caiman crocodilus (cope, 1868) en la costa atlántica y ... · resumen caiman crocodilus es la...
TRANSCRIPT

Análisis filogenético de Caiman crocodilus (Cope, 1868) en la Costa Atlántica y Pacífica
Colombiana empleando marcadores de ADN mitocondrial.
Gonzalo Jiménez Alonso
Universidad Nacional de Colombia
Facultad de Ciencias, Area Curricular de Biología
Bogotá D.C., 2016

2
Análisis filogenético de Caiman crocodilus (Cope, 1868) en la Costa Atlántica y Pacífica
Colombiana empleando marcadores de ADN mitocondrial.
Gonzalo Jiménez Alonso
Tesis de Maestría en Biología
Línea Genética
Directora
Martha Lucia Calderón Espinosa
(Instituto de Ciencias Naturales)
Asesor
Paul Bloor
Universidad Nacional de Colombia
Facultad de Ciencias, Area Curricular de Biología
Bogotá D.C., 2016

3
Gracias por llevarme en tus alas y ayudarme a creer en mí…
En la Ceiba te encuentro…

4
A Sarita y Anita,
el motor mi vida.
A Nancy,
por su amor, paciencia y compañía incondicional en este camino, gracias por creer en mí.
A mis Padres,
por su apoyo y guía en el camino.

5
TEXTO DE AGRADECIMIENTOS
Todo mi agradecimiento a mis tutores, Paul y Martha por haberme dado la oportunidad de
seguir con el desarrollo este proyecto a pesar de los muchos tropiezos. Gracias por su co-
nocimiento, experiencia y apoyo.
A Manuel Hoyos, por ser un compañero incondicional y compartir desinteresadamente sus
conocimientos, tiempo, experiencia y apoyo en los momentos difíciles.
A Ursula aunque te fuiste pronto, siempre estabas ahí escuchando y dando un consejo, me
hiciste mucha falta.
A la Universidad Nacional de Colombia y el programa de posgrado por haber contribuido
enormemente a mi formación académica.
Al Ministerio de Ambiente y Desarrollo Sostenible, Medio Ambiente, por la financiación del
presente trabajo a través del CONVENIO 27 de 2009, SUSCRITO ENTRE EL MINISTERIO DE
AMBIENTE Y DESARROLLO SOSTENIBLE Y LA UNIVERSIDAD NACIONAL DE COLOMBIA - INS-
TITUTO DE GENÉTICA para “La Consolidación del Laboratorio de Criminalistica Ambiental”
Este trabajo se llevó a cabo bajo CONTRATO No. 48 DE 2012 ACCESO A RECURSOS GENÉTI-
COS PARA INVESTIGACIÓN CIENTÍFICA SIN INTERÉS COMERCIAL.
A Carolina, Sandra, Carlos, Tomas, Sergio, Sebastián, Lina, Angélica y todas las personas que
compartieron conmigo sus opiniones, pensamientos, conocimientos y consejos, en diferen-
tes momentos durante el trabajo de campo, laboratorio y en conversaciones casuales del
día a día, gracias por su apoyo.
A todos, muchas gracias.

6
RESUMEN
Caiman crocodilus es la especie con la distribución más amplia entre los Crocodylia neotro-
picales. Ocupa toda la franja intertropical desde México, a través de América Central, lle-
gando hasta América del Sur en el centro de Brasil. La sistemática del género Caiman, pre-
sente en Centro y Suramérica, es aún objeto de profunda controversia y confusión; las es-
pecies reconocidas por algunos autores no son aceptadas por otros como consecuencia de
las incongruencias resultantes de los análisis morfológicos y moleculares.
Existe además una gran discusión respecto a la existencia o no de las diferentes subespecies
de C. crocodilus. Los hallazgos provenientes de estudios recientes en los que se plantean
hipótesis de filogenia para la especie no incluyen datos de las subespecies presentes en
Colombia, por lo que se desconoce aún el parentesco e identidad taxonómica de las pobla-
ciones de C. crocodilus presentes en el país. Evaluar si las poblaciones de C. crocodilus dis-
tribuidas en el Caribe y Pacífico Colombiano representan uno o más linajes es de gran rele-
vancia no solo por la necesidad de dar una respuesta a la controversia taxonómica, sino por
la importancia que tiene la especie en el contexto del uso que se le da en el mercado de
pieles a nivel mundial.
En este trabajo, analizamos la variación molecular de Caiman crocodilus con el objetivo se
evaluar el parentesco filogenético de las poblaciones de Caiman crocodilus de la Costa Ca-
ribe y Pacífica Colombiana y su correspondencia con el estado taxonómico actual de C. c.
fuscus y C. c. chiapasius.
La variación molecular fue realizada con los genes Citocromo b (CytB) y Citocromo oxidasa
I (COI). La matriz se construyó con muestras de 78 individuos de 15 localidades distribuidas
en las regiones Pacífica y Caribe de Colombia, el Valle del Magdalena y Catatumbo. Se iden-
tificaron 10 sitios polimórficos en la secuencia del gen COI y 23 sitios polimórficos en el gen

7
CytB. Al analizar las secuencias en conjunto (con 33 sitios variables) se encontraron 19 ha-
plotipos distintos.
En el presente estudio, el grupo C. c. fuscus/chiapasius se compone de tres subclados que
corresponden a: (1) Caiman crocodilus fuscus / Caiman crocodilus chiapasius en México a
Honduras, (2) Caiman crocodilus fuscus de Nicaragua al Norte de Costa Rica y (3) Caiman
crocodilus fuscus del Sur de Costa Rica a Colombia (Costa Pacífica, Costa Atlántica, Valle del
Magdalena y Catatumbo; incluidos los paratipos de la Isla Gorgona.
Los haplotipos encontrados en el presente estudio se agruparon en dos haplogrupos en los
cuales para el primero se encuentran 11 haplotipos localizados en zona de la costa Atlántica,
Valle del Magdalena, Catatumbo y Urabá Chocoano, el segundo haplogrupo se encuentra
agrupandos 7 haplotipos los cuales se encuentran localizados en el Pacifico Colombiano y
norte del Chocó solapando con el primer haplogrupo en la zona del Urabá y en el magda-
lena medio.
Los linajes de Caiman crocodilus mostraron un patrón filogeográfico concordante con atri-
butos fisiográficos presentes en el área de distribución. La fecha de divergencia obtenida
mostró que los eventos cladogénicos en Caiman crocodilus coinciden con el periodo má-
ximo de orogénesis de las cadenas montañosas en Mesoamérica y en el noroccidente de
América del sur durante el Plio-Pleistoceno.
Palabras clave: ADN Mitocondrial, Babilla, Caiman crocodilus, Citocromo B, Citocromo Oxi-
dasa I, filogenética.

8
INTRODUCCIÓN
En el género Caiman (Spix 1825), Medem (1981, 1983) reconoció tres especies: Caiman
sclerops, C. yacare y C. latirrostris. Años más tarde, Groombridge (1987) dividió a Caiman
en dos especies: C. crocodilus (sinónimo de C. sclerops) y C. latirrostris desconociendo la
propuesta de Medem. No obstante, King y Burke (1989) así como Busack y Pandya (2001)
reevaluaron esta propuesta dividiendo nuevamente el género en tres especies: C. crocodi-
lus, C. yacare y C. latirrostris. En la actualidad, ésta es la taxonomía aceptada para el género,
el cual se encuentra únicamente en el Neotrópico, donde se distribuyen hasta una altura
máxima de 500 metros sobre el nivel del mar (msnm). Las últimas dos especies son endé-
micas de América del sur, mientras que C. crocodilus se extiende desde México meridional
hasta el Norte de Suramérica, incluyendo las zonas de desborde del Río Amazonas. De
acuerdo con evidencia geográfica, filogenética y fósil, se ha planteado que C. crocodilus
puede tener su origen en América del sur (Vanzolini y Heyer, 1985; Brochu, 2000, 2004;
Aguilera et al., 2006; Martin, 2007).
Caiman crocodilus es la especie con la distribución más amplia entre los Crocodylia neotro-
picales. Ocupa toda la franja intertropical desde el noroccidente de Chiapas, México, hasta
la vertiente Pacífica de Honduras, ambas costas de Nicaragua, Costa Rica y Panamá, el litoral
Pacífico de Colombia y el suroccidente del Ecuador, la región del Caribe, y al oriente de los
Andes en toda la cuenca de la Orinoquia y Amazonia hasta el noroccidente de Bolivia. Ha
sido introducido intencionalmente en la Isla de Pinos, Cuba, Puerto Rico, las Antillas Meno-
res, Florida (E.U.A) y en la Isla de San Andrés (Colombia). Se encuentra desde el nivel del
mar hasta los 500 m de altitud (Rueda-Almonacid et al., 2007).
El nombre más antiguo para la especie es Lacerta crocodilus (Linnaeus, 1758). En 1801, Sch-
neider propuso el nombre Crocodylus sclerops, pero más adelante, Anderson (1900) demos-
tró que el ejemplar tipo no correspondía al cocodrilo africano, por lo que retomó el nombre

9
inicial. Schmidt (1928) prefirió adoptar la combinación Caiman sclerops, como el más apro-
piado para la especie; sin embargo, actualmente se recomienda la combinación nomencla-
tural Caiman crocodilus teniendo en cuenta el principio de prioridad (Chirivi, 1971; Rodrí-
guez, 2000). Como es de esperarse en una especie tan ampliamente distribuida, existe una
apreciable variedad intraespecifica, es así como dentro de Caiman crocodilus (Linnaeus,
1758) se reconocen cuatro subespecies: C. c. fuscus, C. c. crocodilus, C. c. chiapasius y C. c.
apaporiensis (Medem, 1981, 1983).
C. c. fuscus (Cope, 1868: Perosuchus fuscus), descrito del río Magdalena, Colombia;
fue reconocido como especie distinta por varios autores, entre ellos Schimidt (1928)
y Dunn (1945), pero se consideró más adelante como subespecie de Caiman croco-
dilus (Medem, 1958).
C. c. chiapasius (Bocourt, 1876: Alligator chiapasius), descrito en el Valle de Tonalá,
Chiapas, México; es considerada por algunos autores sinónimo de C. c. fuscus, pero
como subespecie diferente según Medem (1962; 1983).
C. c. apaporiensis (Medem, 1955) desde el río Apaporis superior, Colombia.
C. c. crocodilus (Linnaeus, 1758: Lacerta crocodilus) descrito a partir de ejemplares
colectados en Guyana (Smith y Smith, 1977; Busack y Pandya, 2001) (Escobero-Gal-
van et al., 2011).
Caiman crocodilus crocodilus, comprende las poblaciones localizadas al oriente y sur de la
cordillera de los Andes y por tanto, incluye las poblaciones de la Amazonia (excepto las del
alto río Apaporis en Colombia), Orinoquia, Guayanas y las Islas de Trinidad y Tobago; el lí-
mite suroriental se localiza en el río Parnaíba en la Costa Atlántica del Brasil (Medem, 1981;
Medem, 1983; Rueda-Almonacid et al., 2007; Venegas-Anaya et al., 2008; Velasco y Ayar-
zagüena, 2010). Linnaeus (1758) observó las siguientes características: color del dorso verde
oliva con numerosas manchas y punteaduras café oscuras y pecas negras sobre la cabeza,
cuerpo y cola. Varias barras diagonales sepia o negras que descienden sobre el cuerpo y la
cola. Superficies ventrales amarillo crema uniforme. Sin grandes manchas negruzcas sobre

10
los lados de las mandíbulas, escamas grandes de los flancos agrupadas al menos en dos
hileras longitudinales superiores; escamas ventrales externas, de la mitad del cuerpo, me-
nos de dos veces el tamaño de las grandes escamas adyacentes de los flancos (Busack y
Pandya, 2001; Rueda-Almonacid et al., 2007).
Caiman crocodilus apaporiensis, restringida al alto río Apaporis, entre los raudales de Jiriji-
rimo y Puerto Yavilla, incluyendo los caños Tunía, Rerorú o Macayá y el río Ajaju; alto caño
Tacunema, en Colombia (Medem, 1981; Medem, 1983; Rueda-Almonacid et al., 2007; Ve-
lasco y Ayarzagüena, 2010). Medem (1955) lo diferencia de otras poblaciones de la especie
basado en su rostro muy alargado y comparativamente estrecho, lados de hocico paralelos
anteriormente, tabla craneal plana, fenestras supratemporales grandes, color del dorso
café amarillento brillante con manchas y vermiculaciones negras sobre la cabeza, 4 -6 barras
oscuras sobre el cuerpo y la cola, extremidades grises oscuras o negras y abdomen amarillo,
escamas grandes de los flancos sin osificaciones, pequeñas escamas granulares arregladas
regularmente en series longitudinales entre las grandes escamas laterales, cresta caudal
doble en los primeros 11-13 verticilos (Busack y Pandya, 2001; Rueda-Almonacid et al.,
2007).
Caiman crocodilus fuscus, se encuentra confinada a la hoya del río Magdalena (excepto el
alto Cauca), el Valle del río Sinú, el Río Ranchería y la planicie costera del Caribe en Colombia
(desde Turbo); en Venezuela habita la cuenca del Lago de Maracaibo y el litoral Caribe hasta
Yaracuy en la región de Chichiriviche (Medem, 1981; Medem, 1983; Rueda-Almonacid et
al., 2007; Velasco y Ayarzagüena, 2010). Algunos autores extienden su área de distribución
desde Nicaragua a los drenajes costeros del Atlántico de Colombia (Venegas-Anaya et al.,
2008; Escobero-Galvan et al., 2011). Cope (1868) lo describió basado en un único espéci-
men de esta manera: coloración del dorso de los adultos café claro, café oliva o amarillento,
sin flecos o manchas oscuras; extremidades café amarillento como el dorso, barras oscuras
tan solo visibles sobre la cola; cuerpo y miembros verde oliva, cabeza sin manchas negras

11
escamas grandes de los flancos débilmente careadas y osificadas; escamas pequeñas gra-
nulares forman series cortas e irregulares entre las grandes escamas laterales; cresta caudal
doble en los primeros 12-15 verticilos; hocico ancho (distancia entre el cuarto diente ma-
xilar y el extremo del rostro igual o menor a la anchura del hocico entre estos dos dientes),
palatinos expandidos lateralmente (Medem, 1981; Medem, 1983; Busack y Pandya, 2001;
Rueda-Almonacid et al., 2007).
Caiman crocodilus chiapasius, distribuido desde el sur de México, por la costa Pacífica de
Guatemala, El Salvador, y hacia el sur, en ambas costas de Honduras, Nicaragua, Costa Rica
y Panamá, hasta el litoral Pacífico de Colombia y Ecuador (río Esmeraldas y río Guayas),
incluyendo el valle del río Atrato, Golfo de Urabá y la Isla Gorgona (Medem, 1962; Medem,
1981; Medem, 1983; Rueda-Almonacid et al., 2007; Velasco y Ayarzagüena, 2010). Algunos
autores limitan su distribución a partir del Pacífico en Oaxaca y Chiapas, en la costa México
a El Salvador (Venegas-Anaya et al., 2008; Escobero-Galvan et al., 2011). Bocourt (1876) lo
describió con un hocico muy corto y liso; dorsalmente, escamado entero pardo oscuro, con
numerosas manchas y vermiculaciones negras. Lateralmente la mandíbula gris plomizo o
gris ratón en ambos sexos. La parte ventral es amarillo uniforme sin manchas negras, y
existe una sombra oscura en las tres primeras hileras de escamas. Numerosas escamas la-
terales del cuerpo y nuca anaranjadas (Medem, 1981; Medem, 1983; Busack y Pandya,
2001; Rueda-Almonacid et al., 2007).
Caiman crocodilus chiapasius fue descrito por Bocourt (1876) basándose en pocos ejempla-
res y haciendo comparaciones además con C. yacare, una especie diferente a C. crocodilus
pero que en esa época era considerada como una subespecie más de Caiman crocodilus
(Groombridge, 1987; King y Burke, 1989) y no tuvo en cuenta a C. c. fuscus en su análisis.
De otro lado, algunos autores (Boulenger, 1889; Schmidt, 1928) consideran a C. c. chiapa-
sius, como sinónimo de C. c. fuscus. No obstante la propuesta de Bocourt fue aceptada por
Medem (1962) como subespecie de Caiman crocodilus, dejando un manto de duda sobre la
validez de la subespecie C. c. chiapasius. La subespecie se distribuye a lo largo del pacifico

12
según Medem (1962), inclusive en las islas del pacifico de Colombia, sosteniendo que C. c.
chiapasius tenía centro de origen en México y que se había desplazado a través de América
Central, hacia la Vertiente del Pacífico y del Caribe de América Central, a las tierras bajas
del Pacífico de Colombia y Ecuador.
Venegas-Anaya et al. (2008) y Escobero-Galvan et al. (2011), son los autores que más cues-
tionan la distribución de C. c chiapasus propuesta por Medem, restringiéndola únicamente
a Mesoamérica, y se basan en las conclusiones encontradas de Busack y Pandya (2001),
quienes estadísticamente infieren que las diferencias en los caracteres morfológicos pro-
puestos para la asignación de las diferentes subespecies de C. crocodilus no son significati-
vas, planteando la necesidad de una revisión de la taxonomía de C. crocodilus. Por esta ra-
zón surge la idea de realizar otro tipo de análisis que clarifique la taxonomía de la especie
(Rueda-Almonacid et al., 2007; Venegas-Anaya et al., 2008; Velasco y Ayarzagüena, 2010;
Escobero-Galvan et al., 2011).
En el contexto de complejidad taxonómica en el que se encuentra el género Caiman, el uso
de herramientas moleculares constituye un complemento a los métodos clásicos de clasifi-
cación taxonómica mediante morfología. Puesto que el desarrollo de técnicas moleculares
ofrece metodologías certeras para la identificación de especies, poblaciones e individuos.
En la proposición de filogenias y de estudios intra-específicos, los caracteres moleculares
han sido paulatinamente preferidos por sobre otras fuentes de información (Wiens & Ree-
der, 1995; Sumida et al., 1998; Lanteri & Confalonieri, 2003). Lo anterior se debe, por una
parte, a la relativa facilidad con que se puede obtener gran cantidad de información, y por
otra, al desarrollo de métodos computacionales cada vez más confiables y objetivos al mo-
mento de inferir relaciones de parentesco (Huelsenbeck & Crandall, 1997; Avise, 2000; Nei
y Kumar, 2000; Felsenstein, 2004). La tipificación a partir de secuencias de ADN mitocon-
drial (mtADN) proporciona una huella genética de la especie y su origen geográfico (Palo-
mares et al., 2002). Las secuencias de mtADN surgieron como una opción popular dentro

13
de los marcadores moleculares, para la discriminación de especies y la identificación de es-
tructura filogeográfica, dado que los niveles de polimorfismo son capaces de proporcionar
la identificación de especies y relaciones entre distintas áreas geográficas (Eizirik et al.,
1998; Palomares et al., 2002).
Algunos autores como Vasconcelos et al. (2006) y Hrbek et al. (2008) realizaron estudios
filogenéticos con el apoyo de marcadores moleculares con la subespecie C. c. crocodilus, y
revelaron la existencia de linajes divergentes dentro de la subespecie. Adicionalmente,
Hrbek (2008) ha encontrado evidencia de hibridación (clados de mezcla) entre las especies
Caiman crocodilus y Caiman yacare.
Venegas-Anaya et al. (2008) realizaron un estudio filogenético de tres de las cuatro subes-
pecies de C. crocodilus, basado en marcadores mitocondriales (Citocromo b y Citocromo
Oxidasa I), en 45 individuos en diferentes zonas geográficas del área de distribución de la
especie. Los análisis hallaron dos clados al interior C. c. crocodilus y de C. c. fuscus/C. c.
chiapasius. Las relaciones entre C. c. fuscus y C. c. chiapasius no fueron claramente resuel-
tas, ya que se presenta una división de estos dos grupos en dos linajes diferentes, uno de
los cuales correspondió a C. c. fuscus y otro que corresponde a individuos de C. c. fuscus y
C. c. chiapasius (México). La presencia de haplotipos de C. c. chiapasius y C. c. fuscus en el
mismo clado se interpretó como producto de un evento de introgresión reciente. Estos re-
sultados confirmaron la divergencia entre los linajes Mesoamericano (C. c. chiapasius y C.
c. fuscus) y caimanes sudamericanos (C. c. crocodilus y C. yacare). El tiempo de divergencia
estimada entre C. c. chiapasius/C. c. fuscus de México y C. c. fuscus fue entre 2,5 y 2,9 mi-
llones de años, lo que corresponde con el cierre de la vía marítima del Pacífico-Caribe.
Desafortunadamente, dichos estudios genéticos han basado sus análisis en muestras que
no comprenden la distribución total de la especie ni representantes de todas las subespe-
cies reportadas, haciendo que las designaciones sub-específicas propuestas sean difíciles de
evaluar; en particular se plantean hipótesis de filogenia para la especie que no incluyen

14
datos de las subespecies presentes en Colombia, por lo que se desconoce aún el parentesco
e identidad taxonómica de las poblaciones de C. crocodilus presentes en el país.
Evaluar si las poblaciones de C. crocodilus distribuidas en el Caribe y Pacífico Colombiano
representan uno o más linajes es de gran relevancia no solo por la necesidad de dar una
respuesta a la controversia taxonómica, sino por la importancia que tiene la especie en el
contexto del uso que se le da en el mercado de pieles a nivel mundial, dado que Colombia
es el mayor exportador de pieles de crocodilianos en el mundo, con un volumen de cerca
de 600000 pieles en anuales (Larriera, 2004; Morales-Betancourt et al., 2013). El mercado
legal es posible sólo por medio de la cría en cautiverio y bajo la modalidad de “ciclo ce-
rrado”. Aun así, la Policía Nacional de Colombia, entre el año 2006 y el 2011, ha realizado
incautaciones de cerca de 18000 individuos de crocodílidos en Colombia provenientes del
tráfico de poblaciones silvestres (Morales-Betancourt y Lasso, 2013).
Dado que cualquier programa de manejo debe empezar con la identificación de las poten-
ciales unidades de conservación, es necesario contar con datos genéticos, como los que se
generarán de esta investigación, que brinden a las autoridades conocimiento más preciso
sobre áreas y/o poblaciones prioritarias de conservación. Esto con el fin de proporcionar
criterios filogenéticos y taxonómicos para plantear estrategias de control y vigilancia que
mitiguen el comercio ilegal de las poblaciones de C. crocodilus en el Caribe y Pacífico Co-
lombiano.
El objetivo de esta investigación es realizar un análisis filogenético de Caiman crocodilus en
la Costa Caribe y Pacífica de Colombia. Según lo reportado por varios autores (Medem,
1958, 1981; Chirivi, 1971; King y Burke, 1989; Rueda-Almonacid et al., 2007) en estas áreas
se encuentran dos de las cuatro subespecies reportadas en Colombia: C. c. fuscus en la costa
Caribe y C. c. chiapasius a lo largo de la costa Pacífica. Los resultados de esta estudio apor-
tarán criterios para definir las unidades de conservación de C. crocodilus en estas dos eco-
rregiones del país.

15
2. OBJETIVOS
2.1. Objetivo general
Evaluar al parentesco filogenético de las poblaciones de Caiman crocodilus de la Costa Ca-
ribe y Pacífica Colombiana y su correspondencia con el estado taxonómico actual de C. c.
fuscus y C. c. chiapasius.
2.1. Objetivos específicos
Determinar las relaciones de parentesco filogenético a partir de secuencias de ADN
mitocondrial de los genes citocromo b y citocromo oxidasa I de las poblaciones de
C. crocodilus en la Costa Caribe y Pacífica Colombiana.
Establecer la validez del estado taxonómico de las dos subespecies de C. crocodilus
reconocidas en estas regiones del país.

16
METODOLOGÍA
Muestreo
Se muestrearon 15 localidades representativas de las áreas de distribución propuestas en
este estudio, para las dos subespecies que se encuentran en la zona (Figura 1).
Figura 1. Localidades de muestreo de poblaciones de Caiman crocodilus. Los números y puntos negros indican
las localidades muestreadas, en verde el área de distribución de C.c. chiapasius y en naranja el área de distri-
bución de C. c. fuscus (Medem, 1962)
ID LOCALI-
DAD NOMBRE LOCALIDAD DEPARTAMENTO
NUMERO DE
INDIVIDUOS
1 Cerrejón La mina Guajira 6
2 Arjona Bolívar 5
3 Arboletes Antioquia 6
4 Puerto Berrio Antioquia 6
5 Ciénaga Costilla Cesar 1
6 Zulia- Tres Bocas Norte de Santander 6
7 Ciénaga de Marriaga Chocó 6
8 La Honda/ La Poza Chocó 6
9 Ciénaga La pedeguita /Rio
sucio Chocó 6
10 Ciénaga de bellavista /Bo-
jaya Chocó 6
11 Condoto/Jigualito Chocó 6
12 Laguna Barrigona Cundinamarca 6
13 Palermo Huila 6
14 Guapi Cauca 1
15 Roberto Payan Nariño 5

17
El trabajo de campo se llevó a cabo en los cuerpos de agua durante la noche; se realizó el
desplazamiento con bote y por medio de linternas se encontraron los especímenes, ubicán-
dolos por el color del reflejo de sus ojos (Naranja-Rojo), como lo describe Rueda-Almonacid
et al. (2007). La captura se realizó con la ayuda de pértigas y cuerdas (Figura 2).
Figura 2. A. Método de Captura de Individuos de Caiman crocodilus. B. Contención de Individuo de Caiman
crocodilus. C. Toma de muestra de escama caudal en Caiman crocodilus. D. Toma de muestra de sangre en
Caiman crocodilus. [Foto: Sandra Hernández]
De cada uno de los individuos se tomó muestra de sangre de la vena caudal con jeringa de
2ml y aguja N°20 y/o se cortó un fragmento de escama caudal (incluyendo músculo), con
un sacabocado. Las muestras se preservaron en tubos con etanol al 95% y se almacenaron
a -20° C hasta realizar la extracción de ADN.
A B
C D

18
Adicionalmente se tomaron muestras de cráneos de paratipos descritos por Medem (dos
de C. c. chiapasius (ICN-R 8688 y ICN-R 8709), una de C. c. apaporiensis (ICN-R 8763), que
reposan en el Instituto de Ciencias Naturales de la Universidad Nacional de Colombia.
Trabajo de laboratorio
Extracción y cuantificación de ADN
La totalidad de las muestras se procesaron en el Instituto de Genética de la Universidad
Nacional de Colombia. El ADN genómico total fue aislado de las muestras de tejido de Cai-
man crocodilus, mediante la digestión con Proteinasa K y su posterior extracción con el pro-
tocolo de sílice y tiocianato de guanidino, modificado de Rohland y Hofreiter (2007). Se
evaluó la calidad y cantidad del ADN aislado mediante geles de agarosa al 0,8% y
SYBR® Safe.
El método protocolizado fue la extracción de ADN basada en la adhesión al Oxido de Silicio
(SiO2 - mejor conocido como Sílice o Glass Milk) originalmente revisado por Rohland y Hof-
freiter (2007). Este método usa la adsorción selectiva de partículas de Sílice en suspensión
al ADN en presencia de sales caotrópicas (Vogelstein y Gillespie 1979; Boom et al., 1990;
Melzak et al., 1996; Rohland y Hofreiter, 2004; Parida et al., 2006; Rohland y Hofreiter,
2007). Aunque la solución cuente con la presencia de proteínas, carbohidratos, lípidos, RNA
y contaminantes que inclusive sean inhibidores de la reacción en cadena de la polimerasa
(PCR). Y es que la unión específica que se produce entre la Sílice y el ADN en medios ácidos,
asegura el aislamiento efectivo de los ácidos nucleicos, asegurando la remoción de los inhi-
bidores de PCR (Kemp et al. 2006; Hwan Young et al. 2010).

19
Tras el proceso de digestión, se adicionaron 800 mL de un buffer de unión a la Sílice que
contenía GuSCN (Tiocianto de Guanidina) [5 M], NaCl [25 mM] y Tris-HCl [50 mM]. La mez-
cla se agitó durante 5 minutos a 55°C (nunca se usó vortex) y se acidificó usando HCl [37%]
a un pH de 5. Luego, se adicionaron 10 µL de una suspensión de Sílice (Sigma-Aldrich, 0,5 a
10 micrones de tamaño de partícula) con un pH 3. La suspensión de Sílice (Rohland y
Hofreiter, 2007) debe prepararse previamente mediante decantación, la cual consiste en la
suspensión de 4,8 gramos de Sílice en un volumen de 40 mL de agua desionizada y estéril.
Tras una hora de decantación, 39 mL de la mezcla se transfieren a un nuevo tubo que se
deja decantar por cuatro horas adicionales. Finalmente 35 mL del sobrenadante se descan-
tan y se añade suficiente HCl [37%] para dejar la suspensión en un pH de 3%.
El procedimiento de extracción estandarizado para esta tesis fue el siguiente: Las muestras
frescas de tejido fueron almacenadas en tubos de colecta con etanol al 100% y refrigeradas
con una temperatura de -20°C. En el caso de las muestras de cráneos su tamaño permitió
separar los dos ramos de la mandíbula para poder obtener material del extremo anterior
interno y fueron pulverizadas siempre que fue posible utilizando un disruptor mecánico Tis-
sueLyser® QIAGEN. El polvo del cráneo fue almacenado en viales a -20° para evitar cual-
quier tipo de infestación por hongos o bacterias que puedieran actuar sobre ese material.
Al momento de realizar la extracción, un volumen cercano a los 3 mm3 de muestra fue de-
positado en un tubo de 1,5 mL. Posteriormente, las muestras fueron digeridas a 55°C du-
rante una noche usando 20 mL de Proteinasa K [2,0 mg/mL] en un buffer de lisis (el cual
contenía EDTA [0,5 M], Tris-HCl [10 mM], NaCl [100 mM], SDS Buffer [2%] y Tritón X-100
[0,5%]).
Tras adicionar la suspensión de Sílice a las muestras en reacción, estas se sometieron a una
agitación suave (por rotación) durante una hora a 55°C. Luego, las muestras fueron centri-
fugadas a máxima velocidad por dos minutos. El sobrenadante, fue removido y el pellet de
Sílice lavado dos veces por re-pipeteo usando un buffer de lavado (Etanol 50% v/v, NaCl

20
[125 mM], EDTA [1 mM], y Tris-HCl [10 mM]). El pellet de Sílice se secó finalmente en una
centrífuga de vacío a 55°C para luego ser re-diluido en 70 µL de una solución de pH 9 que
contienía Tris-HCl [10 mM] y EDTA [0.5 mM].
Amplificación, purificación y secuenciación
En el caso de tejido de escama y musculo, se amplificaron los genes completos mitocon-
driales citocromo b (CytB) y citocromo oxidasa I (COI), para la totalidad de las muestras
usando primers diseñados con base a datos de GenBank
(http://www.ncbi.nlm.nih.gov/genbank/) (ver Tabla 1).
Tabla 1. Primers mitocondriales utilizados
Las Reacciones en Cadena de la Polimerasa (PCR) tuvieron condiciones similares en todos
los casos. Se llevaron a cabo en un volumen final de 30 µL que contenían; 3X de Buffer de
PCR (Fermentas/Thermo), 2,4 mM de MgCl2, 0,6 µM de cada dNTP, 0,15 Unidades de
DreamTaq® DNA polymerase (Fermentas/Thermo) and 0,9 µM de cada primer.
Marcador Nombre Pri-
mer Secuencia (5'-3')
Tamaño fragmento (pares
de bases)
CytB Ccr_GluL1T GTCTGAAAAACCCCCGTTG 1236
Ccr_ThrH1T CCCGTCTCTGTCTTACAAGG
COI Ccr_SerH1T AGATGGTGGGGGTTCGATT 1558
Ccr_TyrL1dT AACACTCGGCCACTTTACC

21
Se realizó la amplificación de los marcadores CytB y COI en un Termociclador Mastercycler®
eppendorf, con el siguiente programa:
94°C …………………. 2 ̓
94°C …………………. 30 ̓ ̓
53°C …………………. 30 ̓ ̓
72°C …………………. 30 ̓ ̓
72°C …………………. 10 ̓
4°C …………………. Final ̓
Para las muestras de cráneo de museo, se amplificó el marcador mitocondrial citocromo b
(CytB). Para esto se diseñó una estrategia de amplificación del gen Citocromo b, usando un
set de 10 parejas primers que amplificaban regiones internas y flanqueantes del gen, par-
tiendo desde las últimas bases del complemento del gen ND6 hasta la región flanqueante
Treonina (THR) en la cadena pesada del genoma de la mitocondria.
Se buscó que las 10 parejas de primers amplificaran fragmentos cortos de alrededor de 200
pares de bases (pb) que pudieran solaparse en al menos 20 pb. Ello con el fin de “ensam-
blar” el gen Citocromo b completo más adelante. A estos amplicones que podían solaparse
se les denominó cóntigos (calco lingüístico del inglés: contig). Cada uno de los cóntigos fue
amplificado al menos 5 veces para cada uno de los individuos sometidos al análisis. Como
se explicará en la sección de ensamblaje de secuencias este procedimiento aseguró que no
se incorporaran errores en las secuencias definitivas debido a problemas como la desami-
nación (Briggs, 2007).
Se prepararon 10 soluciones “master mix” de 37µL que contenían; 1X de Buffer de PCR
(Fermentas/Thermo), 2 mM de MgCl2, 0,2 µM de cada dNTP, 0,4 Unidades de DreamTaq®
DNA polymerase (Fermentas/Thermo) and 0,5 µM de cada primer.
35 ciclos

22
Se realizó la amplificación de los marcadores CytB y COI en un Termociclador Mastercycler®
eppendorf, con el siguiente programa:
95°C …………………. 2 ̓
94°C …………………. 30 ̓ ̓
54°C …………………. 40 ̓ ̓
72°C …………………. 1̓ ̓
70°C …………………. 1 ̓
4°C …………………. Final
Los fragmentos amplificados fueron aislados mediante geles de agarosa al 1,2% y
SYBR® Safe. Se cortaron las bandas resultantes y se purificaron con el protocolo de purifi-
cación por precipitación con etanol y fueron resuspendidos en agua ultrapura. Posterior-
mente, se obtuvo secuencias con los mismos primers utilizados para la amplificación me-
diante el kit BigDye Terminator 3.1 y el secuenciador automático de análisis genético ABI
3500 (Servicio de Secuenciación y análisis Molecular – SSiGMol – Instituto de Genética, Uni-
versidad Nacional de Colombia).
Edición de secuencias y alineamiento
Los cóntigos de CytB obtenidos en el caso de cráneos de museo y que fueron amplificados
al menos cinco veces para corroborar los polimorfismos. Que posteriormente se ensambla-
ron y editaron usando los programas CodonCode Aligner ver. 4.2 (CodonCode-Corporation;
www.codoncode.com) y Geneious R9 ver. 9.0.5 (www.geneious.com). En ambos programas
se mantuvieron en default las condiciones de alineamiento de cóntigos.
40 ciclos

23
A las secuencias completas resultantes tanto de la amplificación de muestras de escama y
musculo y la de muestras de cráneos, se mejoró la intensidad de cada una de ellas por
medio del algoritmo PeakTrace Basecaller ® presentado en el sitio web nucleics.com (2015),
y se realizó el consenso de cada una de ellas mediante el uso del programa CodonCode
Aligner (CodonCode Aligner, Centerville, MA, USA). Los genes fueron alineados utilizando el
algoritmo CLUSTALW (Thompson et al., 1994) implementado en el programa BioEdit (Hall,
1999) y posteriormente traducidos a secuencias de aminoácidos utilizando MEGA 6 (Ta-
mura et al., 2013) para encontrar la presencia de codones de parada prematuros o pseudo-
genes. No se detectaron Indels en las secuencias de Citocromo b ni en las secuencias de
Citocromo oxidasa I. Las secuencias de los dos genes fueron unidas y analizadas en conjunto
para todos los análisis. Las secuencias fueron editadas en el editor de texto NOTE PAD++
(notepad-plus-plus.org, 2016) para construir matrices de secuencias donde se demarcaron
las posiciones ausentes con el símbolo “?”.
Curva de acumulación de haplotipos
Para estimar el número de haplotipos esperados a partir del muestreo, se construyó una
curva de acumulación de los haplotipos encontrados en cada localidad usando el programa
EstimateS 9 (Colwell et al., 2012), y aleatorizando la curva resultante con 999 iteraciones.
Tal curva muestra como el número de haplotipos se va acumulando en función al número
acumulado de muestras (Villarreal et al., 2004).
Análisis filogenético
Se realizaron dos análisis filogenéticos. El primer análisis evaluó la relación filogenética de
las especies de Aligatóridos americanos para establecer la monofilia del género Caiman y
su posición dentro de los Aligatóridos americanos. Para este análisis, se usó el gen CytB,

24
puesto que se podían encontrar secuencias homólogas a las encontradas en el presente
estudio en Genbank (http://www.ncbi.nlm.nih.gov/genbank/). La matriz analizada incluyó
71 secuencias con una longitud de 1236 pares de bases correspondientes al fragmento
completo de CytB. Diecinueve secuencias correspondieron a los haplotipos de la especie
Caiman crocodilus encontrados en este estudio, treinta y un secuencias correspondieron a
haplotipos reportados por Venegas et al. (2008) (Ocho haplotipos de C. c. crocodilus, 21 de
C. c. fuscus y dos haplotipos de C. c. chiapasius), tres secuencias obtenidas a partir de crá-
neos de paratipos descritos por Medem (dos de C. c. chiapasius (ICN-R 8688 y ICN-R 8709),
una de C. c. apaporiensis (ICN-R 8763), que reposan en el Instituto de Ciencias Naturales de
la Universidad Nacional de Colombia. Finalmente, se incluyeron las siguientes secuencias
obtenidas de Genbank: C. c. chiapasius (EU496845), C. c. crocodilus (EU496851, EU496852,
EU496857, EU496858, EU496860 y JF315306), Caiman yacare (JF315314), Caiman latirostris
(EU161673 y EU161674), Melanosuchus niger (JF315312 y JF315315), y las secuencias, co-
rrespondientes a las especies Alligator missisipiensis (EU496862), Paleosuchus palpebrosus
(KJ920382) y Paleosuchus trigonatus (EU496863), que actuaron como grupos externos en
los análisis filogenéticos.
En el segundo análisis se buscó dilucidar las relaciones filogenéticas al interior del género
Caiman, para esto se utilizaron dos marcadores moleculares, el fragmento completo de
CytB (1236pb) y una fracción de COI (658pb) puesto que solo se localizaron secuencias ho-
mologas de este tamaño para el mismo. Se incluyeron cincuenta y dos secuencias con una
longitud total de 1894 pares de bases, diecinueve secuencias correspondieron a los haplo-
tipos de la especie Caiman crocodilus encontrados en este estudio, treinta y un secuencias
correspondieron a haplotipos reportados por Venegas-Anaya et al., (2008) (8 haplotipos de
Caiman crocodilus crocodilus, 21 haplotipos de Caiman crocodilus fuscus y 2 haplotipos de
Caiman crocodilus chiapasius). Finalmente se incluyeron dos secuencias (GenBank
EU26061, EU496862, EU26062 y EU496863) correspondientes a las especies Alligator mis-
sisipiensis y Paleosuchus trigonatus, que actuaron como grupos externos en los análisis fi-
logenéticos.

25
En ambos análisis se realizó un esquema de partición discriminando los nucleótidos en cada
codón en primera, segunda y tercera posición. El mejor modelo de sustitución fue seleccio-
nado para cada partición usando el Criterio de Información Bayesiana (BIC), el Criterio de
Información de Akaike (AIC) y la Teoría de la Decisión (DT) implementados en el programa
JModeltest (Darriba et al., 2012).
La reconstrucción filogenética se llevó a cabo usando tres metodologías: (1) Máxima parsi-
monia - MP, (2) Máxima Verosimilitud - ML y finalmente (3) Inferencia Bayesiana - IB. MP
se condujo usando el programa NONA 2.0 (Goloboff, 1998) implementado en el programa
WinClada 10.0.8 (Nixon, 2002), usando el algoritmo de bisección y reconexión de árboles
(Tree Bisection-Reconection, TBR) + TBR. Se hicieron 1000 réplicas del análisis iniciando con
10 árboles cada análisis y manteniendo 100 árboles para el muestreo. Finalmente, se realizó
un consenso estricto de los árboles resultantes. Por su parte, para ML se utilizó el programa
GARLI v 2.0 (Adam et al., 2014). Se realizó una búsqueda estocástica para encontrar el mejor
árbol. El soporte de los nodos fue estimado mediante un bootstrap en GARLI usando 1000
réplicas. Finalmente, el análisis de IB se llevó a cabo utilizando el programa MrBayes 3.2
(Huelsenbeck y Ronquist, 2001), con 10.000.000 de generaciones y 4 cadenas ejecutándose
en paralelo, muestreando árboles cada 100 generaciones.
Se realizó una evaluación de la divergencia de las secuencias usando distancias genéticas
no corregidas (Distancias P) tanto para el gen CytB, como para los genes concatenados
(CytB/COI), por medio del programa MEGA versión. 6.06 (Tamura et al., 2013), utilizando
como método de estimación de la varianza Bootstrap con 500 iteraciones, incluyendo tran-
siciones y transversiones con deleciones parciales. El análisis de distancias puede conce-
birse como un soporte auxiliar al análisis de árboles los cuales fueron basados en el gen
CytB y en los genes concatenados CytB/COI (Neil y Kumar, 2000; Saldarriaga-Cordoba et al.,
2009). No es sorprendente que las distancias jueguen un papel importante en muchos as-
pectos de análisis de secuencias. En primer lugar, por definición, proporciona una medida

26
de la similitud entre las secuencias. En segundo lugar, si se asume un reloj molecular, la
distancia genética es linealmente proporcional al tiempo transcurrido. En tercer lugar, para
secuencias relacionadas por un árbol evolutivo, las longitudes de rama representan la dis-
tancia entre los nodos (secuencias) en el árbol. Por lo tanto, si la cantidad exacta de diver-
gencia de secuencia entre todos los pares de secuencias de un conjunto de n secuencias es
conocido, la distancia genética proporciona una base para inferir el árbol de la evolución
relativa de las secuencias (Strimmer y Haeseler, 2009).
Se realizó un análisis de redes de haplotipos usando los haplotipos encontrados en la matriz
de secuencias le los genes COI y CytB (en conjunto) del presente estudio y los encontrados
por Venegas-Anaya et al. (2008) para los mismos genes, con el fin de obtener una mayor
resolución de las relaciones intra-específicas (Mardulyn, 2012) y encontrar corresponden-
cias geográficas (Wang et al., 2008), que expliquen el origen de los haplotipos de C. croco-
dilus en Colombia. Este se realizó mediante la construcción de una red de haplotipos utili-
zando el programa TCS 1.21 (Clement et al., 2000). En el caso de asociación significativa
entre haplotipos y localidad geográfica, el análisis de clados agrupados es capaz de diferen-
ciar si esa asociación es resultante de eventos históricos, como fragmentación o expansión
poblacional, o de procesos recurrentes, como flujo génico. Para esto, los haplotipos fueron
agrupados en clados progresivamente inclusivos, diferenciándose por un único paso muta-
cional, de acuerdo con las reglas definidas por Templeton (2004; 2008) Templeton et al.
(1987) y Templeton & Sting (1993).
Estimación del tiempo de divergencia
Multiples puntos de calibración y la estimación de la distribución de cada punto ofrecen
más confianza al análisis de reloj molecular (Ho y Phillips, 2009). De acuerdo con la hipótesis
del reloj molecular, dos taxones que comparten un ancestro común hace t años deberían
haber acumulado más o menos el mismo número de sustituciones durante el tiempo t. En

27
la mayoría de casos, sin embargo, el antepasado es desconocido y no es posible probar
directamente la constancia de la tasa de evolución. El problema se puede resolver conside-
rando un outgrup, es decir, una o más especies lejanamente relacionadas (Lemey y Posada,
2009). Para aumentar los puntos de calibración este análisis utilizó 26 secuencias con 1140
pb, del gen mitocondrial CytB, de las diferentes especies que componen la familia Croco-
dylia, 24 obtenidas en GenBank (http://www.ncbi.nlm.nih.gov/genbank/), y 2 del presente
estudio (Tabla 2).
Las calibraciones de tiempo se aplicaron en cuatro nodos del árbol de especies (en millones
de años, Ma. 1 Unidad de Tiempo = 1 Ma) y la estimación de la distribución de cada punto
se basó en el trabajo de Oaks (2011). El primer punto de calibración fue el fósil, Allognat-
hosuchus mooki usando un Prior Gamma (64; 2) (Brochu, 2004), para calibrar el clado de la
Familia Alligatoridae en un mínimo 64 Ma (Oaks, 2011). Para calibrar el clado del género
Crocodylus, se utilizó al fósil Crocodylus palaeindicus, puesto que varios autores (Brochu et
al., 2000; Brochu et al., 2010; Oaks, 2011), lo sitúan en una posición basal entre los miem-
bros del género con 13 Ma, usando un Prior Exponential. Para calibrar la edad del grupo
Caimaninae se utilizó el fósil de Kuttanacaiman iquitosensis que data de cerca de 13 Ma
(Salas-Gismondi et al., 2015), se utilizó distribución lognormal. Y para apoyar los datos de
la calibración general del grupo de Aligatoridos se utilizó el tiempo de aparición del género
Paleosuchus en 9.5 Ma según los hallazgos de Oaks (2011), con una distribución Normal.
El tiempo de divergencia para este análisis se estimó usando aproximación de dateo mole-
cular Bayesiano (reloj relajado) implementado en el programa BEAST ver. 1.8.1 (Drum-
mond et al., 2012) utilizando de especiación en el árbol del tipo Birth_Death. La alineación
fue repartida en codones y el modelo de sustitución HKY con distribución gamma. Cada
análisis se ejecutó a partir de un árbol al azar, de 10000000 de generaciones asegurando
suficiente muestreo de parámetros. Los arboles fueron muestreados cada 100. Arboles
muestreados cada 1000 generaciones. El primer millón de árboles fueron eliminados verifi-
cándolo con el Software Tracer (Rambaut et al., 2014).

28
Tabla 2. Secuencias de CytB de Crocodilidos utilizados en análisis de tiempos de divergen-
cia.
Especie GenBank
Alligator sinensis JF315321.1 Alligator mississipiensis U496863_1 Paleosuchus palpebrosus KJ920382_1 Paleosuchus trigonatus EU496862_1 Melanosuchus niger JF315312_1 Caiman latirostris U161673_1 Caiman yacare JF315314_1 Caiman crocodilus crocodilus EU496860 Caiman crocodilus chiapasius EU496848 Caiman crocodilus crocodilus EU496855 Caiman crocodilus fuscus Cc 01 Crocodylus acutus JF315295.1 Crocodylus intermedius JF315297.1 Crocodylus johnsoni JF315277.1 Crocodylus mindorensis JF315252.1 Crocodylus moreletii JF315294.1 Crocodylus niloticus JF315273.1 Crocodylus novaeguineae JF315301.1 Crocodylus palustris FJ173276.1 Crocodylus porosus JF315290.1 Crocodylus rhombifer JF315262.1 Crocodylus siamensis JF315292.1 Gavialis. Gangeticus JF315302.1 Mecistops. Cataphractus JF315279.1 Osteolaenus tetraspis JF315288.1 Tomistoma schlegelii JF315305.1

29
RESULTADOS
Descripción de secuencias
Se obtuvo secuencias de los genes mitocondriales CytB (1236 pb) y COI (1558 pb), de un
total de 78 individuos de la especie Caiman crocodilus. Se identificaron 10 sitios polimórfi-
cos en la secuencia del gen COI y 23 sitios polimórficos en el gen CytB. Al analizar las se-
cuencias en conjunto (con 33 sitios variables) se encontraron 19 haplotipos distintos (Tabla
3). El haplotipo más frecuente para C. crocodilus es Cc 12 (n = 20), seguido del Cc 19 (n =
12). Los haplotipos Cc 04, Cc 05, Cc 06, Cc 07, Cc 09, Cc 13, Cc 16 y Cc 18 presentaron la
menor frecuencia, siendo solamente encontrados en un individuo (Figura 3).
7
4
6
1
1
1
1
5
1
3
4
20
1
2
2
1
5
1
12
Cc 01
Cc 02
Cc 03
Cc 04
Cc 05
Cc 06
Cc 07
Cc 08
Cc 09
Cc 10
Cc 11
Cc 12
Cc 13
Cc 14
Cc 15
Cc 16
Cc 17
Cc 18
Cc 19

30
Figura 3. Frecuencia (número de individuos) de los haplotipos de los 78 individuos de C. crocodilus encontra-
dos en diferentes zonas del país.
Tabla 3. Polimorfismo de las 33 posiciones variables que componen los diecinueve haploti-
pos mitocondriales de C. crocodilus, polimorfismos de COI y CytB y frecuencia por haplotipo.
Haplotipo Número de Posición Variable
n CytB COI
1 1 1 1 1 1 1 1 1 1 1 2 2 2 2 2 2 2 2 2 2 2 2
1 3 4 5 6 7 8 8 9 9 3 3 6 6 7 7 8 8 9 9 9 0 0 0 1 1 2 4 4 5 6 7 7
6 1 8 4 2 1 4 8 1 5 1 4 5 7 1 3 0 6 0 8 9 0 5 5 2 3 2 0 8 6 0 0 1
2 5 0 0 8 4 0 6 8 2 1 0 9 7 0 0 1 6 5 0 5 7 7 8 1 9 3 9 4 2 4 1 5
Cc 01 A C C T G C T T C C G T A C T T T T G G C G C T T T G C C C C A A 7
Cc 02 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . T . . 4
Cc 03 . . . . . . . . . . . . . . . C . . . . . . . . . . . . . . T . . 6
Cc 04 . . . C . . . . . . . . . . . C . . . . . . . . . . . . . . T . . 1
Cc 05 . . . . . . C . . . . . . . . . . . . . . . . . . C . . . . T . . 1
Cc 06 . . . . . . . . . . . . . . . . . . . . . . . . . . . . T . . . . 1
Cc 07 G . . . A . . C . . . . . . . . . . . . . . . . . . . . . . . . . 1
Cc 08 G . . . A . . C . . . . . . . . . . . . . . . . . . . T . . . . . 5
Cc 09 G . . . A . . C . . . . . . . . . . . . . . T . . . . T . . . . . 1
Cc 10 . . . . . A . . . . . . . T . . . . . . . . . . . . . T . . . . . 3
Cc 11 . . . . . A . . . . A . . T . . . . . . . A . . . . . T . . . . . 4
Cc 12 . . . . . . . . . . . . G . . . C . . A . . . . . . . . . T . . . 20
Cc 13 . . . . . . . . . . . . G . . . C . A A . . . . . . . . . T . . . 1
Cc 14 . T . . . . . . . . . . G . . . C . . A . . . . . . . . . T . . . 2
Cc 15 . . . . . . . . . . . . G . . . C . . A . . . . . . . . . T . G . 2
Cc 16 . . . . . . . . . . . . G . . . C . . A . . . . . . A . . T . . G 1
Cc 17 . . . . . . . . T T . . G . . . . . . A . . . . . . . . . . . . . 5
Cc 18 . . . . . . . . T T . A G . . . . . . A . . . . . . . . . . . . . 1
Cc 19 . . A . . . . . . T . . G . C . . C . A T . . C C . . . . . . . . 12
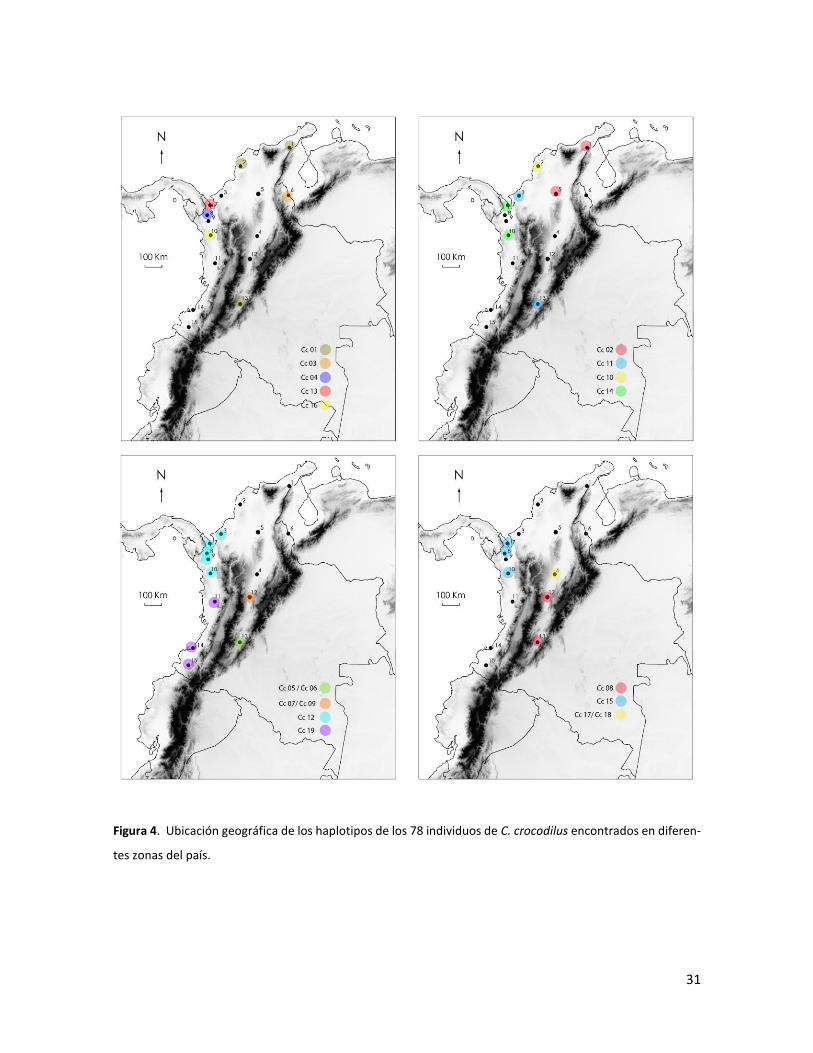
31
Figura 4. Ubicación geográfica de los haplotipos de los 78 individuos de C. crocodilus encontrados en diferen-
tes zonas del país.

32
Diversidad genética encontrada
De acuerdo con la diversidad genética encontrada (19 haplotipos utilizando dos marcadores
moleculares: COI y CytB) y con el ritmo de captura descrito en la curva de acumulación (con
999 iteraciones) de haplotipos de la Figura 5, se registra un valor cercano al 50% de la di-
versidad genética esperada para los marcadores COI y CytB para los sitios muestreados.
Esto contextualiza en el escenario regional los valores de la diversidad genética reportada
en este trabajo.
Figura 5. Curva de acumulación de haplotipos que demuestra el número de haplotipos encontrados en el
estudio (19) y el número de haplotipos que es posible encontrar en el área muestrada de acuerdo al ritmo de
captura (36 según la media de Michaelis Menten y 39 según la media del estimador Chao 2).
Relaciones filogenéticas inter e intraespecificas
De acuerdo con los criterios de selección de Inferencia Bayesiana y Criterio de información
de Akaike, implementados en el programa Jmodeltest (Darriba et al., 2012), los mejores
modelos de sustitución para cada una de las posiciones de los codones del gen CytB fueron
19
39,53
36,35
0
5
10
15
20
25
30
35
40
45
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
No
de
hap
loti
po
s
Oportunidades de Captura
Haplotipos
Chao 2 Media
Michalis Menten Media

33
el modelo General-Time-Reversible (GTR) +G para las posiciones uno y dos, y General-Time-
Reversible (GTR) +G +I para la posición codónica 3. En el caso del gen COI los mejores mo-
delos de sustitución fueron el modelo Hasegawa Kishino Yano (HKY) para las posiciones uno
y dos, y Hasegawa Kishino Yano (HKY) +G para la posición 3. Al momento de realizar los
análisis con una matriz que contenía los dos genes se mantuvieron los mismos modelos para
cada una de las particiones por posición de codón en los genes. De esta manera, el mismo
modelo de sustitución fue implementado en los análisis Bayesiano y de Máxima verosimili-
tud.
En el análisis de secuencias de CytB para conocer la relación de Caiman con otros grupos de
Aligatóridos, el árbol resultante del análisis bayesiano presentó la misma topología, que el
árbol de máxima verosimilitud. A su vez, estos árboles fueron similares (aunque no iguales)
a la reconstrucción de las relaciones evolutivas obtenida del análisis de parsimonia con con-
senso estricto, todos presentan valores de soporte altos en sus ramas para los tres análisis
(Figura 6).
En relación a los grupos externos, Alligator mississipiensis dista en un 21,3 a un 22,7 % de
los géneros Paleosuchus, Melanosuchus y Caiman; las dos especies del género Paleosuchus
(P. palpebrosus y P. trigonatus) presentan una divergencia del 17 al 18,6% con los otros
clados, y Melanosuchus niger con una divergencia del 11,3 al 12,6% con en genero Caiman.
El género Caiman se recupera como monofilético, con valores de soporte del 100%. Este
género está compuesto por siete clados que corresponden a Caiman latirostris, Caiman ya-
care, Caiman crocodilus crocodilus al oriente de Sudamérica, Caiman crocodilus crocodilus
occidente de Sudamérica, Caiman crocodilus chiapasius agrupado con Caiman crocodilus
fuscus (México hasta Honduras), Caiman crocodilus fuscus (Nicaragua al Norte de Costa
Rica) y finalmente Caiman crocodilus fuscus (Sur de Costa Rica hasta Colombia) incluyendo
los paratipos identificados como C. c. chiapasius por Medem (1962) (ICN8688 y ICN8709)
(Figuras 6, 7 y 8).

34
En el caso del clado que contiene los haplotipos de Caiman latirostris, éste se encuentra
separado de Caiman crocodilus/Caiman yacare, divergiendo en un 8,2 a 8,8%. En este úl-
timo se involucran los clados de C. c. fuscus/C. c. chiapasius, C. c. crocodilus/C. c. apaporien-
sis y C. c. crocodilus/ C. yacare, en los cuales internamente se puede observar que los ha-
plotipos de una misma región geográfica presentan distancias bajas entre ellos (0,0 – 0,5%)
(Figuras 6, 7 y 8).
Dentro del clado C. c. fuscus/C.c. chiapasius, se observan claramente dos grupos que pre-
sentan una divergencia del 1,8 al 2,1% entre sí, el primero incluye las secuencias de indivi-
duos localizados desde México hasta Honduras, cuatro de estas identificadas como C. c.
fuscus (Fuscus IV, Fuscus V, Fuscus VI (Venegas-Anaya et al., 2008) y EU496828 1), y tres
muestras correspondientes a individuos identificados como C. c. chiapasius (Chiapasius XX,
Chiapasius XXI, (Venegas-Anaya et al., 2008) y EU496845 1). En el segundo se encuentran
dos clados, uno con secuencias de individuos identificados como C. c. fuscus ubicadas desde
Nicaragua hasta el norte de Costa Rica (Fuscus I, Fuscus II, Fuscus III, Fuscus XV, Fuscus XVI
(Venegas-Anaya et al., 2008) y EU496839 1), y el otro agrupa secuencias de individuos loca-
lizadas desde el Sur de Costa Rica hasta Panamá (Fuscus VII, Fuscus VIII, Fuscus IX, Fuscus X,
Fuscus XVII, Fuscus XIX, Fuscus XXII, Fuscus XI, Fuscus XIII, Fuscus XII, Fuscus XIV, Fuscus
XVIII, Fuscus XXIII (Venegas-Anaya et al., 2008), EU496849 1), dos secuencias obtenidas de
los paratipos identificados como C. c. chiapasius en la Isla Gorgona, Colombia (Medem,
1962) y secuencias de individuos provenientes de Colombia y obtenidas en el presente es-
tudio (Cc 01 a Cc 19) (Figuras 6, 7 y 8).
Al comparar las divergencias entre los haplotipos de poblaciones de Caiman crocodilus de
la Costa Pacífica, Costa Caribe, Valles Interandinos y Catatumbo Colombiano, Panamá y Sur
occidente de Costa Rica con las poblaciones de Nicaragua y Nororiente de Costa Rica pode-
mos encontrar distancias del 1,1 al 1,5% (Figuras 6, 7 y 8).

35
Figura 6. Árbol de reconstrucción filogenética a partir de los métodos de Inferencia Bayesiana (IB), en el cual
se incluyen soportes de ramas indicados por los valores de probabilidad posterior/bootstrap con Máxima Ve-
rosimilitud (ML)/bootstrap con Parsimonia (MP), para Aligatóridos americanos, resultados provenientes del
gen completo del Citocromo B. Se muestran en las ramas soportes correspondientes a valores de la probabi-
lidad posterior y Bootstrap (IB/ML/MP).
Alligator mississipiensis
Paleosuchus trigonatus y Paleosuchus palpebrosus
Melanosuchus niger
Caiman latirostris
Caiman crocodilus chiapasius/Caiman crocodilus fuscus (Mé-
xico hasta Honduras)
Caiman crocodilus crocodilus
(Oriente de la Amazonia)
Caiman crocodilus crocodilus/Caiman crocodilus apaporiensis (Sur y Occidente de la Amazonia)
Genero Caiman
Caiman crocodilus fuscus (Sur de Costa Rica hasta Colombia)
Paratipos identificados como C. c. chiapasius (Medem, 1962)
(ICN8688 y ICN8709).
Caiman yacare
Caiman crocodilus fuscus (Nicaragua
al Norte de Costa Rica)
Ca
ima
n

36
0
50
100
150
200
250
300
00
,001
0,0
020
,003
0,0
040
,005
0,0
060
,007
0,0
080
,009
0,0
10
,011
0,0
120
,013
0,0
140
,015
0,0
160
,017
0,0
180
,019
0,0
20
,021
0,0
220
,023
0,0
240
,025
0,0
260
,027
0,0
280
,029
0,0
30
,031
0,0
320
,033
0,0
340
,035
0,0
360
,037
0,0
380
,039
0,0
40
,041
0,0
42
Nu
mer
o d
e co
mp
arac
ion
es En los clados C. c. crocodilus/ C. yacare vs. C. c. crocodilus/C. c. apaporiensis la divergencia
es del 3,8 al 4,2%. El clado C. c. crocodilus / C. yacare, involucra individuos identificados
como C. c. crocodilus ubicados en la parte central y oriental de la amazonia (Crocodilus
XXVIII, Crocodilus XXV, Crocodilus XXIV, Crocodilus XXVI, Crocodilus XXVII, EU496857,
EU496852 y EU496851 Venegas-Anaya et al., 2008) y como clado hermano a C. yacare
(JF315314). El clado C. c. crocodilus/C. c. apaporiensis contiene secuencias de individuos
identificados como C. c. crocodilus del centro y occidente de la amazonia (JF315306,
EU496858, EU496860, Crocodilus XXIX, Crocodilus XXXI y Crocodilus XXX) y uno identificado
a partir del paratipo identificado por Medem como C. c. apaporiensis (ICN8763, Medem,
1955) (Figuras 6, 7 y 8).
Figura 7. Patrones de divergencia intraespecifica de las secuencias de Citocromo B para el Genero Caiman.
Divergencias de las secuencias al interior de las subespecies dentro de la misma región geográfica (0 - 0,005)
Divergencias de las secuencias al interior de las subespecies de regiones geográficas adyacentes: C. c. fuscus (Sur de
Costa Rica, Panamá y Colombia) V.S. C. c. fuscus (Norte de Costa Rica y Nicaragua).(0,011-0,015)
Divergencias de las secuencias al interior de las subespecies de regiones adyacentes C. c. fuscus (Nicaragua, Costa Rica,
Panamá y Colombia) V.S. C. c. fuscus/C. c. chiapasius (México y Hondura). (0,018-=,022)
Divergencias de las secuencias al interior de las subespecies de región amazónica: C. c. crocodilus (Oriente) V.S. C. c. cro-
codilus/C. c. apaporiensis (Occidente). (0,038-0,042)

37
Figura 8. Patrones de divergencia interespecifica de las secuencias de Citocromo B para la Subfamilia Alliga-
torinae.

38
En el análisis de secuencias de COI y CytB en conjunto, el árbol de máxima verosimilitud
(Figura 9) obtuvo soportes mayores a 90% para la mayoría de los clados. En ésta hipótesis,
Alligator mississippiensis presenta divergencias del 18,4 al 19,1% (Figura 8) con respecto al
género Caiman y del 16,6% con respecto a Paleosuchus trigonatus (Figura 8), quien a su vez
diverge del género Caiman en un 15 a 15, 6% (Figura 8). De manera similar al análisis basado
únicamente en CytB, se recuperó Caiman crocodilus como monofilético con un soporte de
100%; y al interior de la especie se mantiene la misma estructura entre las hipótesis obte-
nidas con MP, IB y ML, es decir, se recuperan 7 Clados y la relación entre estos y el soporte
de las ramas es similar.
El primer clado que diverge dentro del género es Caiman crocodilus crocodilus (Oriente de
la Amazonia) con un soporte del 100% en el análisis ML. Esta división separa a C. c. crocodi-
lus del Oriente de la amazonia (Crocodilus XXVIII, Crocodilus XXV, Crocodilus XXIV, Crocodi-
lus XXVI, Crocodilus XXVII en Venegas-Anaya et al., 2008) del resto de clados del género. La
siguiente divergencia presenta un soporte de solo el 41%, y muestra dos divisiones, en una
división está C. c. crocodilus del Occidente de la amazonia (Crocodilus XXIX, Crocodilus XXXI
y Crocodilus XXX) y en la otra división se integran dos linajes, el primer linaje corresponde a
C. c. fuscus/C. c. chiapasius de México y Honduras (Chiapasius XX, Chiapasius XXI, Fuscus IV,
Fuscus V y Fuscus VI Venegas-Anaya et al. 2008), mientras que el segundo linaje comprende
a los clados C. c. fuscus ubicados de Nicaragua al norte de Costa Rica (Fuscus I, Fuscus II,
Fuscus III, Fuscus XV y Fuscus XVI Venegas–Anaya et al., 2008) y al clado C. c. fuscus del sur
de Costa Rica hasta Colombia (Fuscus VII, Fuscus VIII, Fuscus IX, Fuscus X, Fuscus XI, Fuscus
XII, Fuscus XIII, Fuscus XIV, Fuscus XVII, Fuscus XVIII, Fuscus XIX, Fuscus XXII, Fuscus XXIII
Venegas-Anaya et al. (2008), Cc 01 a Cc19 presente estudio) (Figura 9).

39
Alligator mississipiensis
Paleosuchus trigonatus
Caiman crocodilus fuscus (Sur de Costa
Rica hasta Colombia)
Caiman crocodilus fuscus (Nicaragua
al Norte de Costa Rica)
Caiman crocodilus chiapasius/Caiman crocodilus fuscus (Mé-
xico hasta Honduras)
Caiman crocodilus crocodilus (Oriente de la Amazonia)
Caiman crocodilus crocodilus (Occidente de la Amazonia)
Genero Caiman
Figura 9. Árbol de reconstrucción filogenética a partir del método de Máxima Verosimilitud (ML). Se muestran
en las ramas soportes correspondientes a bootstrap, resultados provenientes de los genes completos del CytB
y COI.
El análisis de parsimonia (Figura 10) arrojó 84 árboles igualmente parsimoniosos, con una
longitud de 317 pasos, Índice consistencia: 80, Índice de retención: 92; el árbol de consenso
estricto presenta una topología muy similar a la obtenida con el análisis de ML, en cuanto a
la relación entre los clados principales, pero la resolución por ejemplo al interior del clado

40
que tiene las muestras de Colombia es mayor con MP (Figura 9). En el clado Caiman croco-
dilus se presenta una división que separa a C. c. crocodilus del Oriente de la amazonia del
resto de los grupos de Caiman crocodilus con un soporte de solo el 53%. Adicionalmente,
presenta una bifurcación separando a C. c. fuscus del sur de Costa Rica hasta Colombia en
dos linajes, en el primero agrupa a algunos Caiman crocodilus de Colombia (Cc1 al Cc11) y
el Fuscus XIII (Muestra proveniente de la costa atlántica colombiana, Venegas–Anaya et al.,
2008). En el otro asocia a Caiman crocodilus de Colombia (Cc 12 al Cc 19) y otros haplotipos
hallados por Venegas-Anaya et al. (2008), provenientes de del sur de Costa Rica y Panamá
(Fuscus VII, Fuscus VIII, Fuscus IX, Fuscus X, Fuscus XI, Fuscus XII, Fuscus XIV, Fuscus XVII,
Fuscus XVIII, Fuscus XIX, Fuscus XXII, Fuscus XXIII).
El árbol obtenido mediante inferencia bayesiana (Figura 11), también tuvo una topología
similar al árbol obtenido por medio de Máxima Verosimilitud (Figura 9), pero a diferencia
de éste, el árbol obtenido mediante inferencia bayesiana presenta una politomía que
agrupa a cinco clados de Caiman crocodilus, que corresponden a Caiman crocodilus croco-
dilus al oriente de Sudamérica, Caiman crocodilus crocodilus al occidente de Sudamérica,
Caiman crocodilus chiapasius agrupado con Caiman crocodilus fuscus (México hasta Hon-
duras), Caiman crocodilus fuscus (Nicaragua al Norte de Costa Rica) y finalmente Caiman
crocodilus fuscus (Sur de Costa Rica hasta Colombia). Se registran soportes mayores a 90 en
los nodos.
Las distancias encontradas para este análisis actúan como un soporte adicional para las ra-
mas de los árboles, donde se pueden ver divergencias del 2,8 al 3,9% entre los clados C. c.
crocodilus frente a los C. c. fuscus y C. c. chiapasius, al igual que los linajes de C. c. crocodilus
del oriente de la amazonia y C. c. crocodilus del occidente de la amazonia. Los clados C. c.
fuscus de Nicaragua, Costa Rica, Panamá y Colombia, divergen del clado C. c. fuscus/C. c.
chiapasius de México y Honduras en un 1,6 a 2,1% (Figura 12). En el caso del linaje de C. c.
fuscus que se extiende desde Nicaragua a Colombia se encontró una divergencia del 1 al
1,4%, entre los clados de Nicaragua y norte de Costa Rica frente al clado que se extiende
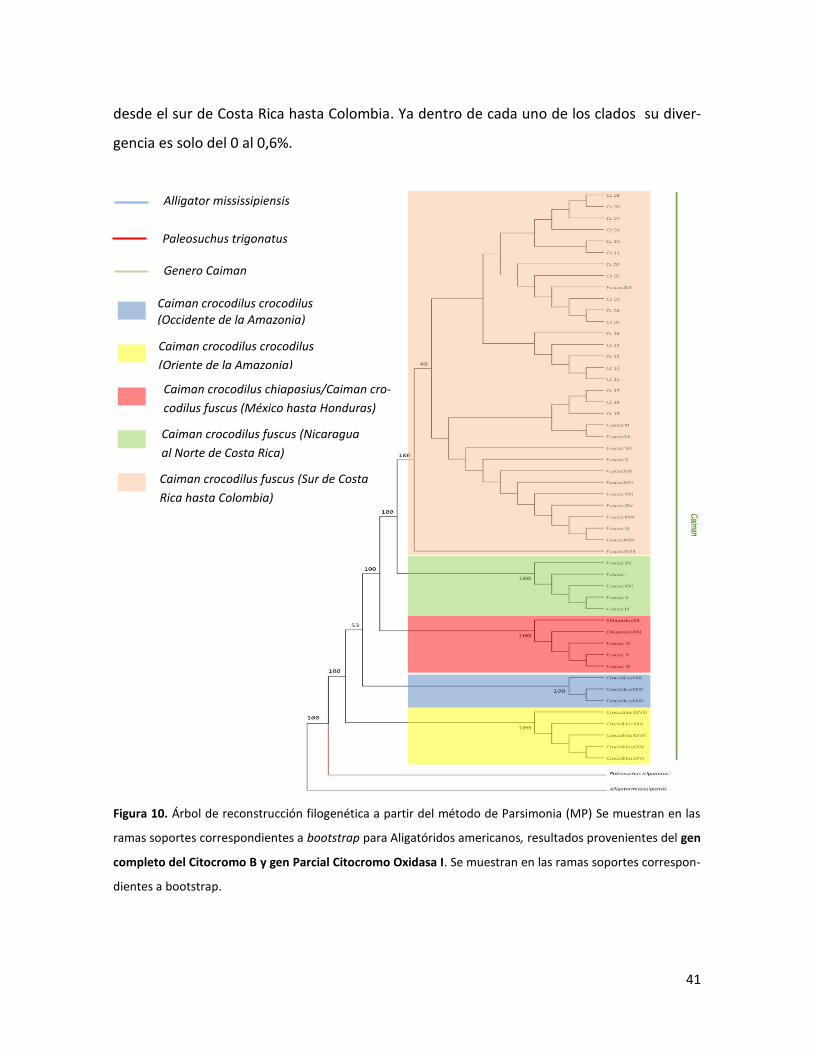
41
desde el sur de Costa Rica hasta Colombia. Ya dentro de cada uno de los clados su diver-
gencia es solo del 0 al 0,6%.
Figura 10. Árbol de reconstrucción filogenética a partir del método de Parsimonia (MP) Se muestran en las
ramas soportes correspondientes a bootstrap para Aligatóridos americanos, resultados provenientes del gen
completo del Citocromo B y gen Parcial Citocromo Oxidasa I. Se muestran en las ramas soportes correspon-
dientes a bootstrap.
Alligator mississipiensis
Paleosuchus trigonatus
Caiman crocodilus fuscus (Sur de Costa
Rica hasta Colombia)
Caiman crocodilus fuscus (Nicaragua
al Norte de Costa Rica)
Caiman crocodilus chiapasius/Caiman cro-
codilus fuscus (México hasta Honduras)
Caiman crocodilus crocodilus
(Oriente de la Amazonia)
Caiman crocodilus crocodilus (Occidente de la Amazonia)
Genero Caiman

42
Figura 11. Árbol de reconstrucción filogenética a partir del método de Inferencia Bayesiana (IB) para Aligató-
ridos americanos, resultados provenientes del gen completo del Citocromo B y gen Parcial Citocromo Oxi-
dasa I. Se muestran en las ramas soportes correspondientes a valores de la probabilidad posterior
Alligator mississipiensis
Paleosuchus trigonatus
Caiman crocodilus fuscus (Sur de Costa
Rica hasta Colombia)
Caiman crocodilus fuscus (Nicaragua
al Norte de Costa Rica)
Caiman crocodilus chiapasius/Caiman crocodilus fuscus (Mé-
xico hasta Honduras)
Caiman crocodilus crocodilus (Oriente de la Amazonia)
Caiman crocodilus crocodilus (Occidente de la Amazonia)
Genero Caiman

43
0
20
40
60
80
100
120
140
160
180
200
0,0
00
0,0
01
0,0
02
0,0
03
0,0
04
0,0
05
0,0
06
0,0
07
0,0
08
0,0
09
0,0
10
0,0
11
0,0
12
0,0
13
0,0
14
0,0
15
0,0
16
0,0
17
0,0
18
0,0
19
0,0
20
0,0
21
0,0
22
0,0
23
0,0
24
0,0
25
0,0
26
0,0
27
0,0
28
0,0
29
0,0
30
0,0
31
0,0
32
0,0
33
0,0
34
0,0
35
0,0
36
0,0
37
0,0
38
0,0
39
0
5
10
15
20
25
30
35
0,1
50
0,1
51
0,1
52
0,1
53
0,1
54
0,1
55
0,1
56
0,1
57
0,1
58
0,1
59
0,1
60
0,1
61
0,1
62
0,1
63
0,1
64
0,1
65
0,1
66
0,1
67
0,1
68
0,1
69
0,1
70
0,1
71
0,1
72
0,1
73
0,1
74
0,1
75
0,1
76
0,1
77
0,1
78
0,1
79
0,1
80
0,1
81
0,1
82
0,1
83
0,1
84
0,1
85
0,1
86
0,1
87
0,1
88
0,1
89
0,1
90
0,1
91
A
B
| 2Q
Figura 12. A. Patrones de divergencia intraespecifica de las secuencias de Citocromo B Completo y para el gen
parcial Citocromo oxidasa I para el Genero Caiman. B. Patrones de divergencia interespecifica de las secuen-
cias de Citocromo B Completo y para el gen parcial Citocromo oxidasa I para para la Subfamilia Alligatorinae.
Divergencias de las secuencias al interior de las subespecies dentro de la misma región geográfica (0 - 0,006)
Divergencias de las secuencias al interior de las subespecies de regiones geográficas adyacentes: C. c. fuscus (Sur de
Costa Rica, Panamá y Colombia) V.S. C. c. fuscus (Norte de Costa Rica y Nicaragua).(0,010-0,014)
Divergencias de las secuencias al interior de las subespecies de regiones adyacentes C. c. fuscus (Nicaragua, Costa Rica,
Panamá y Colombia) V.S. C. c. fuscus/C. c. chiapasius (México y Hondura). (0,016-=,021)
Divergencias de las secuencias al interior de las subespecies de región amazónica: C. c. crocodilus (Oriente) V.S. C. c. cro-
codilus (Occidente). (0,028-0,039)
Divergencias interespecifica entre Paleosuchus trigonatus con el género Caiman (0,150- 0,156)
Divergencias interespecifica entre Alligator mississipiensis con Paleosuchus trigonatus.(0,166)
Divergencias interespecifica entre Alligator mississipiensis con el géneros Caiman. (0,184 - 0,191).

44
Redes de haplotipos
Adicionalmente, en un análisis de redes basado en parsimonia (TCS, Clement et al. 2000),
la totalidad de las secuencias de caimanes distribuidos en Colombia y obtenidas en este
estudio, se agruparon con los haplotipos de individuos identificados por Venegas-Anaya et
al. (2008) como C. c. fuscus ubicados en el Sur de Costa Rica y Panamá. Estos se encuentran
separados por 18 pasos mutacionales del haplogrupo conformado por haplotipos de los in-
dividuos distribuidos en el Norte de Costa Rica y Nicaragua. Algo que se observa es que no
se ve ninguna conexión de estos haplogrupos con el conformado por C. c. fuscus /C. c.
chiapasius identificado por Venegas-Anaya et al. (2008) localizados en México y Honduras.
Estos linajes guardan la misma relación que se recupera en la construcción de los árboles.
Al igual que los dos linajes de C. c. crocodilus, las cuales no se encuentran conectadas a la
red de C. c. fuscus, ni tampoco entre sí (Figura 13).
Al realizar el agrupamiento de haplotipos según la metodología Nested Clade Analysis
(Templeton, 1998), se observa la recuperación de dos hablogrupos. El primer haplogrupo
agrupa los haplotipos Cc 01 al Cc 11, provenientes geográficamente de la Costa Atlántica,
Catatumbo, Alto Magdalena, Norte del Chocó. En el segundo haplogrupo se recuperan los
haplotipos Cc 12 al Cc 19, con localidades del Pacifico Colombiano, Urabá Antioqueño y la
zona del Magdalena medio. (Figuras 14 y 15).

45
Figura 13. Red de haplotipos de Caiman crocodilus, basada en los dos genes mitocondriales citocromo b y
citocromo oxidasa I. Los círculos representan los haplotipos encontrados de Caiman crocodilus.
Ca
ima
n cro
cod
ilus fu
scus C
osta
Ca
-
ribe, C
osta
Pa
cífica, C
ata
tum
bo
y
Va
lles intera
nd
ino
s de C
olo
mb
ia
Ca
ima
n cro
cod
ilus fu
scus
(Sur d
e Co
sta R
ica y P
a-
na
má
)
Ca
ima
n cro
cod
ilus fu
scus
(No
rte de C
osta
Rica
y
Nica
rag
ua
)
Ca
ima
n cro
cod
ilus
croco
dilu
s (Occid
ente
de la
Am
azo
nia
)
Ca
ima
n cro
cod
ilus ch
iap
a-
sius/C
aim
an
croco
dilu
s fus-
cus (M
éxico h
asta
Ho
nd
u-
ras)
Ca
ima
n cro
cod
ilus
croco
dilu
s (Orien
te
de la
Am
azo
nia
)

46
Figura 14. Agrupamiento de Haplotipos de Caiman crocodilus, según la metodología Nested Clade Analysis
(Templeton, 1998), basada en los dos genes mitocondriales citocromo b y citocromo oxidasa. Los círculos
representan los haplotipos, Los rectángulos naranja el nivel 1 de agrupamiento, los rectángulos azules el nivel
2 de agrupamiento y en verde el nivel 3 de agrupamiento.

47
Figura 15. Mapa de distribución de haplogrupos Caiman crocodilus fuscus según la construcción de redes
haplotípica.
Nivel de agrupación de
haplotipos
Haplogrupo I
Haplogrupo II

48
Tiempos de Divergencia
Los resultados del análisis del método de reloj molecular "relajado" basado en la estadística
Bayesiana son mostrados en la Figura 15. Para cada nodo encontrado en el árbol se obtuvo
un tiempo en millones de años (Ma). Se dataron los nodos del árbol de Inferencia Bayesiana
con PP≥ 95. En la hipótesis que se plantea que el ancestro de Caimaninae, data cerca de 13
Ma, y con una ubicación en Sudamérica y el cual se encontraba aislado en la zona de la
Amazonia por el sistema de aguas lacustres Pebas y a medida que este se fue retirando, se
inicia el proceso de colonización o de irradiación el cual dio como origen al ancestro del
genero Caiman hace cerca de 5,4 Ma.
De acuerdo con los resultados, el ancestro del género Caiman, posiblemente se dispersó
hacia el norte y sur de Sudamérica en donde el siguiente evento de vicarianza data hace 5,4
Ma cuando divergieron dos linajes ancestrales, el primer lianje originó a Caiman latirostris
y el segundo originó a Caiman yacare y Caiman crocodilus. El evento más relevante que
puede generar este proceso de especiación es el drenaje de lago Acre. Hace 2,27 Ma termi-
nado el drenaje de Acre se ve un proceso de divergencia entre los linajes de C. c. apaporien-
sis y C. c. crocodilus del occidente amazónico de los linajes ancestrales de C. yacare y C. c.
crocodilus del oriente amazónico. La divergencia de los linajes de C. yacare y C. c. crocodilus
del oriente amazónico data hace 1,53 Ma.
El siguiente evento de vicarianza data en 2,7 Ma, el cual separa los ancestros de Caiman
crocodilus que se desplazaron hacia el norte dado por el levantamiento de los andes, este
evento separo los linajes amazonicos de los linajes ancestrales de Caiman crocodilus fuscus
y Caiman crocodilus chiapasius. En el proceso de dispersión del género Caiman se desplaza
hacia Centro América y se genera un proceso de vicarianza hace 1,15 Ma que separa los
linajes ancestrales de C. c. fuscus y C. c. chiapasius.

49
Figura 16. Tiempos de divergencia en millones de años (Ma), de especies de Crocodilidos.
Para el análisis de reloj molecular se puede ver que la aproximación realizada en el presente
estudio presenta un grado de calibración mucho más robusto que el presentado en el aná-
lisis realizado por Venegas – Anaya et al. (2011), en el cual no se describen los puntos de
calibración. De otro lado y de manera contrastante los tiempos de divergencia para los
grupos Caimaninea y Paleosuchus en este estudio son mas recientes que los decritos por
Venegas et al. (2008). En la medida que este estudio proporciona diferentes puntos de
calibración y una metodología clara, se considera que los tiempos presentados en este es-
tudio son más robustamente sustentados y en consecuencia más recomendables para des-
cribir los procesos evolutivos en el grupo.

50
DISCUSION
De acuerdo con la información aportada por los árboles filogenéticos se puede ver que en
el género Caiman y las relaciones evolutivas de sus especies se mantienen similares utili-
zando tanto el gen CytB en solitario como cuando se usa en conjunto los genes CytB y COI.
Lo que demuestra que el comportamiento evolutivo de los dos marcadores es similar, si
bien la mayor parte de la información que permite generar la hipótesis es aportada por el
CytB. Estas relaciones son coherentes con la información aportada por el análisis de distan-
cias p y es congruente con la red haplotípica, pues en ambos casos se identifican los mismos
clados o haplogrupos.
Diversidad de haplotipos
En el muestreo realizado en las regiones de la Costa Pacífica, Costa atlántica, Valle del Mag-
dalena y Catatumbo Colombiano, se recuperaron 19 haplotipos de los genes mitocondriales
CytB y COI a partir de 78 individuos. De acuerdo con las curvas de acumulación, este mues-
treo representa algo así como el 50% de los haplotipos que posiblemente se podrían encon-
trar en las áreas muestreadas. Esto muestra una posibilidad de encontrar cerca de 38 ha-
plotipos posibles para el Valle del Magdalena, Catatumbo y la Costa Pacífica y Atlántica Co-
lombiana. Esto pone en contexto el número de haplotipos reportados por Venegas-Anaya
et al. (2008), quienes encontraron 23 haplotipos para la zona de Meso y Centro America. Al
no conocer la intensidad de muestreo de Venegas-Anaya et al. (2008), ni su proceso de
colecta no se pueden comparar directamentamente los dos estudios, por lo tanto, no se
puede decir si hay una mayor diversidad haplotípica en Colombia que en Centro y Meso
America.

51
Relaciones filogenéticas entre el grupo Caiman crocodilus con respecto a Aligatóridos
americanos.
En las hipótesis obtenidas con el gen completo CytB, el género Caiman se recupera como
monofilético y como su clado hermano encontramos al género Melanosuchus (Groom-
bridge, 1987; King y Burke, 1989; Hrbek et al., 2008), con un tiempo de divergencia de 8,4
Ma. En el presente estudio, el grupo C. c. fuscus/chiapasius se compone de tres subclados
que corresponden a: (1) Caiman crocodilus fuscus / Caiman crocodilus chiapasius en México
a Honduras, (2) Caiman crocodilus fuscus de Nicaragua al Norte de Costa Rica y (3) Caiman
crocodilus fuscus del Sur de Costa Rica a Colombia (Costa Pacífica, Costa Atlántica, Valle del
Magdalena y Catatumbo; incluidos los paratipos de la Isla Gorgona reportados por Medem,
1962). Algunos de estos hallazgos concuerdan con lo reportado por Venegas-Anaya et al.
(2008), en los cuales se ve la asociación de las muestras de Colombia al Clado del Sur de
Costa Rica y Panama.
La estructura del árbol filogenético muestra una gran separación entre los linajes de Caiman
de Mesoamerica y Amazonia, quienes presentan tiempos de divergencia de 2,7 Ma. En el
estudio de Venegas-Anaya et al. (2008), reporta divergencias de estos dos linajes de 5,7 a
6,7 millones de años (Ma). Aunque estos resultados difieren en cerca de 3 a 4 Ma entre sí
son concordantes con el período de máximo levantamiento de la Cordillera de los Andes
(Gregory-Wodzicki, 2000), el cual sugiere que la altura de la Cordillera de los Andes del
Norte se incrementó rápidamente durante el Plioceno, entre 2 y 5 Ma a tasas de un orden
de 0,5-3,0 mm/año, y alcanzó las elevaciones modernas alrededor de 2,7 Ma. Este periodo
temporal de diversificación en el noroccidente de América del Sur, es coincidente con los
datos inferidos por Eberhard y Bermingham (2004) para el complejo de loros Amazona och-
rocephala y por Cheviron et al. (2005) para las aves de bosques de tierras bajas, Lepidothrix
coronata y se encuentran asociados a las incursiones marinas ocurridas hacia el Pleistoceno.

52
En el caso de la diversificación de C. c. crocodilus en dos clados, uno distribuido al Oriente
amazónico y otro en el Occidente amazónico, y que corresponden a los linajes 4 y 5 en el
estudio de Venegas –Anaya et al. (2008), se estimó una divergencia menor en este estudio
(2,27 Ma). Estos patrones de divergencia pueden estar asociados a cambios en los niveles
de ríos y fuentes hídricas especialmente procesos vicariantes como la aparición y drenaje
del Sistema Pebas y la conformación del río Amazonas en el Mioceno medio y tardío (Ribas
et al., 2012; Solomon et al., 2008; Llorente-Bousquets y Morrone, 2001; Van der Hammen,
2000). En este proceso las alteraciones temporales y espaciales entre sistemas de agua sa-
lada y dulce ocasionaron una historia dinámica y diversa para diferentes áreas geográficas,
por lo que pudo generar un aislamiento de poblaciones ancestrales a los dos clados (Ribas
et al., 2012; Solomon et al., 2008; Llorente-Bousquets y Morrone, 2001; Van der Hammen,
2000).
Relación filogenética de los caimanes (C. crocodilus) del pacifico y atlántico colombiano
con ejemplares de Mesoamérica
Al analizar la estructura del árbol filogenético de los Aligatóridos americanos se encuentra
una división de los clados de Caiman crocodilus en Centroamérica, asociada a barreras geo-
gráficas presentes en la zona. Esta región separa dos masas de tierras altas, las tierras altas
del bloque Chortis al norte (Honduras y Nicaragua) y las tierras altas de la porción baja de
América Central (Costa Rica y Panamá). La evidencia geológica sugiere que existió una se-
paración marina entre las tierras altas de Chortis y la porción baja de América Central du-
rante el Plioceno (Coates y Obando, 1996; Iturralde-Vinent, 2006). La Depresión de Nicara-
gua ha sido identificada como una de las mayores causas de separación filogeográfica para
taxones como ranas (Savage, 1987; Duellman, 1999), salamandras (Parra-Olea et al., 2004),
serpientes (Savage, 1982; Cadle, 1985), plantas, insectos y peces (Marshall y Liebherr, 2000;
Halas et al., 2005). Esto puede llegar a explicar la separación de los linajes de C. c. fuscus/C.
c. chiapasius localizados desde el Sur de México hasta Honduras, de las otras poblaciones

53
de C. crocodilus de Centro y Sudamérica, los cuales alcanzan tiempos de divergencia de 1,15
Ma en el presente estudio, frente a los 2,5 a 2,9 Ma reportados por Venegas-Anaya et al.
(2008).
Los linajes C. c. fuscus localizados desde Nicaragua hasta el Norte de Costa Rica y el corres-
pondiente al Pacífico de Costa Rica, Panamá y Colombia están separados por cordilleras con
alturas aproximadas entre 1000 y 2000 m (Cordilleras de Guanacaste, Central y Talamanca
en Costa Rica). Estudios realizados en diversos organismos (anfibios: García-París et al.,
2000; Wang et al., 2008; reptiles: Saldarriaga-Cordoba et al., 2009; Zamudio y Greene, 1997;
Parkinson et al., 2000; peces de agua dulce: Bermingham y Martín, 1998; Mamíferos: De-
mastes et al., 1996; Cropp y Boinski, 2000; Harris et al., 2000; árboles: Cavers et al., 2003;
Novick et al., 2003), han demostrado que el patrón filogeográfico de estos taxones dentro
de Mesoamérica ha sido fuertemente influenciado por la historia geológica compleja de
esta región, por los cambios cíclicos en el clima, la vegetación y el nivel del mar, además de
los procesos orogénicos, generando procesos de vicarianza.
Durante el Plioceno con el cierre del Istmo de Panamá, hace aproximadamente 3.5 a 3.0
m.a (Coates y Obando, 1996), permitió el intercambio entre la biota de América Central y
América del Sur, favoreciendo la dispersión a gran escala de la fauna terrestre, fenómeno
conocido como "El Gran Intercambio Biótico Americano (GABI) (Coates y Obando, 1996;
Webb y Rancy, 1996). Un aspecto interesante de la biogeografía Neotropical es la posible
dispersión entre América Central y América del Sur, durante periodos de separación marina
(pre-ístmico) de múltiples grupos taxonómicos, principalmente anfibios y reptiles (Hanken
y Wake, 1982; Estes y Baez, 1985; Vanzolini y Heyer, 1985; Cadle, 1985; Bermingham y Mar-
tin, 1998; Weigt et al., 2005; Wang et al., 2008). La dispersión de los reptiles entre América
Central y América del Sur probablemente ocurrió a través de la cadena de islas que confor-
maban el área que hoy ocupa la porción baja de América Central (Zamudio y Greene, 1997;
Saldarriaga-Cordoba et al., 2009).

54
Relación genética de los caimanes (C. crocodilus) de la costa atlántica y pacífica en Co-
lombia.
De acuerdo con la curva de acumulación de haplotipos (Figura 4) se logró recuperar el 50%
de los haplotipos esperados para la Costa Pacífica y Atlántica colombiana, Valle del Magda-
lena y Catatumbo, encontrando buena parte de la diversidad de la especie Caiman crocodi-
lus en las zonas muestreadas. En consecuencia, debido a que están sustentadas en un mues-
treo apropiado se hacen diferentes consideraciones sobre los posibles escenarios en los que
pudo desarrollarse la aparición en este territorio, de los diferentes linajes del género Cai-
man mediante un análisis de redes de haplotipos.
De acuerdo con los análisis de este trabajo, individuos de la especie C. crocodilus distribui-
dos tanto en el Pacífico y en el Atlántico colombianos, así como en el Catatumbo y el Valle
del Magdalena, corresponden a la subespecie C. c. fuscus, identificada en el trabajo de Ve-
negas-Anaya et al. (2008) como el clado “Sur de Centro América” (linaje II). En consecuencia,
los haplotipos de esta subespecie, distribuidos en Colombia, son topológicamente herma-
nos de un clado centroamericano (descrito como linaje I) según Venegas-Anaya et al.
(2008).
En el análisis de redes de haplotipos se puede observar que los haplotipos de individuos
colectados en el presente estudio forman dos haplogrupos, cercanamente relacionadas con
haplotipos provenientes del sur de Centro América. Dos de estos haplogrupos se distribu-
yen en el corredor biogeográfico que forma parte de la Eco-Región Chocó-Darién, la cual se
extiende desde el oriente de Panamá en la provincia del Darién y comarca de Kuna-Yala
hasta el Cabo Pasado en la provincia de Manabí en el norte de Ecuador (WWF, Fondo Mun-
dial para la naturaleza, Dinerstein et al., 1995; Saldarriaga-Cordoba et al., 2009). El primero
agrupa los haplotipos Cc 17 a Cc 19, provenientes de Centro y Sur del Pacifico Colombiano
y Puerto Berrio, Antioquia. El segundo haplotipo se encuentra ubicado al Norte del Pacifico
y Arboletes, Antioquia y agrupa los haplotipos cc 12 a Cc16. El otro se extiende por la costa

55
atlántica, los valles interandinos del Magdalena y Catatumbo, es el haplogrupo con mayor
diversidad haplotípica la cuales comprende los haplotipos, Cc 01 al Cc 11, este se encuentra
enmarcada por las estribaciones de las tres Cordilleras de los Andes, y éstas quizá limitan la
dispersión de las poblaciones desde esta región hacia la Vertiente Pacífica.
Teniendo en cuenta el análisis de redes de haplotipos se puede observar que los tres ha-
plogrupos encontrados en Colombia se agruparon (compartiendo el haplotipo ancestral)
dentro del clado más al sur de C. c. fuscus del sur de America Central según lo reportado
por Venegas-Anaya et al. (2008). El patrón de diversidad haplotipica dentro de C. c. fuscus
indica que las poblaciones actuales de C. c. fuscus en Colombia probablemente son el resul-
tado de una expansión historica reciente de un único linaje del sur de America Central.
En contraste y comparando los tiempos de divergencia hallados con el reloj molecular, se
puede observar Caimaninea, presentando como fósil más antiguo del grupo a Kuttanacai-
man iquitosensis, cuyo origen es suramericano, ampliando su distribución hacia el norte y
sur del continente con diferentes procesos de vicarianza que originaron diversos linajes evo-
lutivos, llegando a los ancestros de Caiman, que cruzaron el puente formado por el Itsmo
de Panamá y aumentando su areal de distribución más al norte de Centro América. Esto
sugiere un proceso de extinción local en el área del Caribe, Pacifico, Catatumbo y Valle del
Magdalena, y luego una re-colonización desde Centro América, dado el patron de compor-
tamiento de los haplotipos y viendo su origen a partir de haplotipos Centroamericanos.
Dentro de esta hipótesis se pueden plantear algunos eventos que pudieran generar la desa-
parición de los linajes ancestrales dentro de las zonas mencionadas; uno de ellos puede
asociarse a las incursiones marinas del norte de Suramérica que generaron ecosistemas
acuáticos de aguas saladas en los cuales el género Caiman no puede sobrevivir (Fritss 1998;
McCoid y Kleberg 1995; Forero-Medina et al., 2006), aislándolos o desplazándolos hacia
Centro America. Dada la topografía de hoy en día, un aumento del nivel del mar de 100 m

56
habría producido incursiones marinas del mar Caribe (a lo largo de la costa norte de Colom-
bia y Venezuela) y desde el Océano Pacífico (a lo largo de la costa occidental de Panamá,
Colombia, Ecuador y Perú) durante las fases cálidas del Plioceno (Haq et al., 1987) y el Pleis-
toceno (Nores, 2004)
Varias áreas de bosques tropicales tendrían que ser aislados en: (1) las colinas y tierras bajas
adyacentes del Darién, San Blas, SAPO y las montañas Baudó; (2) las estribaciones occiden-
tales y las tierras bajas adyacentes de los Andes Occidentales; (3) las estribaciones del norte
y las tierras bajas adyacentes de la cordilleras Occidental y Central de Colombia, y el Valle
del Magdalena Medio; (4) las estribaciones orientales de la montaña y el Perijá tierras bajas
de Maracaibo; y (5) los barrancos, laderas y tierras bajas adyacentes de las cordilleras Chon-
gón Colonche y al mismo tiempo, varias áreas no forestales también habrían sido aislados
por transgresiones marinas en el norte de Colombia y Venezuela y el noroeste de Perú: (a)
las montañas de la Península de la Guajira; (B) la base de la Sierra de Santa Marta; (E) de la
Serranía de San Jacinto; (D) estribaciones septentrionales de los Andes; (E) Foca; y (f) Punta
Aguja (Nores, 2004).
Identidad taxonómica de las poblaciones trans Andinas de Caiman crocodilus en Colombia
En concordancia con las opiniones de Venegas-Anaya et al. (2008) y Escobero-Galván et al.
(2011), - quienes limitan a C. c. chiapasius únicamente a Mesoamérica, y de acuerdo con
Busack y Pandya (2001), los cuales infieren estadísticamente que las diferencias en los ca-
racteres morfológicos propuestos para la delimitación de la subespecie, respecto a las otras
subespecies del género?, no son significativas -, los resultados encontrados en los diferen-
tes análisis en este trabajo muestran que las poblaciones provenientes de la Isla Gorgona y
del Pacifico Colombiano identificadas por Medem (1962) como C. c. chiapasius, no están
relacionadas con la subespecie C. c. chiapasius de Centroamérica. Estas poblaciones esta-
rían relacionadas con el clado C. c. fuscus de Sudamérica y Panamá, al igual que todas las

57
muestras colectadas en el presente estudio, no pueden asignarse a la sub especie C. c.
chiapasius.
Por lo tanto puede concluirse que las secuencias obtenidas a partir de los cráneos de espe-
címenes identificados por Medem (1962), como C. c. chiapasius, ubicados en la colección
del Instituto de Ciencias Naturales de la Universidad Nacional de Colombia (ICN8699,
ICN8707) y de individuos muestreados a lo largo de las zonas de Pacifico reconocidas por el
mismo autor como localidades de la subespecie en Colombia, están asociados a un linaje de
ubicado en Panamá y Costa Rica (identificado a partir de los datos de Venegas-Anaya et al.
(2008)) y que se encuentra muy alejado del Clado de C. c. chiapasius de México.
Implicaciones en conservación
Mientras que la distribución de la variación geográfica dentro y entre los taxones puede
proveer información sobre los procesos tanto históricos como contemporáneos de la de-
mografía y la evolución de las poblaciones muestreadas, estos datos también pueden ser
valiosos para su conservación en tanto que son útiles para definir las unidades de conser-
vación y determinar las acciones de manejo (Moritz 1994, Crandall et al. 2000, Frankham et
al. 2002). Es relevante decir que las diferentes poblaciones de una misma especie pueden
evidenciar una diferenciación genética tan significativa, que se puede justificar perfecta-
mente un programa de conservación a nivel de población, sin que sean consideradas como
especies distintas, pero si como unidades de conservación distintas (Moritz 1994, Crandall
et al. 2000, Frankham et al. 2002). Entonces, la delimitación apropiada de esas unidades de
conservación, requiere de herramientas como la genética de poblaciones y la sistemática
molecular (Moritz 1994, Crandall et al. 2000, Frankham et al. 2002) (REF). Las unidades de
conservación descritas, pese a que tengan una historia común “muy” reciente, deben tener
programas de manejo independientes para preservar la diversidad biológica y el potencial
evolutivo de la especie. Aunque podemos ver una estructura de tres agrupaciones en las

58
poblaciones de Caiman crocodilus fuscus en la región Pacifica y Atlántica, Valle del Magda-
lena y el Catatumbo Colombiano, estas están estrechamente ligadas a las poblaciones de
Centro América lo cual sugiere que para Colombia, se debe hablar de una sola unidad evo-
lutivamente significativa de C. crocodilus fuscus (Ryder 1986; Vogler & DeSalle 1994).
La identificación de tres haplogrupos de C. c. fuscus en Colombia, genera una herramienta
que permite la toma de decisiones al momento de delimitar áreas de liberación o reubica-
ción de especímenes, y al igual apoya la delimitación de las zonas de extracción de pies
parentales para procesos de zoocría o zonas de extracción de pieles.

59
Conclusiones
1. Las poblaciones de Caiman crocodilus distribuidas al oriente de los Andes en Colom-
bia, en las regiones del Caribe, Pacifico, Catatumbo y Valle del Magdalena, están
relacionadas con poblaciones del Sur de Costa Rica y Panamá, las cuales correspon-
den a la subespecie C. c. fuscus.
2. Se recuperan dos linajes de haplotipos asociados a distintas regiones geográficas en
Colombia ((1) Costa Atlántica, Catatumbo, Alto Magdalena, Norte del Chocó. (2) Pa-
cifico Colombiano, Urabá Antioqueño y la zona del Magdalena medio). las cuales
deben tenerse en cuenta para los planes de manejo de la especie.
3. Se reconoce que para la región trans Andina en Colombia solo se encuentra una
única subespecie, la cual está estrechamente relacionada con poblaciones del Sur
de Costa Rica y Panamá y que corresponden a la subespecie C. c. fuscus, por tanto
para el país se excluiría la especie C. c. chiapasius, y se reconocería que la subespecie
presente es C. c. fuscus.
4. Nuevos marcadores moleculares y una ampliación del muestreo en la zona de con-
fuencia de los dos hapologrupos encuntrados podrían ayudar en la comprensión de
los procesos demográficos y determinar zonas de confluencia de este grupo.

60
BIBLIOGRAFIA
Adam L. Bazinet, Derrick J. Zwickl, and Michael P. Cummings. 2014. A Gateway for Phylo-
genetic Analysis Powered by Grid Computing Featuring GARLI 2.0 Syst Biol first published
online April 30, 2014 doi:10.1093/sysbio/syu031
Aguilera O, Riff D, Bocquentin-Villanueva J. 2006. A new giant Purussaurus (Crocodyli-
formes, Alligatoridae) from the upper Miocene urumaco formation, Venezuela. J Syst
Palaeontol 4:221–232.
Amato, G. y J. Gatesy. 1994. PCR assays of variable nucleotide sites for identification of con-
servation units. p. 215-226. En: Molecular ecology and evolution: Approaches and applica-
tions. B. Schierwater, B. Streit, G.P.Wagner and R. DeSalle (Eds.). Birkhauser, Basel, Switzer-
land.
Avise, J.C. 1994. Molecular Markers, Natural History, and Evolution. Chapman & Hall, New
York
Avise, J.C. 1998. The history and purview of phylogeography: a personal reflection. Molec-
ular Ecology. 7: 371-379.
Avise, J.C. 2000. Phylogeography, the history and formation of species. Harvard University
Press, Cambridge, EUA.
Barrios, L. M. y López-Victoria, M. 2001. Gorgona marina: Contribución al conocimiento de
una isla única. INVEMAR, Serie de Publicaciones especiales N° 7. Santa Marta, Colombia.

61
Bermingham, E., Martin, A.P., 1998. Comparative mtDNA phylogeography of Neotropical
freshwater fishes: testing shared history to infer the evolutionary landscape of lower Cen-
tral America. Mol. Ecol. 7, 499-517.
Bocourt, M.F. 1876 Note sur quelques reptiles de l’isthme de Tehuantepec (Mexique)
donnés par M. Sumichrast au Muséum. Journal de Zoologie, Paris, 5, 386–411.
Boom, R., Sol, C.J., Salimans, M.M., Jansen, C.L., Wertheim-Van Dillen, P.M., y Van Der Noor-
daa, J. 1990. Rapid and simple method for purification of nucleic acids. J. Clin. Microbiol. 28,
495- 503.
Boulenger, G.A. 1889. Catalogue of the Chelonians, Rhynchocephalians, and Crocodiles in
the British Museum (Natural History). London: Printed by order the Trustees, 8, 298 pp.
Briggs AW, Stenzel U, Johnson PLF, Green RE, Kelso J, Prüfer K, Meyer M, Krause J, Ronan
MT, Lachmann M, et al. 2007. Patterns of damage in genomic DNA sequences from a Nean-
dertal. Proc Natl Acad Sci 104: 14616–14621
Brochu, C.A., 2000. Phylogenetic relationships and divergence timing of Crocodylus based
on morphology and the fossil record. Copeia, 2000:657-673.
Brochu C.A, 2004. Alligatorine phylogeny and the status of Allognathosuchus Mook, 1921. J
Vert Paleontol 24:857–873.
Brochu, C. A., J. Njau, R. J. Blumenschine, and L. D. Densmore, 2010. A newhornedcroco-
dilefromthePlio-PleistocenehominidsitesatOlduvai Gorge, Tanzania. PLoS One 5:e9333
Busack SD y Pandya S. 2001. Geographic variation in Caiman crocodilus and Caiman yacare
(Crocodylia: Alligatoridae): systematics and legal implications. Herpetologica 57: 294–312.

62
Cadle, J.E., 1985. The neotropical colubrid snake fauna (Serpentes: Colubridae): lineage
components and biogeography. Syst. Zool. 34, 1-20.
Cavers, S., Navarro, C., Lowe, A.J., 2003. Chloroplast DNA phylogeography reveals coloniza-
tion history of a Neotropical tree, Cedrela odorata L., in Mesoamerica. Mol. Ecol. 12, 1451-
1460.
Cheviron, Z.A., Hackett , S.J., Capparella, A.P., 2005. Complex evolutionary history of a Ne-
otropical lowland forest bird (Lepidothrix coronata) and its implications for historical hy-
potheses of the origin of Neotropical avian diversity. Mol. Phyl. Evol. 36, 338-357.
Chirivi, H. G. 1971. Notas sobre el manejo de los Crocodylia en Colombia con especial refe-
rencia a la babilla y la factibilidad de su cría en cautiverio. INDERENA.
Clement, M., Posada. D., Crandall KD. 2000. TCS: a computer programe to estimate gene
genealogies. Molecular Ecology 9 (10): 1657-1660.
Coates, A.G., Obando, J.A., 1996. The geological evolution of the Central American Isthmus.
In: Jackson J.B.C., Budd. A.F., Coates, A.G. (Eds.), Evolution and Environment in Tropical
América. University of Chicago Press, Chicago. IL, pp. 21-56.
Colwell, R. K. 2013. EstimateS: Statistical estimation of species richness and shared species
from samples. Version 9. User's Guide and application published at: http://purl.oclc.org/es-
timates.
Cope, E.D. 1868. On the crocodilian genus Perosuchus. Proceedings of the Academy of Nat-
ural Sciences of Philadelphia, 1, 203 pp.

63
Crandall, K. A., O. R. P. Bininda-Emonds, G. M. Mace y R. K. Wayne. 2000. Considering evo-
lutionary processes in conservation biology. Trends in Ecology & Evolution 15: 290-295.
Cropp, S., Boinski, S., 2000. The Central American squirrel monkey (Saimiri oesterdii): intro-
duced hybrid or endemic species?. Mol. Phylogenet. E vol. 16, 350-365.
Darriba D, Taboada GL, Doallo R and Posada D. 2012. "jModelTest 2: more models, new
heuristics and parallel computing". Nature Methods 9, 772
DeSalle R y Amato G. 2004. The expansion of conservation genetics. Nature Reviews Genet-
ics 5, 702-712.
Demastes J, W., Hafner, M.S., Hafner, D.J., 1996. Phylogeographic variation in two Central
American pocket gophers (Orthogeomys). J. Mammal. 77, 917- 927.
Dinerstein, E., Olson, D., Graham, D., Webster, A., Primm, S., Bookbinder, M., Ledec, G.,
1995. Una evaluación del estado de conservación de las ecorregiones terrestre de América
Latina y El Caribe. WWF - Banco Mundial.
Washington, D .C.
Duellman, W.E., 1999. Distribution patterns of amphibians in South America. Patterns of
distribution of amphibians: a global perspective, Johns Hopkins University Press, Baltimore,
MD., pp. 255-328.
Drummond, A. J., Suchard, M. A., Xie, D., & Rambaut, A. 2012. Bayesian phylogenetics with
BEAUti and the BEAST 1.7. Mol. Biol. Evol. http://dx.doi.org/10.1093/molbev/mss075
Eberhard, J.R., Bermingham, E., 2004. Phylogeny and Biogeography of the Amazona ochro-
cephala (Aves: Psitacidae) complex. The Auk 121, 318-332.

64
Eguiarte Luis E., V. Souza y X. Aguirre (Compiladores). 2007. Ecología molecular. Semanrant,
Conabio, Inst. de Ecología UNAM. D. F., México.
Eizirik E, Bonatto S, Johnson W, Crawshw P, Vie JC, Brousset D, O`Brien SJ, Salzano F. 1998.
Phylogeographic patterns and Evolution of the Mitochondrial DNA Control Region in two
Neotropical Cats (Mammalia, Felidae). Molecular Evolution. 47: 613-624.
Escobero-Galvan, A, Cupul-Magaña, F y Velasco, J. 2011. Misconceptions about the taxon-
omy and distribution of Caiman crocodilus chiapasius and C. crocodilus fuscus (Reptilia:
Crocodylia: Alligatoridae). Zootaxa 3015: 66–68
Estes, R., Baez, A., 1985. Herpetofaunas of North and South America during the late Creat-
aceous and Cenozoic: evidence for interchange? In: Stehli, F.G., Webb, S.D. (Eds.), The Great
American Biotic Interchange. New York: Plenum Press, 139-197
Felsenstein, J. 2004. Inferring Phylogenies. Sinauer Associates, Sunderland, Massachusetts.
664 pp.
Forero-Medina, G., Castaño-Mora, O. y Rodriguez-Melo, M. 2006. Ecología de Caiman cro-
codilus fuscus en San Andrés Isla, Colombia: Un estudio preliminar. Caldasia 28(1):115-124. 2
Frankham, R., J. D. Ballou y D. A. Briscoe. 2002. Introduction to conservation genetics. Cam-
bridge University Press, United States of America, New York. 617 pp.
Fritts, T. H. 1988. The brown tree snake, Boiga irregularis, a threat to Pacific Islands. U.S.
Fish and Wildlife Service. Biological Report 88 (31): 1-36.

65
García-París, M., Good, D.A., Parra-Olea, G., Wake, D.B., 2000. Biodiversity of Costa Rican
salarnanders: irnplications of high levels of genetic differentiation and phylogeographic
structure for species forrnation. Proc. Natl. Acad. Sci. U.S.A. 97, 1640- 1647.
Genbank. http://www.ncbi.nlm.nih.gov/genbank/
Goloboff, P. 1999. NONA (NO NAME) ver. 2 Published by the author, Tucumán, Argentina.
Gregory-Wodzicki, KM., 2000. Uplift history of the central and northern Andes a review.
Geol. Soc. Am. Bull. 112, 1091-1105
Groombridge, B. 1987. The distribution and status of world crocodilian. En Grahme J. W.
Webb, S. Charlie Manolis, Peter Whitehead, Surrey Beatty and Son in Association with The
Conservation Comission of the Northen Territory (eds) Wildlife Management. Crocodiles
and Alligators. Australia.
Halas, D., Zamparo, D., Brooks, D.R., 2005. A historical biogeographical protocol for studying
biotic diversification by taxon pulses. J. Biogeogr. 32, 249-260.
Hall T.A. 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis
program for Windows 95/98/NT. Nucleic Acids Symposium Series, 41, 95-98.
Haq, B.U., Hardenbol, J., Vail, P.R., 1987. Chronology of fluctuating sea levels since the Tri-
assic. Science, 235, 1156- 1167.
Hanken, J.,Wake, D.B,. 1982. Genetic differentiation among plethodontid salamanders (ge-
nus Bolitoglossa) in Central and South America: implications for the South American inva-
sion. Herpetologica 38, 272-287.

66
Harris, D., Rogers, D.S., Sullivan, J., 2000. Phylogeography of Peromyscus furvus (Rodentia;
Muridae) based on cytochrorne b sequence data. Molecular Ecology, 9, 2129- 2135.
Hrbek, T., Vasconcelos, W., Rebelo, G. y Farias I. 2008 Phylogenetic Relationships of South
American Alligatorids and the Caiman of Madeira River. Journal of Experimental Zoology
309a:588–599
Ho, S.Y.W. and Phillips, M.J. 2009. Accounting for calibration uncertainty in phylogenetic
estimation of evolutionary divergence times. Systematic Biology, 58:367-380
Huelsenbeck JP, Ronquist F. 2001. MRBAYES: Bayesian inference of phylogenetic trees. Bi-
oinformatics. 17:754–755
Huelsenbeck, J. & Crandall, K. 1997. Phylogeny estimation and hypothesis testing using max-
imum likelihood. Annu. Rev. Ecol. Syst., 28: 437-466.
Iturralde-Vinent, M.A., 2006. Meso-Cenozoic Caribbean paleogeography: implications for
the historical biogeography of the region. Int. Geol. Rev. 48, 791-827.
Kemp GJ, Boning D, Beneke R, Maassen N. 2006. Explaining pH change in exercising muscle:
lactic acid, proton consumption, and buffering vs. strong ion difference. Am J Physiol Regul
Integr Comp Physiol.2006; 291:R235–R237. Author reply R238–R239.
Kessing B, Croom H, Martin A, McIntosh C, McMillan W, Palumbi SP. 1989. The simple fool’s
guide to PCR, version 1.0. Honolulu: Special Publication of the Department of Zoology, Uni-
versity of Hawaii.
King F. W., Burke R. L. 1989. Crocodilian, Tuatara and Turtle Species of the World. A taxo-
nomic and geographic reference. Assoc. Sistematic Collection, Washington, D.C.

67
Lanteri, A. & Confalonieri, V. 2003. Filogeografía: Objetivos, Métodos y Ejemplos. 185-193.
En: Bousquets, J. & Morrone, J. (eds.) Una Perspectiva Latinoamericana de la Biogeografía.
Facultad de Ciencias, UNAM, México.
Larriera A, Webb G., Velasco A., Rodriguez M. y Ortiz B. 2004. Informe Final Visita Colombia.
IUCN-SSC Crocodile Specialist Group.
Lemey, P. y Posada D. 2009. Molecular clock analysis. Chapter 11 in: P. Lemey, M. Salemi,
A.M. Vandamme (eds.). The Phylogenetic Handbook 2nd edition. Cambridge University
Press, Cambridge, UK, pp. 362-380.
Linnaeus, C. 1758. Tomus I. Syst. nat., ed. 10. Holmiae, Laurentii Salvii: [1-4], 1-824
Llorente-Bousquets, J y Morrone, J. J. 2001. Introducción a la biogeografía en Latinoamérica:
teorías, conceptos, métodos y aplicaciones. Editorial, UNAM, pp. 129-137.
Mardulyn, P. 2012. Trees and ⁄ or networks to display intraspecific DNA sequence varia-
tion?. Molecular Ecology (2012) 21, 3385–3390
Marshall, J.S., 2006. The geomorphology and physiographic provinces of Central America.
In: Bundschuh, J., Alvarado,G. (Eds.), Central America: Geology, Resources, and Hazards.
Taylor and Francis, pp. 75-122.
Martin J. 2007. New material of the late cretaceous globidontan Acynodon iberoccitanus
(Crocodylia) from Southern France. J Vert Paleontol 27:362–372.
Mccoid, M. J. y Kleberg C.. 1995. Nonnative reptiles and amphibians. Págs. 433- 437. En: E.
T. LaRoe, G. S. Farris, C. E. Puckett, P. D. Doran & M. J. Mac (eds.), Our living resources: a

68
report to the nation on the distribution, abundance, and health of U.S. plants, animals, and
ecosystems. U.S. Department of the Interior, National Biological Service, Washington, D.C
Medem, F. 1955. A new subspecies of Caiman sclerops. Fieldiana (Zoology), 37, 339–343.
Medem, Federico. 1958. Problemas faunísticos de Colombia. El conocimiento actual sobre
la distribución geográfica y ecológica de los Crocodylia en Colombia. Revista Universidad
nacional de Colombia, 23, 37-57.
Medem, Federico. 1962. La distribución geográfica de los Crocodylia y Testudinata en el
departamento del Chocó. Revista de la Academia Colombiana de Ciencias exactas físicas y
naturales. Vol. XI. Nº 44.
Medem, Federico. 1981. Los Crocodylia de Sur América I. Los Crocodylia de Colombia. COL-
CIENCIAS-Universidad Nacional de Colombia. Bogotá.
Medem, Federico. 1983. Los Crocodylia de Sur América II. COLCIENCIAS-Universidad Nacio-
nal de Colombia. Bogotá.
Morales-Betancourt, M. A. y Lasso, C. A. 2013. Amenaza a los Crocodylia en Colombia. Pp.
275-297. En: Morales-Betancourt, M. A., C. A. Lasso, J. De La Ossa V. y A. Fajardo-Patiño
(Editores). VIII. Biología y conservación de los Crocodylia de Colombia. Serie Editorial Recur-
sos Hidrobiológicos y Pesqueros Continentales de Colombia. Instituto de Investigación de
Recursos Biológicos Alexander von Humboldt (IAvH). Bogotá, D. C., Colombia.
Moritz C. 1994. Defining ‘Evolutionarily Significant Units’ for conservation. Trends in Ecology
& Evolution, 9, 373-375.
Nei, M. & Kumar, S. 2000. Molecular Evolution and Phylogenetics. Oxford University

69
Press, New York. 333 pp.
Nixon, K. C. 1999-2002. WinClada ver. 1.0000 Published by the author, Ithaca, NY, USA.
Nores, M., 2004. The implications of Tertiary and Quaternary sea level rise events for avian
distribution patterns in the lowlands of northern South America. Global. Ecol. Biogeogr, 13,
149-161.
Novick, R.R., Dick, C.W., Lemes, M.R., Navarro, C., Caccone, A., Bermingham, E. 2003. Ge-
netic structure of Mesoamerican populations of Big-leaf mahogany (Swietenia macrophylla)
inferred from microsatellite analysis. Mol. Ecol. 12, 2885-2893.
Oaks, J.R. 2011. A time-calibrated species tree of Crocodylia reveals a recent radiation of
the true crocodiles. Evolution, 65, 3285–3297.
Palomares, F., Godoy, J.A., Piriz, A, O’Brien, S.J., Johnson, W.E. 2002. Faecal genetic analysis
to determine the presence and distribution of elusive carnivores: design and feasibility for
the Iberian lynx. Molecular Ecology. 11: 2171-2182.
Parida S. K., RajKumar K. A., Dalal V., Singh N. K., Mohapatra T. 2006. Unigene derived mi-
crosatellite markers for the cereal genomes. Theor. Appl. Genet. 112 808–817.
10.1007/s00122-005-0182-1
Parkinson, C.L., Zamudio, K.R., Greene, H.W., 2000. Phylogeography of the pitviper clade
Agkistrodon: historical ecology, species status, and conservation of cantils. Mol. Ecol. 9,
411-420.

70
Parra-Olea, G., García-París, M., Wake, D.B., 2004. Molecular diversification of salamanders
of the tropical American genus Bolitoglossa (Caudata: Plethodontidae) and its evolutionary
and biogeographical implications. Biol. J. Linn. Soc. Lond 81, 325-346.
Posada D. y Crandall K.A. 1998. Modeltest: testing the model of DNA substitution. Bioinfor-
matics 14 (9): 817-818.
Rambaut, A., Suchard, M.A., Drummond, A.J., 2014. Tracer v1.6.
Ribas, C. C., Aleixo, A., Nogueira, A. C. R., Miyaki, C. Y. y Cracraft, J. 2012. A palaeobiogeo-
graphic model for biotic diversification within Amazonia over the past three million years.
Proc. R. Soc. B 279, 681–689 doi:10.1098/rspb.2011.1120
Rodríguez M, 2000. Cocodrilos (Archosauria: Crocodylia) de la Región Neotropical. Biota Co-
lombiana. 1(2)135-140.
Rohland, N., Siedel, H y Hofreiter, M., 2004. Nondestructive DNA extraction method for mi-
tochondrial DNA analyses of museum specimens. BioTechniques 36:814-821
Rohland, N., Hofreiter, M., 2007. Ancient DNA extraction from bones and teeth. Nature Pro-
tocols 2, 1756–1762
Ronquist F, Huelsenbeck JP. 2003. MrBayes 3: Bayesian phylogenetic inference under mixed
models. Bioinformatics 19:1572–1574.
Rueda-Almonacid et al., J; Carr, J; Mittermeier, R; Rodríguez-Mahecha, Mast, J; Vogt, R;
Rhodin, A; de la Ossa-Velásquez, J; Rueda-Almonacid et al., J y Mittermeier, C. 2007. Las
tortugas y los cocodrilianos de los países andinos del trópico. Serie de guías tropicales de

71
campo Nº 6. Conservación Internacional. Editorial Panamericana, Formas e Impresos. Bo-
gotá, Colombia. 538 pp.
Ryder, O. A. 1986. Species conservation and systematics - the dilemma of subspecies.
Trends in Ecology & Evolution 1: 9-10.
Salas-Gismondi, R.; Flynn, J. J.; Baby, P.; Tejada-Lara, J. V.; Wesselingh, F. P.; Antoine, P. -O.
2015. A Miocene hyperdiverse crocodylian community reveals peculiar trophic dynamics in
proto-Amazonian mega-wetlands. Proceedings of the Royal Society B: Biological Sci-
ences 282 (1804): 20142490.
Saldarriaga-Córdoba MM., Sasa M., Pardo R., Méndez MA. 2009. Phenotypic differences in
a cryptic predator: factors influencing morphological variation in the terciopelo Bothrops
asper (Garman, 1884; Serpentes: Viperidae). Toxicon. 2009 Dec 1;54 (7):923-37
Sambrock J., Fritsn E. y Maniatis T. 1989. Molecular cloning: A laboratory manual. 2nd edi-
cion. V1. Cold spring HarborLaboratory Press. New York.
Savage, J. M., 1982. The enigma of the Central American herpetofauna: dispersals or vicar-
iance. Ann. Mo. Bot. Gard. 69, 464- 547.
Savage, J.M., 1987. Systematics and distribution of the Mexican and Central American rain-
frogs of the Eleutherodactylus gollmeri group (Amphibia: Leptodactylidae). Fieldiana Zool-
ogy, 33, 1-57.
Schmidt, K.P. 1928. Notes on South American caimans. Field Museum of Natural History,
Publication 252, Zoological Series, 7, 205–231.

72
Smith, H.M. y Smith, R.B. 1977. Synopsis of the Herpetofauna of México. Vol. V. Guide to
Mexican Amphisbaenians and Crocodilians. Bibliographic Addendum II. John Johnson, North
Bennington, Vt. 187 pp.
Strimmer, K. y Haeseler Von A. 2009. Genetic distances and nucleotide substitution models:
Theory. Chapter 4 in: P. Lemey, M. Salemi, A.M. Vandamme (eds.). The Phylogenetic Hand-
book 2nd edition. Cambridge University Press, Cambridge, UK, pp. 111-125.
Solomon SE, Bacci M Jr, Martins J Jr, Vinha GG, Mueller, U.G. 2008. Paleodistributions and
Comparative Molecular Phylogeography of Leafcutter Ants (Atta spp.) Provide New Insight
into the Origins of Amazonian Diversity. PLoS ONE 3(7): e2738. doi:10.1371/jour-
nal.pone.0002738
Sumida, M., Agata, M., Kaneda, H. & Yonekawa, H. 1998. Evolutionary relationships Among
Japanese pond frogs inferred from mitochondrial DNA sequences of cytochrome b and 12S
ribosomal RNA genes. Genes Genet. Syst., 73: 121-133.
Swofford, DL. 2003. PAUP_: phylogenetic analysis using parsimony (and other methods),
version 4.0b 10. Sunderland, MA: Sinauer Associates.
Tamura, K, Stecher, G, Peterson, D, Filipski, A. y Kumar S. 2013. MEGA6: Molecular Evolu-
tionary Genetics Analysis version 6.0. Mol Biol Evol. 2013 Dec; 30(12):2725-9
Thompson, Julie, HIGGINS, Desmond, GIBSON, Toby. 1994. CLUSTAL W: improving the sen-
sitivity of progressive multiple sequence alignment through sequence weighting, positions-
specific gap penalties and weight matrix choice. En: Nucleic Acids Research. 22, (1994):
4673-4680.

73
Templeton, A. 2004. Statistical phylogeography: methods of evaluating and minimizing in-
ference errors. Molecular Ecology. 13: 789-809.
Templeton, A. 2008. Nested clade analysis: an extensively validated method for strong phi-
logeographic inference. Molecular Ecology. 17: 1877-1880.
Templeton, A.R. & Sting, C.F. 1993. A cladistic analysis of phenotypic associations with hap-
lotypes inferred from restriction endonuclease mapping. IV. Nested analyses with clado-
grams uncertainty and recombination. Genetics. 134: 659-669.
Templeton, A.R., Boerwinkle, E. & Sing, C.F. 1987. A cladistic analysis of phenotypic associ-
ation with haplotypes inferred from restriction endonuclease mapping. I. Basic theory and
an analysis of alcohol dehydrogenase activity in Drosophila. Genetics. 117: 343-351.
Thorbjarnarson, J. B. y G. Hernandez. 1988. Recent Investigations into the Status of Orinoco
Crocodiles in Venezuela. Proceedings of the 9th Working Meeting of the Crocodile Specialist
Group of the Species Survival Commission of IUCN- The World Conservation Union, vol 2.
Vanzolini, P.E., Heyer, W.R. 1985. The American herpetofauna and the interchange. In:
Stehli FG, Webb SD, editors. The great American biotic interchange. NewYork:
Plenum Press. p 475–487.
Vasconcelos W. R., Hrbek T. II, Da Silveira R, de Thoisy B, Marioni B, Farias IP. 2006. Popula-
tion genetic analysis of Caiman crocodilus (Linnaeus, 1758) from South America. Genet Mol
Biol 29:220–220.
Velasco, A. & Ayarzagüena, J. 2010. Spectacled Caiman Caiman crocodilus. In: Manolis, S.C.
& Stevenson, C. (Eds), Crocodiles. Status Survey and Conservation Action Plan. Third Edition,
Crocodile Specialist Group, Darwin, Australia, pp. 10–15.

74
Venegas-Anaya M., Crawford A., Escobedo A., Sanjur O., Densmore I. y Bermingham E. 2008.
Mitochondrial DNA Phylogeography of Caiman crocodilus in Mesoamerica and South Amer-
ica. Journal of Experimental Zoology 309ª.
Van der Hammen, T. 2000. Aspectos de historia y ecología de la biodiversidad norandina y
amazónica. Rev. Acad. Colomb. Cienc. 24(91): 231-245, 2000.
Villarreal H., M. Alvarez, S. Cordoba, F. Escobar, G. Fagua, F. Gast, H. Mendoza, M. Ospina y
A. M. Umaña. 2004. Manual de métodos para el desarrollo de inventarios de biodiversidad.
Programa de Inventarios de Biodiversidad. Instituto de Investigaciones de Recursos Biológi-
cos Alexander Von Humboldt. Bogotá, Colombia. 236 p.
Vogelstein B, Gillespie D. 1976. Preparative and analytical purification of DNA from agarose.
Proc Nat Acad Sci USA 76: 615-619
Vogler, A. P. y R. Desalle. 1994. Diagnosing units of conservation management. Conserva-
tion Biology 8: 354-363.
Wang, I. J., Crawford, A. J, Bermingham, E. 2008. Phylogeography of the Pygmy Rain Frog
(Pristimantis ridens) across the lowland wet forests of isthmian Central America. Molecular
Phylogenetics and Evolution 47: 992–1004
Webb, SD., Rancy, A., 1996. Late Cenozoic evolution of the Neotropical mammal fauna. In:
Jackson, J.B.C., Budd, A.F, Coates, A.C. (Eds.), Evolution and Environmental in Tropical Amer-
ica. University of Chicago Press, Chicago, pp.335-358.
Weigt, LA., Crawford, A.J., Rand, A.S., Ryan, M.J., 2005. Biogeography of the túngara frog,
Physalaemus pustulosus: a molecular perspective. Mol. Ecol. 14, 3857-3876.

75
Wiens, J. & Reeder, T. 1995. Combining data sets with different numbers of taxa for phylo-
genetic analysis. Syst. Biol., 44: 548-558
Zamudio, K.R., Greene, H.W., 1997. Phylogeography of the bushmaster (Lachesis muta: Vi-
peridae): implications for Neotropical biogeography, systematics, and conservation. Biol. J.
Linn. Soc. 62, 421-442.