : 4: 5 5 (3) : 6 ( 4) : 4 : 4 5 (3): 6 (5) : 5: 6 6 (¿) : 6 ( ?) : S : S 6(3) : 6(4): 4 : 4 7 (3): 6 (6): 4: 6
THAVUNIIDAE
11." Peltonychia 12.° Kratochviliola 13.0 Hadziana 14.0 Arbasus 15." Abasola 16.0 Dinaria 17." Travunia
3 (2) : 5 (3) : 4 : 4 3 (2): 6 (3): 4: 4 3 (2) : 6 (3) : 4 : 4 3 (2) : 6 ( 4) : 4 : 4 4 (2) : 6 (3) : 4 : 4 5 (2) : 6 (3) : 4 : 4 6(3):6(4):4:4
Hasta tal punto valora RoEWER esta fórmula tarsal que crea varios géneros nuevos únicamente por variaciones de esta fórmula y n~ existe ningún diagnóstico que nos dé los caracteres oponentes entre ellos.
Tenien~o en cuenta ad_emás, que de estos 17 géneros, II son monotípicos, y 3 co:1stan so~o dt; d_os EspeCies cada uno, no debe sorprendernos que al ser revisados, caigan en smommia muchos de ellos.
De los 10 géneros europeos de la subfamilia Phalangodinae sólo dos, Scotolemon y Ptycosoma, están representados en nuestra fauna . '
Subfam. Phalangodinae: géneros Scotolemon y Ptycosoma
El género Scotolemon fue establecido por LUCAS (1860), para unos ejemplares r~colectados por M. LESPE~ en va~ias _cue~as de l'Ariege, Francia: dedicando la espeCie a su recolector. El mismo ano aescnbe una segunda especie procedente de- la cueva Buco del Orso en Lombardía, y que dedica a M. LEPRIEUR.
-9-

A estas dos especies, leprieuri y lespesi, sigue una tercera descrita por SlMO N (1872) , con el nombre de terrícola, procedente de Córcega. La cuarta especie, doriae, fue descrita por PAVESI (1878), con unos ejemplares del Monte Cavo en los Apeninos. Nuevas recolecciones en el departamento de l'Ariege, conducen a la descripción de una quinta especie, lucasi, por SIMON (1879), con unos ejemplares de la cueva de Rieuforcoud. La sexta aportación corresponde a ROEWER (193S) con la especie reclinatus, procedente de Algeciras. Transcurre un cuarto de siglo sin nuevas aportaciones, hasta que KRAUS (1961), nos da a conocer la séptima especie roeweri, también de Algeciras. La octava y última aportación, krausi, es nuestra. Procede de la isla de Ibiza y ha sido descrita en RAMBLA 1972.
El género Ptycosoma fue descrito por SoERENSEN (1873), para la especie vitellinum procedente de Argelia, y se mantuvo monotípico hasta que en 1961, KRAUS, describió la segunda especie, catalonicum, con unos ejemplares de la provincia de Barcelona.
La elección de t::aracteres para las descripcinoes originales de estos dos géneros, no fueron muy apropiadas, siendo además la de SOERENSEN (1873), para el género Ptyosoma, escueta en extremo. La descripción original de LUCAS (1860), para el género Scotolemon es más completa que la del género anterior y sin entrar en detalles indicaremos solamente que a pesar de su mayor extensión no da ningún carácter que le oponga a Ptycosom:.~. .
Poco provecho puede sacarse de estas descripciones que descuidan lo más im· portante, que es la coordinación de caracteres entre los dos diagnósticos. Además, RoEWER (193S), al describir los dos géneros, comete algunos errores, los cuales se van repitiendo en publicaciones posteriores del mismo autor y de otros, llegando a un confusionismo que es preciso esclarecer.
Si analizamos primeramente los siete caracteres que se han usado para di scriminar los géneros de la subfamilia Phalangodinae, vemos que sólo el 3.0 nos sirve como carácter diferencial para separar Ptycosoma de Scotolemon. Habiéndose demostrado el escaso valor de los géneros creados atendiendo a un sólo carácter, nos vemos obligados a indagar si existen otros, cuya combinación pueda justificar el mantener estos dos géneros separados. ·
Estos caracteres existen y se manifiestan claramente en la especie tipo de cada uno de estos dos géneros: Scotolemon l espesi y Ptycosoma vitellinwn_ Además de la fórmula tarsal y de la posición y forma de la prominencia ocular, hemos hallado otros cinco caracteres que oponen claramente una especie a ]a otra, y son los siguientes:
Caracteres
Fórmula tarsal . . Prominencia ocular . Longitud de las patas Forma del cuerpo .
long. l." fémur I = ----
long. cuerpo Organo copulador . . Posición geográfica .
Scotolemon lespesi
3(2) : S(3) : 4 : S . Alejada del borde Largas . Piriforme
r = 112 .
Sin bolsa dorsal Pirenaica
Ptycosoma vitellinum
3 (2): S (3): S: 5 En el borde frontal Cortas Oval
I = 1/ 3
ron bolsa dorsal Mediterránea
Nadie que compare estos elatos podrá poner en duda la validez de estos dos géneros para las dos especies tipo, pero un detenido examen del resto de especies descritas y de los nuevos elementos que aportamos en este trabaio, nos demuestra claramente que estos caracteres genéricos se entrecruzan de tal manera, que la separación de todas las especies conocidas, en los dos géneros mencionados, difícilmente puede sostenerse.
Además hemos comprobado que la fórmula tarso.l utilizada para separarlos no es tan constante como se creyó en un principio, pues entre los 47 ejemplares de Ptycosoma catalonicum examinados, han sido hallados dos d con la fórmula 3(2):S(3):4:5 considerada cúmo propia del género Scotolemon, y otros u-es, un e:$ y dos <¡>. , con un número asimétrice> de artejos en el tercer par de patas.
10-

El numero de representantes europeos de este género es reducido, pues se conocen solamente nueve especies, de las cuales siete torman parte de la ta una ibéricobatear. Ue estas s1e te especies dos sol! nuevas, una de ellas la hemos dado a conocer en una nota sobre Opll!Ones de las baleares, RAMBLA 1972, y la otra viene descrita en el presente estudio.
Familia fravuniidae: estudio de los géneros Peltonychia y Kratochviliola
La familia Travuniidae fue dividida en dos subfamilias Peltonychinae (distarsos de L y j anejos) y J. ravununae (a1starsos de 3 y 4 artejOS). Estas dos subfamilias a eberan ser anuladas por no presen tar el número de arteJOS de los distitarsos, la cuns,ancw que se preteduiO en u n pnnC!piO.
La familia cons ta ele siete géneros, y aparte del número ele artejos de los tarsos, no existe ningún otro carácter para diferenciarlos. Pero estas variaciones de la tórmula tarsal 11ar. resultado ser t«.n inehcaces como en los Phalangodidae, por lo que es de suponer que muchos de estos géneros caerán asimismo en sinonimia, al aplicar las m ismas consideraciones que para la familia anterior. De ellos, sólo dos, están represe ntados en nues tra península.
Estos dos géneros, Peltonychia y Kratochviliola, fueron establecidos por RoEWER (1935 ), por su distinta fórmula tarsai 3(2) : + de 5(3): 4:4 en el primero, y 3(2): + de b( 3) :4:4 en el segundo . Observando estas dos fórmulas vemos que el único número sujeto a variación, es el que se refiere a los artejos del segundo tarso, número que puede ser distinto o interferir, como de hecho así ocurre, por lo que, de no existir otras diferencias, es te criterio no sirve para separar los dos géneros. La búsqueda de otras diferencias genéricas que pudiesen justificar el mantener estos dos géneros separados, ha sido infructuosa, ya que las relativas a la longitud ele la u ña final del tarso, las espinas del palpo y el distinto grado de pigmentación de los ojos, deben valorarse como específicas por ser expresión de diferentes niveles de adaptación a la vida cavernícola, que no alcanzan a modificar los caracteres genéricos. También deben valorarse a nivel específico, las diferencias ele las gnatocoxas y esternón. Todo ello nos demuestra que estos dos géneros deben fusionarse, para lo cual debemos tener en cuenta , que si bien fueron descritos por RO EWER al mismo tiempo, Peltonychia aparece primero con el n o 1 y Kratochviliola con e l 2.0
, por lo que este último debe caer en sinonimia del primero.
De Peltonychia se conocen seis especies y ele Kratochviliola tres. De estas nueve especies han sido halladas en la Penínsu la Ibérica tres, clavigera y piochardi del primer género y navarica del segundo. La especie navarica, no permite una separación genérica y las tres especies ibéricas, deben situarse en el mismo género Peltonychia.
En el original de esta tesis, figuran las llaves dicotómicas que hemos confeccionado para la identificación de las especies de los géneros Scotolemon y Peltonychia, además se describen no só lo los nuevos elementos hallados, sino que se actualiza también el diagnós tico ele los ya conocidos, dado que la identificación de estas especies resu lta harto difícil muchas veces, a causa de los errores cometidos en las descripciones anteriores.(
De la especie tipo de cada género. damos una descripción completa, de las demás exponemos únicamente los caracteres especí ficos diferenciales , ilustrando gráfica mente es tas diferenci as. Cada descripción va acompañada de sinonimias, variabilidad, numeración ele los tipos y paratipos revisados, lista de localidades y relación ele registro ele ejemplares.
Conclusiones sobre la sistemática de los Laniatores Ibéricos
Los r esultados obtenidos de l estudio sistemá tico de los Laniatores de nuestra fa una, nos permite formular las siguientes conclusiones:
J.o El estudio ele los •.:aracteres taxonómicos utilizados para discriminar subfamilias y géneros fueron valorados por encima del nivel que les correspondía, r esu ltando por ello, una profusión ele subfamilias y géneros, muchos de ellos mono-
11-

típicos, que han sido rebatidos en la presente rev1s10n. Así el género Ptycosoma debe pasar a ser sinónimo de Scotolemon, y Kratochviliola de Peltonychia, ya que los caracteres genéricos, sólo tienen tal valor en las dos especies tipo. En todas las demás, se mezclan de tal manera, que la separación de estas especies en dos géneros diferentes es a todas luces inadmisible.
e
d
Figura 54. - Scotolemon es panoli n. sp. ri
¡ 1 .¡ 1 !
a) cara lateral; b) cara dorsal; e) órgano copulador, lateral y dorsal; d) opérculo genital, de perfil y ventral.
-12-
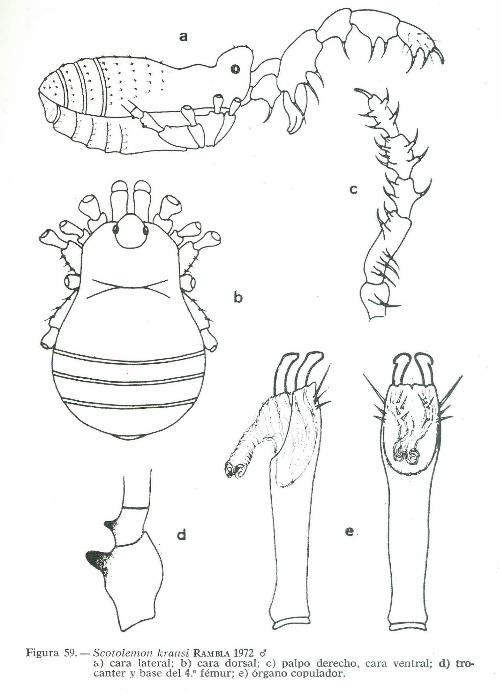
b
d e
Figura 59. - Scotolemon k rausi RAMBLA 1972 <J á) cara lateral; b) cara dorsal; e) palpo derecho , cara ventral; d) trocanter y base del 4.• fémur; e) órgano copulador.

2." Se ha demostrado que la especie Scotolemon lespesi, fue util izada como un cajón de sastre donde iban a parar todas la s espec1es de identificación dudosa, habiéndose identificado como /espesi muchos ejemplares que no Jo son . Así, KRAUS (1959, 1'!61 ), identificó como lespesi, m u c hos ejemplares de la especie cantábri<;a Peltonychia clavígera, procedente de la Corurí.a, Oviedo y Santander. Tamb1en identihcó como lespesi unos ejemplares de Algeciras que pertenecen· a la especie roetveri y otros ele Ibiza que han resultado ser una nueva especie, krausi R~A!BLA (1972). También nosotros cometimos el error de identificar como lespesi unos ejemplares de Portugal qu~ pertenecen a la especie roeweri. Por Jo tanto, ha quedado elemostrado que lespesi es excl usivamente pirenaica y que Jos demás ejemplares mencionados de Portugal, Cordirella Cantábnca, Andalucía e Ibiza, son en realidad otras especies diferentes.
3." La nueva especie espanoli constituye un aporte ele gran valor sistemático, como un elemento cavernícola muy pnmJttvo entre Jos Ylwlangoalaae, presentando caracteres que le sitúan por uu lado, muy próximo a lespesi entre los elementos pirenaicos, y por otro laao, cerca ele vzreLLinus, entre Jos elementos mediterráneos (figu ra 54) .
4." También se ha demostrado que los ejemplares de SntON identificados como lucasi, no son otra cosa que ejemplares ele lespesi, deshidratados y con el abdomen tan retraído que presentan un aspecto muy singular, siendo ello la causa de haberse descrito como especie distinta.
5." Del estudio de las tres especies ibéricas ele la fami lia Travuniidae, se elemuestra que navarica es un descendiente directo de clavigera, con la que está íntimamente emparentada, y ele la que se diferencia únicamente por un mayor grado de adaptación a la vida cavernícola, vié!íclose además, que es un ejemplo patente ele un proceso evolutivo en vigencia y no alcanzado plenamente, confirmado por la presencia actual de individuos en diferentes fases ele evolución. En efecto, dentro de una misma población aparecen en comunidad ejemplares con Jos ojos bien pigmentados y totalmente desprovistos de pigmento. Lo mismo podemos observar en cuanto a l grado de pigmentación ele Jos tegumentos y alargamiento de Jo~ apéndices, de io que se desprende que la cspeciación de navarica y su fragmentación ele ciavigera es un proceso vigente en la actualidad.
6." Por .lltimo, el estudio ele las formas fósiles, su parentesco con las actuales y la dispersión geográfica ele éstas, nos conduce a imaginar que Jos dos grupos de Lamatores 1baicos, [rawliidae y Phalangodidae, han debido llegar a nuestra península, por dos caminos distintos. Les Travuniidae a través de elementos europeos, que aesde ios Balcanes, Alpes y Pirineos, h<".n ::llcanzado la Cordillera Cantábrica. Las tres especies ibéricas se hallan confinadas en esta cordillera, llegando hasta el extremo occidental de los Pirineos, y aunque se las encuentra lo mismo en cuevas que fuera ele ellas, presentan modificaciones muy avanzadas de adaptación a Ja vida cavernícola.
Ei origen ele Jos Phala11godinae, creemos que debe buscarse en el desgajado macizo bético-rifeüo, existiendo un E'strecho parentesco entre las cuatro especies roeweri, recli11atus, krausi y catalonicum. Ellas bordean la costa sureste de nuestra península llegando hasta Ibiza. La especie krausi ele esta isla, parese ser la forma insular fraccionada de roeweri, su pariente más próximo, el cual se extiende por la región andaluza penetrando al Su:c· ele Portugal, siendo catalonicum, como su nombre indica, la forma propia ele la zona costera catalana Estas formas constituyen en la actualidad verdaderas ínsulas geográficas, las cuales ·mientras el aporte ele nuevos materiales no nos demuestre Jo contr;:rio, creemos más acertado consiclerarias como especies genuinas, pues el grado de continuidad genética que pudiese existir entre ellas es muy problemático, debido por un lacio a su aislamiento geográfico y por otro lado, a un probable aislamiento mecánico, debido a las diferencias anatómicas y de tamaño observadas en sus órganos copuladores.
SUBORDEN PALPATORES
En esta segunda parte, han sido estudiadas del suborden Palpatores, un grupo ele especies ele la subfamili::t 0/igolophinae, la cual consta actualmente ele siete géneros; ele los cuales sólo cuatro forman parte ele la fauna ibérica, ocupándonos en
-14-

estas líneas, exclusivamente de los dos géneros Odiellus y Lacinius, por recaer en ellos los errores que pretendemos corregir. De estos dos géneros se conocen en la actual idad un to tal ele 35 especies, 17 del primer género y 18 del segundo.
Un intensivo estudio de estas f01mas proseguido durante más de diez años, nos ha demostrado que los relieves quitinosos en los que se basa la taxonomía, pueden estar presentes o ausentes y variar de forma y tamaño, según la edad y el sexo del individuo, y ser modificados también por vanación geográfica dentro de la especie. Ninguno de estos factores ha sido tenido jamás en cuenta y el armazón dicotómico de estas especies, se ve tristemente afectado por ello. La falta de un justo criterio de valoración de los relieves quitinosos que adornan el cuerpo y los apéndices de estos Opiliones, ha desacreditado su valor, siendo por el contrario la realidad muy otra, ya que una interpretación correcta nos puede proporcionar buenas señales de identirí.caCión.
Al formularnos la hipótesis de si algunas de estas especies, podían no ser más que fases diferentes del desarrollo postembrionario de una misma especie, resulta evidente que el primer paso a segu1r en estos estudios, debería consistir en procurarnos intornnción, sobre dos puntos clave: la presencia de ninfas y adultos en el espacio y en el tiempo, y los cambios morfológicos experimentados durante el desarrollo postembrionario.
Estudio de muestras de la región catalana
Dentro de esta línea de trabajo, empezamos por los estudios experimentales, al objeto de obtener datos sobre el desarrollo estacional de estas especies.
Para ello se efectuaron recolecciones periódicas, en una zona acotada a tal efecto, obteniendo sucesivas muestras de una población, que llamamos piloto y que nos sirvió de referencia.
El estudio de este material nos demostró que estábamos en presencia de una especie con ciclo biológico anual, el cual resumimos como sigue: la eclosión se efectúa al empezar la primavera, las ninfas son abundantes hasta últimos de verano en que aparecen los adultos, la puesta se verifica en octubre y mueren a últimos de otoño. Los huevos pasan el invierno enterrados en el suelo, al empezar la primavera se verifica la eclosión y un nuevo ciclo recomienza. Durante cinco años consecutivos hemos observado este ciclo anual «in vivo», siempre con la misma regularidad y en el mismo biotopo elegido.
Posteriormente, al disponer en el laboratorio de los aparatos necesarios, para la cría de los Opiliones «in vitro», obtuvimos un ciclo de desarrollo completo, no sin antes haber sufrido algunos fracasos.
El conocimiento del desarrollo postembrionario ha puesto en evidencia que los últimos estados de la forma estudiada, coinciden exactamente con las descripciones de las tres especies siguientes: duriusculus, troguloides y galltcus.
1.0 duriusculus. Los ejemplares que se dejan identificar como duriusculus, se hallan presentes en la naturaleza desde primeros de julio a mediados de agosto Su tamaño es relativamente grande, la coloración es la del adulto y muchas de las ·formaciones tegumentarias están ya perfectamente desarrolladas, por lo que no es de extrañar que los autores que han descuidado el estudio de la genitalia, hayan podido tomar esta fase como adulta. No obstante, el opérculo genital está todavía cerrado, y los órganos copuladores se encuentran en un estado de desarrollo muy atrasado, por lo que el abdomen es más pequeño y redondeado y no tiene forma alargada como en las fases más avanzadas .
2.0 troguloides. Los ejemplares que se dejan identificar como troguloides, se hallan presentes en la naturaleza desde mediados de agosto a mediados de septiembre. Se trata de la fase subadulta de la especie, con todas las características propias del adulto, a excepción hecha de los caracteres sexuales secundarios. El opérculo genital empieza a abrirse sólo en parte y la genitalia se quitiniza v adquiere ya una longitud extraordinaria, por lo que el abdomen deja de ser redondeado y alcanza una
-- 15-

forma larga, es trecha y puntiaguda posteriormente, que ha sido descrita como característica de la especie.
3.0 gallicus. Por último, los ejemplares que han sufrido ya la última muda, o sea Jos adultos, se han identificado como gallicus y se hallan presentes desde mediados de septiembre a últimos de noviembre. Se trata de ejemplz,res qLJe han a dquir ido todas las características definitivas, con el opérculo genital tota lmente abierto, los órganos de cópula bien desarrollados y Jos cuatro fémures de las patas con hileras longitudinales de dientes tan to en el eS mo en la 2 . Los caracte res sexua les sesecundarios son bien manifiestos en el <i y aparecen juntos siempre después de la última muda.
Por Jo tanto, las t res especies citadas con más frec uencia de la región catalana por casi todos Jos autores que se han ocupado de estas formas, corresponden a tres tases del desarrollo postemb rionario de una misma especie. La forma ad ulta es gallicus, y las otras dos, duriusculus y troguloides, no son más que las ::íos últimas ninfas de gallicus. Habiendo sido troguloides (LUCAS 1874), la primera forma descrita, este nombre adquiere prior idad y durisculus (SIMÓN 1878) y gallicus (SIMÓX 'J879 ) de· berían ser sinónimos. La especie se denominaría ,pues, Odiellus troguloides (LuCAS 1847).
La revisión de los tipos, o en su defecto ejemplares de la serie típica y demás material de estas especies, nos ha demostrado la existencia de errores de diagnóstico en el material de museo, conduciéndonos a los siguientes resu ltados: las formas estudi ades corresponden a una sola especie con ciclo anual, que deberá ser llamada gallicus, ya que ninguno de los ejemplares europeos identi ficados como troguloides Jo son. La revisión de ejemplares de la serie típica, nos <.clara que lrogoluides, LLICAS 1847, del Norte de Africa, es una buena especie descrita con ejemplares adu ltos y di feren te de las especies europeas. El órgano copulado!' es distinto de Jos supuestos troguloides europeos, y los ejemplares de Europa clasificados como tales pertenecen a otra especie. Las diferencias en la genitalia masculina p ueden constituir un mecanismo de aislamiento eficaz en tre los ejemplares del norte de Africa y los del sur de Europa, a pesar de una semejanza morfológica externa, tan manifies ta, que ha sido la causa de errores de diagnóstico, ;:¡J descu id c: rse el estudio de Ja genitalia.
Consideraciones sobre los dos géneros Odiellus y Lacinius.
Estos dos géneros proceden por escisión del primitivo género Acant holophus, C. L. KocH (1871 ), y se separan exclusivamente por la ausencia en el primero y la presencia en el segundo, de dientes o espinas en los fémures, patelas y tibias de Jos cuatro pares de patas.
Al seguir el ciclo biológico de es1as formas en el laboratorio, hemos visto que todos Jos Odiellus adultos de nuestra fauna, presentan los fémures dentados y que son sólo las ninfas las que tieoen Jos fém ures s in dientes. Además, en aquellas especies de las que disponíamos de abundantes muestras, pudimos comprobar la existencia de una variación clinal para este carácter. Por otro lado, hemos fracasado en todos los intentos de hallar la existencia de otros caracteres que nos permitiesen mantener estas formas separadas en Jos dos géneros mencionados. Al intentar aplicar nuevos caracteres, no conseguíamos más que complicar la nomenclatura, con el trasiego de especies de uno a otro género, que es en definitiva lo que se ha venido haciendo hasta ahora.
Por lo tanto, deberíamos adoptar el nombre del primitivo género Acantholophus para estas especies, pero este nombre fue dado con anterioridad por MAC LeaY (1935) a un coleóptero , po;:- lo que resulta ser homónimo y deberá reservarse para el coleóptero previament~ descrito.
Pero aparte del nombre, la homogeneidad ele este grupo de es pecies es indiscutible, y las formas es tudiad as deben pasar a forma r parte del género Lacinius THORELL (1876), por la presencia en todas ell as de dientes o espinas en los artejos de las patas, pudiendo afirmar que e l género Odiellus, no existe en la Península Ibérica. Si ulteriores estudios nos demostrasen que es ta uniformidad se extiende
-16-

por igual a !as demás especies del género Odiellus, éste debería ser invalidado y caer en sinonimia de Lacinius.
Información sobre otras especies d.e la Península Ibérica
Al estudiar las demás especies de la Península Ibérica, hemos seguido los mismos pasos que en el estudio de la especie gallicus, obteniendo de todas ellas ejemplares adultos con la cría de las ninfas, comprobando que éstas no son diagnosticables. No es de extrañar, pues, que al haberse descuidado el estudio de la genitalia con la consiguiente confusión de ninfas y adultos, hayan sido descritas las primeras, como especies diferentes de estas últimas, llegando incluso a pensar, si no estaríamos en presencia de una sola y única especie, viéndonos obligados a sinonimizar todas las formas descritas de la península.
Pero una tal simplificación no correspondería a la realidad, ya que el estudio de los d adultos (algunos de ellos desconocidos hasta ahora, y obtenidos por pirmera vez en el laboratorio con la cría de las ninfas) nos demuestra que es tamos en presencia de varias especies , cuyos d poseen muy buenos caracteres para diferenciarlos siendo las <;l las que son mucho más difíciles de diagnosticar, hasta el punto de que algunas de ellas son prácticamente indistinguibles.
La revisión de los tipos, paratipos, y de todo el material recolectado de la Península, nos lleva a una integración del número de especies conocidas de nuestra fauna, de tal manera que las quince especies citadas, quedan reducidas a seis: spinosus, gallicus, echinatus, carpetanus, seoanei y carpentieri.
N ' DE ESPECIES CITADAS CORRECCIONES N• DE ESPECIES REALES
ODIELLUS GALLICUS . ... . .. VALIDA LACINIUS GALLICUS
ODIELLUS DURIUSCULUS .......... NINFA
ODIELLUS SIMPLICIPES .... .. . .. NINFA
ODIELLUS BREVISPINA . . .... . ..... NINFA
LACINIUS ECHINATUS .. . .. ... VALIDA LACINIUS ECHINATUS
LACINIUS RUENTALIS SINONIMIA
LACINIUS CARPENTIERI ......... VALIDA ROEWERITA CARPENTIER I
LACINIUS MAGNUS ........... .. SINONIMIA
LACINIUS CARPETANUS ....... . . . VALIDA .... .... LACINIUS CARPETANUS
ODIELLUS SPINOSUS .. .. ...... .. VALIDA ........ LACINIUS SPINOSUS
ODIELLUS SEOANEI ............. V AL IDA ...... .. LACINI US SEOANEI
ODIELLUS TROGULOIDES .• NORTE DE AFRICA
LACINIUS EPHIPPIATUS ...... CENTROEUROPA
L ACIN 1 US DENTIGER . ....... CENTRO EUROPA
LACINIUS ANGULIFER ..... NORTE DE · AFRICA

Los da tos obtenidos para llegar a este resultado, son en síntesis, Jos sigiuentes:
1.0 Las especies simplícipes y brevispina descritas por SIMÓN 1879, no son buenas especies, ya que la revisión de ejemplares de la serie típica, nos demuestra que se trata de ninfas hembras no diagnosticables.
2.0 La revisión de los tipos de echinatus (LUCAS 1847) y ruentalis KRAUS 1961, nos muestra que son la misma especie, por lo que ruentalis cae en sinonimia de echinatus por prioridad en la nomenclatura.
3." También son ~guales las especies carpentieri RoEWER 1953 y magnus RAMBLA 1960. En este caso magnus pasa a ser sinónima de carpentieri por la misma razón.
4.0 La especie seoanei SLV!ÓN 1879, de la cual se ha ex traviado el tipo, fue descrita con una <¡l joven, según nos dice el mismo autor, lo que nos hizo dudar en principio de la validez de la misma. La obtención de o y 'í' adultos de la localidad típica en septiembre de 1971, nos permite afirmar la validez de seoanei, cuyas características difieren de las demás especies conocidas.
5.0 Las citas para España de BACELAR 1925, K!<AUS 1961 y MARCELLlNO 1967, para las especies ephippiatus C. L. KocH 1835, dentiger C. L. KocH 1848 y angulifer SIMÓN 1878, son erróneas y se trata en realidad de otras especies. Las dos primeras habitan exclusivamente en la Europa Central y la tercera en el Norte de Afi·ica.
Valoración de otros caracteres como factores de aislamiento
De las seis especies ibéricas, spinosus, echinatus y carpentieri, se discriminan bien por el uso exclusivo de los caracteres morfológicos señalados. La discontinuidad morfológica entre ellas es de tal magnitud, que es obvio que nos hallamos en presencia de tres formas que han alcanzado un alto grado ele especificación, con el consiguiente aislamiento genético, ostensible no sólo en sus rasgos morfológicos sino ecológicos v biogeográficos, para que podamos considerarlas como especies genuinas. La últinn, a su vez, se separa marcadamente de las demás, hasta el extremo de constituir el género monotípico, Roeweritta SILHAVY 1965.
Por el contrario entre gallicus, carpetanus y seoanei , no existen diferencias tan acusadas, y cabe preguntarnos si estamos o no en presencia de verdaderas especies, que respondan al concepto actual de especie biológica.
El estudio de su biología nos ha proporcionado de>tos de indiscutible valor para contestar a esta pregunta. Por una parte, tieDen diferentes pautas en la épocR ele cría y no coinciden en su desarrollo estacional. Por otra parte, clivergen también ecológicamente y e tológicamente, actuando estas diferencias como factores de aislamiento , cuando co inciden en el espacio (simpatría ).
En este sentido, hemos estudiado muestras ele diferentes poblaciones de Cataluña y Me~::: : ::: Central.
1.° Cataluña . - En esta región existen dos especies simpátridas, gallicus y es7 77inosus. La primera forma pob laciones ele gran número ele individuos, mientras que en la segunda el número ele individuos es muy escaso, de tal manera que al estudiar las primeras muestras, las interpretamos en principio, como aberraciones de la pri~ mera especie, gallicus. Pero la obtención de mayor número de muestras en años sucesivos , no5 permitió estudiar su ciclo biológico, llegando a la conclusión de que se trata de dos especies diferentes que son simpátriclas.
En las zonas donde gallicus y spinosus se superponen, el desarrollo estacional ele esta última, está a lgo avanzado con relación a la primera. Los adultos ele spinosus aparecen ya, desde últimos ele julio a primeros ele agosto, y a mediados ele septiembre cuando los adultos de gallicus son más abundantes, los de spinosus han verificado ya la puesta y han completado el ciclo. ·
Etológicamente también se difere ncian, ya que la especie gallicus muestra tendencia al gt·egarismo y se la puede hallar form ando agrupaciones de gran cantidad
-18-

de individuos Por el contrario spinosus vive más en solitario, ya que de esta especie hemos hallado siempre individuos aislados.
En cuanto a la proporción numérica de las dos especies en la naturaleza, hay una diferencia muy considerable, ya que el número de individuos recolectados de spinosus, representa aproximadamente un 10% del de gallicus.
2." Meseta Central. - Las muestras obtenidas en la Meseta Central, proceden de las provinc ias de Guadalajara, Madr id y Burgos, siendo las más numerosas las de la Sierra de Guadarrama. En esta cordillera conviven también dos especies simpátridas. Una de ellas es spinosus igual que en Cataluña, pero que se hace más abundante, y la otra es carpetanus que aquí sustituye a gallicus.
Hace unos aííos, RAMBLA (1959), en un estudio titulado <<Üpiliones de la Sierra de Guadarrama», identificamos los ejemplares de spinosus, como Odiellus duriusculus. Al revisados hemos podido comprobar que estos ejemplares son ninfas de spinosus, que se dejan identificar como diuriusculos, y al desconocer en aquellas fechas el desarrollo postembrionario de spinosus, fueron mal identificados.
El desarrollo estacional de carpetanus y spinosus tampoco coinciden, siendo el de la primera especie posterior al de la segunda. Las numerosas capturas realizadas a lo largo de la Cordillera de Guadarrama, nos han permitido observar además otro.> fac tores de aislamiento.
Spinosus vive en la meseta hasta alturas no superiores a los 1.000 m., y su biotopo de elección es el bosque seco, donde e l Pinus silvestris va siendo substituido por Quercus toza, con abundante mantillo esclerófilo . Por el contrario, carpetanus aparece a partir de los 1.000 m., en el bosque de Pinus silvestris, en las zonas más húmedas del mismo, con subvuelo forestal invadido por Pteridium aquilinum. Esto no impide que ambas especies se encuentren y se superpongan en las zonas de contacto (alrededor de los 1.000 m.), que son las mesófilas, con subsuelo forestal de Cistus lauriforius (j aras) y Genista purgans. En estas zonas de contacto donde ambas especies se superponen, y se anula el aislamiento ecológico y geográfico (altura), el factor que aquí parece actuar como mecanismo de aislamiento, es su distinto desarrollo es tacional.
Conclusiones sobre la sistemática de las especies ibéricas de los géneros Odiellus y Lacinius
Los resu ltados obtenidos del estudio morfológico y biológico de estas especies nos permiten formular !as siguientes co;Jclusiones sistemáticas:
1.0 La separación de estas especies en los dos géneros Odiellus y Lacinius, no puede mantenerse, ya que no existe ningún carácter morfológico ni. biológico de validez taxonómica, que apoye esta separación. Por lo que todas las especies ibéricas pasan a formar parte del género Lacinius, por la presencia en todas ellas de dientes o espinas en los fémures de las patas.
2. 0 Los caracteres morfológicos utilizados para discriminar las especies, han demostrado ser válidos . pero fueron mal u t ilizados y entre las causan más frecuentes de error des tacan la de confundir la variabilidad ind ividual con la de crecimiento, y el desconocimiento del ciclo biológico, habiendo sido descritas como especies diferentes las ninfas de otras especies ya conocidas.
3.0 Todas las citas que existen de la región catalana son falsas. Así , duriusculus, seoanei, troguloides, brevispina y simplicines, no son más que diferentes estados ninfales de gallicus, especie que se cita de Cataluíía por primera vez.
4.0 El estudio morfológico y biológico de otras poblaciones del resto de la península, nos permite afirmar que ruentalis es sinónima de echinatus, y que magnus lo es de carpent ieri. Además, las especies troguloides y anqulifer son del Norte de Africa, y ephippiatus y dentiger de Cen troeuropa, y ninguna de ellas vive en nuestra fauna, con lo cual las 15 especies citadas de la península, quedan reducidas a seis: gallicus echinalus, carpentieri, carpetanus, spinosus y seoanei.
-19-

5." Hemos pod ido comprobar que cuando dos de estas especies son simpátridas, tienen diferentes pautas en la época de cría y no coinciden en su desarrollo estacional por otra parte también divergen etológica y ecológicamente, actuando estas diferencias como factores de aislam iento .
6.0 El estudio biogeográfico de estas seis especies a que queda reducido el género en nuestra fauna. nos muestra una gran proporción de endemismos. Un 50% son autóctonas: seoanei, carpenlieri y carpelanus. De las tres restantes, dos echinatus y gallicus son mediterráneos, y una, spinosus, es centroeuropea muy abundante en el centro de Europa y mucho menos frecuente en la Península .
7.". Por último, despojadas estas seis formas de todas las interpretaciones erróneas que han perturbado su sis temát ica, y al preguntarnos si cabe considerarlas como especies genuina~, creemos poder dar una respuesta afirmativa para echinatus, carpcntieri y spinosus. En cuanto a gallicus, seoanei y carpetanus, actualmente sólo pueden ser consideradas como tres buenas morfoespecies, por la discontinuidad morfológica entre los <}, pero aceptando la posibilidad de que el aporte de más material, colme estas discontinuidades y nos obligue a modificar nuestro criterio actual sobre su rango taxonómico.
- 20 -

BIBLIOGRAFIA
AVRAM S. - 1968. Laniatores cavernicoles de Roumania. Revision des genres Sco· tolerrwps Rwr. Brigestus Rwr. et Bucowina Rwr. (Opil. Phalangodidae). Ann. Spéléol., 23 (3): 641-665. Moulis
BACELAR, A. - 1928. Arácnidos Portugueses. III Catálogo sistemático, dos Arácnidos de Portugal citados por diversos autores (1931-1926). Bull. Soc. Portug. Sci. Nat. 10 (17): 169-203. Lisboa.
BERLAND, L. - 1949. Ordre des Opilions. En: Traité de Zoologie, Grassé, VI: 761-793. Masson. Paris.
DRESCO, E. - 1952 a Repartition de Querilhacia querilhaci (Lucas) (Opiliones Phalangodidae). Notes biospéo., 7: 81-82.
HANSEN, H. y SOERENSEN, W - 1904. On two orders of Arachnida XI. 178 p . Cambrigde Uni. Press.
JUBERTHIE, C. - 1957 a. Développemmt de deux Opilions Phalangiidae, Odiellus gallicus (E. Simón), et Homalenotus quadridentatus (Cuvier). C. R. Acad. Sci., 244; 2747-2750. Paris.
JUBERTHIE, C. - 1956 b. Notes sur le biotope et la répartition geographique de quelques Opilions franc;ais. Bull Soc Zool. Fr., (5-6): 331-336. París.
KOLOSVARY, G. - 1929. Die Weberknechte Ungarns. Studium Verlag, Budapest. KRATOCHVIL, J. - 1958 a. Die Hohlenweberknechte Bulgaries (Cyphophthalmi und
Laniatores). Prace, 30 (9): 371-396. Praga. KRAUS, A. - 1959 b. Weberknechte aus Spanien (Arachn., Opiliones). Mitt. Zool.
Mus. Berlín. 35 (2): 293-304. Berlín. KRAUS, A. - 1961. Die Weberknechte der Iberischen Halbinsel (Arach., Opiliones).
Senck. biol., 42 (4): 331-363. Frankfurt am Main. LUCAS, H. - 1860 a. Observations sur un genre nouveau d'Arachnide trachéenne
(Scotolemon lespesi) qui habite les grottes de l'Ariege. Ann. Soc. Ent. Fr., 8: 973-983. París
LUCAS, H. - 1860 b. Note sur le Scotolemon Leprieurii, nouvelle espece d'Arachnide trachéenne, qui habite la grotte de l'Ours (Buco del Orso) en Lombardie. Ann. Soc. Ent. Fr., 8: 984-988. París.
MELLO-LEITAO, C. De. - 1936. Les Opilions de Catalogne. Treballls Mus. Cii'x. Nat Barcelona. (sér. ent.), 11 (9): 1-18. Barcelona. ·
RAMBLA, M. - 1959. Contribución al estudio de los Opiliones de la fauna ibérica. Opiliones de la Sierra de Guadarrama. Pub. lnst. Biol. Aplicada, 29: 59-110. Bar· celona.
RAMBLA, M. - 1966. Contribución al estudio de los Opiliones de la Fauna Ibérica sobre el género Dentizacheus Rambla 1956. Descripción de Dentizacheus minor n. sp. Publi. Inst. Biol. Aplic., 40: 153-164. Barcelona.
RAMBLA, M. - 1967 c. Opiliones de Portugal. Revista de Biología, 6 (1-2): 1-34. Lisboa.
RAMBLA, M. - 1972. Opiliones (Arachnida) de las Baleares. Rapp. Comm. int. Mer. Medit., 21: 89-92.
ROEWER, C. F.- 1923. Die Weberknechte der Erde, Jena. ROEWER, C. F. - 1925. Opilioniden aus dem nordlichen und ostlichen Spanien,
gesammelt von Dr. F. HAAS, in den Jahren 1914-1919. Senckenbergiana, 7 (5): 177-179 Frankfurt am Main.
ROEWER, C. F. - 1949 a. Uber Phalangodiden I. Senckenbergiana, 30 (1-3): 11-61. Frankfurt am Main.
ROEWER, C. F.- 1953. Mediterrane Opiliones Palpatores. Abh. naturw. Ver. Bremen, 33 (2): 201-210. Bremen.
SILHAVY, V - 1965. Die Weberknechte der Eupnoi aus Bulgarien. Acta entomal. bohemosloc., 62: 369-406.
SIMON, E. - 1879. Les Arachnides de France. VII. París.