boletÍn nº 2, junio de 2006seprot/docs/boletines/seprot... · empleado en responder correos y...
TRANSCRIPT

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
1
BOLETÍN Nº 2, Junio de 2006
La cuenta atrás para la celebración conjunta del 2º Congreso de la SEProt y del 1. Congreso de la European Proteomics Association (EuPA) está en marcha...

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
2
Sede social: Instituto de Biomedicina de Valencia, C.S.I.C. c/Jaime Roig 11, 46010 Valencia. Tel.: 96 339 1778; Fax: 96 369 0800 C.I.F. G97465629 Nº Registro Nacional de Asociaciones: 584180
http://www.cbm.uam.es/seprot
BOLETÍN DE
LA SOCIEDAD
ESPAÑOLA DE
PROTEÓMICA
(SEProt)©
nº 2, Junio de 2006
Editores: Jesús V. Jorrín, Jesús Vázquez y Juan J. Calvete
© SEProt. Los artículos y colaboraciones reflejan la opinión de sus autores y no necesariamente la opinión de la SEProt. Se autoriza la reproducción del contenido, siempre que se cite la procedencia.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
3
EDITORIAL El número 1 del Boletín vio la luz en agosto del pasado año. El número 2 llega pues a vuestras manos con un considerable retraso sobre la idea original de editar al menos dos números anuales. Varios han sido, a nuestro juicio, los factores que han contribuido a este desfase y, aunque por fuerza hay que enumerarlos en algún orden, éste pudiera ser alterado arbitrariamente. En primer lugar, los editores lejos de ser profesionales del periodismo, somos investigadores de un sistema que cada vez más despiadadamente nos obliga a burocratizarnos con convocatorias (proyectos de investigación, becas FPU, FPI, Comunidades Autónomas, I3P, Ramón y Cajal, Juan de la Cierva, Severo Ochoa, Planes Estratégicos, infraestructura varia, CIBER, Consolider, etc. etc,) cuyos plazos límites de solicitud son irrisorios. No hay tampoco que desdeñar el tiempo empleado en responder correos y faxes, escribir y corregir manuscritos y tesis doctorales, evaluar la actividad científica de colegas, redactar memorias, etc. Y algunos de nosotros hasta pretendemos mantenernos al día en nuestra disciplina e incluso desarrollar en el laboratorio líneas de investigación propias, evitando en lo posible el riesgo cardiovascular, o tener que recurrir a velocidades einstenianas para estirar el marco espacio-temporal. Ejercemos, pues, nuestro compromiso editorial con dedicación entusiasta pero amateur. Es por ello que aplaudimos la incorporación de Jesús Vázquez, actual responsable de la página web de la Sociedad, al equipo editorial del Boletín. Su curriculum proteómico y su espíritu imaginativo a la vez que crítico están en línea con la orientación más científica que testimonial que pretendemos dar a la revista.
Pero cualquier iniciativa, por voluntariosa que sea, está destinada a diluirse si no encuentra eco entre los destinatarios. Para que el Boletín llegue a ser el vehículo de comunicación de la SEProt, y no solo un reflejo de monólogos de su Junta Directiva, son necesarias las contribuciones de los socios. ¡Animaos! Recordad que ¡La SEProt somos tod@s! y participad activamente para darle contenido a la existencia de nuestra Sociedad y del Boletín. Por de pronto, este número del Boletín incorpora artículos firmados por miembros de la Junta Directiva sobre las últimas reuniones de la Junta Directiva, la organización del 2º Congreso de la SEProt, la edición del número especial de Proteomics, marcha de la EuPA, ProteoRed, HUPO, cursos impartidos, etc. Este 2º Boletín incluye también las primeras reseñas de Tesis Doctorales, específicamente las defendidas por E. Santamaría (del grupo de Fernando Corrales, Universidad de Navarra), M.A. Castillejo (del grupo de Jesús Jorrín, Universidad de Córdoba), de A.M. Rodríguez Piñeiro (Departamento de Bioquímica, Genética e Inmunología de la Universidad de Vigo, dirigida por los Dres. María Páez de la Cadena Tortosa y Francisco Javier Rodríguez Berrocal.) y de D. López (del grupo de Jesús Vázquez, CBM, Madrid). Este número del Boletín, cuya distribución por motivos presupuestarios y “de ahorro de árboles” se realiza de forma electrónica y podrá asimismo descargarse libremente desde la página web de la SEProt, está en parte dedicado a los Servicios/ Unidades de Proteómica. En ediciones sucesivas pretendemos que los diversos grupos de investigación que utilizan de una u otra manera la proteómica presenten sus líneas de

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
4
trabajo. El Boletín puede, y debe, ser asimismo un foro de discusión e intercambio de ideas reflejadas en artículos de opinión, especulación, ensayo, divulgación, etc. ¿Te animas? Nuestro objetivo a corto plazo es darle un contenido más científico a nuestro Boletín y quien sabe si quizás esta
gaceta llegue a transformarse con el tiempo en una revista científica. Pero, una vez más, todo dependerá esencialmente de las aportaciones que cada uno de nosotros estemos dispuestos a hacer. Ahí queda lanzado el reto...

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
5
INDICE EDITORIAL 3CONTRIBUCIONES DE LA JUNTA DIRECTIVA 7La columna del presidente. Juan J. Calvete 8Reuniones de la Junta Directiva de la SEProt. Jesús V. Jorrín Novo 10Presupuesto 2006. David Andreu 13Becas de Formación de la Sociedad Española de Proteómica. David Andreu.
14
La singladura de la Sociedad Española de Proteómica a través de su página web. Salvador Martínez-Bartolomé y Jesús Vázquez
16
Algunos avances, novedades y consideraciones en el proyecto HUPO. Fernando J. Corrales
18
INFORMACIÓN DE INTERÉS 21Publicación del Número Especial de Proteomics dedicado al “I Congreso de la SEProt”. Jesús V. Jorrín
22
II Spanish Proteomics Society (SEProt)/I European Proteomics Association (EuPA) Join Meeting. Juan J. Calvete
24
2º Premio SEProt. Juan José Calvete 26ARTÍCULOS DE OPINIÓN 28Sobre la identificación mediante espectrometría de masas de sitios de fosforilación y de nitrosilación y otras sutilezas postraduccionales. Jesús Vázquez
29
TESIS DOCTORALES 33Análisis de los mecanismos moleculares asociados al desarrollo de EHNA y HCC en un modelo de ratón deficiente en el gen MAT1A mediante proteómica funcional. Identificación de potenciales biomarcadores. Enrique Santamaría
34
La Proteómica aplicada al estudio de las respuestas de las plantas a estreses bióticos y abióticos. Mari Ángeles Castillejo Sánchez
36
Análisis del proteoma de suero humano para la identificación de proteínas con utilidad clínica en el cáncer colorrectal: Estudio proteómico de las isoformas de clusterina. Ana María Rodríguez Piñeiro
38
Desarrollo de nuevas metodologías para el análisis del proteoma. Daniel López-Ferrer
44
EL RINCÓN DEL SOCIO 46Ludoproteómica. Jesús Vázquez 47SERVICIOS DE PROTEOMICA 48Servicio de Proteómica Centro de Investigación Príncipe Felipe. Manuel M. Sánchez del Pino
49
Unidad de Proteómica del Servicio General de Investigación de Genómica y Proteómica de la Universidad del País Vasco. Kerman Aloria, Miren Josu Omaetxebarria, Asier Fullaondo, Jesus M. Arizmendi
52
Unidad de Proteómica. Fundación Centro Nacional de Investigaciones Cardiovasculares Carlos III. Juan Antonio López del Olmo, Emilio Camafeita, Enrique Calvo
54
Unidad de Proteómica de la Universidad Complutense de Madrid 60

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
6
Parque Científico de Madrid. Mª Luisa Hernáez y Concha Gil Servicio de Proteómica del CNIO. J. Ignacio Casal 65Servicio Científico-Técnico de Proteómica Universidad Pompeu Fabra/Parc de Recerca Biomèdica de Barcelona. David Andreu
68
Servicio de Proteómica del Centro de Biología Molecular “Severo Ochoa”. Anabel Marina Ramírez
73
Plataforma de Proteómica, Parc Cientific de Barcelona. Eliandre de Oliveira, David Bellido, Antonia Odena, Stéphanie Boussert
79
Unidad de Proteómica, Genómica y Bioinformática del Centro de Investigación Médica Aplicada. Fernando J. Corrales
83
Unidad de Servicios de Proteómica del CIC-Biogune. Nere Alkorta, Eva Rodriguez, Vladimir Montero, Stephanie Richer y Felix Elortza
86
Grupo de Proteómica de Alimentos Marinos. José Manuel Gallardo, Carmen Piñeiro, Mónica Carrera, Ignacio Ortea y Lorena Barros
90
Unidad de Proteómica de la Universidad de Córdoba. Samuel Ogueta, Consuelo Gómez, Raquel González, M. Ángeles Castillejo
94
INSTRUCCIONES A LOS AUTORES 98IMPRESOS DE INSCRIPCIÓN A LA SEProt 99DIRECTORIO DE SOCIOS 105SOCIOS PATROCINADORES 116

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
7
CONTRIBUCIONES DE LA
LA JUNTA DIRECTIVA

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
8
La Columna del Presidente "276, 275, 274..."
La cuenta atrás para la celebración de nuestro 2º Congreso está irremediablemente activada, y a estas alturas del año parece que los días tengan menos horas y los minutos menos segundos! Comienzan a querer materializarse los mismos fantasmas que acompañan a cualquier estreno escénico y que no se esfumarán hasta el mismísimo día de la inauguración, cuando se alce el telón y todos los actos se sucedan en el orden previsto. Y es que este 2º Congreso de la SEProt cuenta con
un programa complicado y que difícilmente va a ser del agrado de todos. A la idea original de darle una orientación iberoamericana al 2º Congreso de la SEProt se ha sumado la oportunidad de contribuir al desarrollo de la European Proteomics Association organizando su primer congreso conjuntamente con el nuestro. Aunque ha habido que cambiar el idioma oficial al Inglés, creo, no obstante, que el esfuerzo vale la pena y que hemos conseguido elaborar un programa sólido y equilibrado para todas las partes.
Además de las conferencias invitadas, están previstas 3 sesiones de exposiciones orales cortas (formato 10 + 5 min) elegidas entre las comunicaciones
libres, especialmente de jóvenes proteómicos nacionales. Los detalles del congreso se actualizarán en el portal http://Proteomics-Valencia2007.ibv.csic.es.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
9
Cualquier sugerencia para mejorar el contenido y desarrollo del evento será muy bienvenida!
Tengo también el placer de anunciar que Bruker Biosciences Española S.A. continuará patrocinando el Premio SEProt, que en esta edición se repartirá entre un trabajo publicado entre 2005-2006 y un póster presentado al congreso (ver bases de la convocatoria en este Boletín). Quiero, en particular, agradecer públicamente esta iniciativa a su promotor Alberto Sánchez, a la vez que animo a otros líderes del sector proteómico a promover otras acciones similares.
La celebración de nuestro 2º Congreso coincidirá con el ecuador del periodo de gestión de su Junta Directiva. El Artículo 6 de los Estatutos de la SEProt establece que "La Junta se renovará por mitades cada dos años, alternando el cese de Presidente, Tesorero y la mitad de los vocales con los de Vicepresidente, Secretario y la
otra mitad de los vocales. Los candidatos podrán ser propuestos por cualquier Socio Ordinario siempre que su petición sea apoyada por la firma de otros cuatro Socios Ordinarios y se reciba en la Secretaría, al menos treinta días antes de la votación correspondiente." Animo, pues, desde ya mismo a todos aquellos socios con voluntad decidida y desinteresada de contribuir al desarrollo de nuestra Sociedad a elaborar una candidatura audaz que aporte nuevas ideas. Durante el bienio siguiente a su elección por la Asamblea General, los nuevos cargos podrán asistir (con voz aunque sin voto) a las asambleas de la Junta Directiva para familiarizarse con el funcionamiento actual del órgano de gestión de la SEProt. Vaya por delante el apoyo de todos los miembros de la presente Junta Directiva. Yo no hice la mili, pero intuyo que solo resta que los voluntarios den un paso al frente...

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
10
Reuniones de la Junta Directiva de la SEProt
Jesús V. Jorrín Novo
En el Capítulo III, Artículo 17 de los Estatutos de la SEProt, se establece que la Asamblea General es el órgano supremo de gobierno y que está constituida por todos los asociados, reuniéndose al menos (es lo normal) una vez cada dos años, durante los congresos. No obstante, es necesario asegurar el funcionamiento diario de la sociedad, responsabilidad que recae en el Presidente y la Junta Directiva como órgano de representación (Capítulo II, artículos 6-16). Es objetivo de esta Directiva contar a los asociados ese día a día y para ello utilizamos como vehículo de comunicación los correos electrónicos o la página web. Además,
pretendemos haceros llegar, utilizando el boletín, el orden del día, contenido y conclusiones de nuestras reuniones, por lo que inauguramos este serial: “Reuniones de la Junta Directiva de la SEProt”.
Desde la publicación de nuestro anterior boletín, hemos celebrado dos reuniones, el 10 de octubre de 2005, en Valencia, y el 23 de febrero de 2006, en Barcelona.
Los aspectos más relevantes de estas reuniones han sido:
1. Valencia, 10 de octubre de 2005

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
11
i) La puesta en marcha del II
Congreso de la SEProt (http://Proteomics-Valencia2007.ibv.csic.es).
ii) La propuesta, aprobada por la mayoría de los socios, de nombramiento de J. Fenn como socio de honor.
iii) La participación activa de nuestra sociedad en el Proyecto EuPA. Concha Gil es miembro del Comité Ejecutivo, y junto con Juan Pablo Albar, responsable del Comité de Educación. Juanjo Calvete es miembro del Comité Económico,
Fernando Corrales del de HUPO y Jesús Jorrín del de Comunicaciones y Congresos. Todos ellos fueron elegidos por votación de entre los representantes de las distintas sociedades.
iv) Fernando Corrales ha sido nombrado como co-coordinador del grupo de genómica y proteómica de la SEBBM.
v) Por motivos personales, José M. Gallardo dimite como vocal y se incorpora a la directiva Ángela Moreno.
2. Barcelona, 23 de febrero de 2006
i) Se aprueba, por unanimidad, tanto el resumen económico 2005 como el presupuesto para el 2006. Existe dificultad en el cobro de algunas cuotas. En este punto se acuerda, como procedimiento, enviar el mes de octubre una carta a los socios indicándoles que se va a pasar al cobro la cuota del año siguiente y que las bajas se comuniquen por escrito.
ii) Acciones para mejorar las relaciones con los socios de la SEProt: cursos, becas, etc. Este punto es una de las mayores preocupaciones de la Junta Directiva. Se acuerda: a) conceder ayudas y becas para viajes de los socios. A ello se le ha asignado una partida presupuestaria, para el año 2006, de 4500 Euros. Las ayudas tendrán un tope de 1000 Euros y se

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
12
abrirán dos convocatorias abiertas que finalizarán el 30 de julio y 30 noviembre. La intención es ayudar a cubrir los gastos de socios en laboratorios de proteómica, tanto extranjeros como nacionales; b) la edición, en castellano, de un libro/manual/monografía de proteómica; c) la impartición de
cursos teórico-prácticos de proteómica.
iii) El II Congreso de la SEProt se ha convertido también en el I Congreso EuPA.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
13
Presupuesto 2006 David Andreu
INGRESOS (En Euros) Cuotas patrocinadores (Agilent, Applera, Beckman Coulter, BioGen, BioRad, Bruker, GE Healthcare, Sigma-Aldrich, Thermo, Waters)
6000.00
Cuotas socios 3500.00Total ingresos 9500.00
GASTOS (E Euros) Becas SEProt* 4000.00Administración (correo, oficina, …) 500.00Reuniones Junta Directiva 2000.00Honorarios webmaster 600.00Auxiliares secretaría y tesorería 1200.00Cuota COSCE 194.00Cuota EuPA 300.00Otros gastos 50.00Total gastos 8844.00Superávit previsto (a fondos de reserva) 656.00*La cantidad presupuestada (40% ingresos previstos) es modesta pero considerable para las posibilidades económicas actuales de la Sociedad. La Sociedad mantendrá en lo posible, y gracias a la colaboración de los socios patrocinadores, el compromiso de dedicar una parte importante de sus recursos mediante becas de formación

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
14
Becas de Formación de la Sociedad Española de
Proteómica D. Andreu
En la declaración fundacional de la Sociedad Española de Proteómica (SEProt) figuran objetivos y actividades tales como “impulsar la formación científica y técnica de sus miembros” (objetivo 3c), “propiciar la participación de los asociados en actividades” (objetivo 3d), y “fomentar la asistencia de estudiantes de doctorado a los eventos científicos auspiciados por la sociedad mediante la concesión de becas” (actividad 4e).
En consecuencia con dichos propósitos, y en la medida de sus posibilidades económicas, la SEProt convoca becas para contribuir a la formación de jóvenes investigadores en proteómica. La concesión de dichas becas se regirá por las siguientes normas: 1. Las actividades susceptibles de financiación mediante becas SEProt tendrán como objetivo fundamental la formación del solicitante en áreas y tecnologías relacionadas con la proteómica. Como actividades preferentes, sin descartar otras, se contemplan la participación en cursos o workshops de especialización, y la realización de estancias formativas en laboratorios debidamente acreditados. En la actual convocatoria de 2006 se concederán becas para actividades que tengan lugar, preferentemente, antes del 30 de septiembre de 2007.
2. Los/as solicitantes deberán cumplir los siguientes requisitos:
Ser miembro de la SEProt, y hallarse al corriente de la cuota
Nacionalidad española o vinculación estable con un laboratorio de investigación radicado en España.
Fecha de nacimiento posterior al 31 de Diciembre de 1976.
3. Además de acreditar lo anterior, la solicitud deberá incluir la siguiente documentación:
Carta de presentación donde se indique claramente la actividad para la que se solicita la beca y el interés de ésta para la formación del/de la solicitante.
DNI o pasaporte (digitalizado) Curriculum vitae Breve memoria de la actividad a
realizar, incluyendo plan de trabajo y estimación desglosada (viajes, manutención) de gastos.
Documento de aceptación por parte del laboratorio/institución donde vaya a llevarse a cabo la actividad prevista
Carta de un/a supervisor/a científico/a, recomendando la realización de la actividad para la que se solicita la beca.
4. La Junta Directiva de la SEProt designará para cada convocatoria de becas un comité de selección compuesto por tres miembros, más un suplente. En la presente convocatoria de 2006, el

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
15
comité estará compuesto por Juan Pablo Albar, David Andreu (tesorero SEProt) y Fernando Corrales, con Jesús Vázquez como suplente. 5. Las solicitudes, totalmente en formato electrónico (word o pdf), deberán enviarse a la secretaría de la SEProt, [email protected], antes del 30 de Septiembre de 2006. 6. En su resolución, el comité propondrá a la Junta Directiva de la SEProt la concesión de un número discrecional de becas, de un máximo de 1000€ (mil euros) cada una y hasta un total de 4000€ (cuatro mil euros) por anualidad. Dentro de este marco, el comité de selección estará facultado para decidir el número y cuantía de las becas a otorgar, en función de la calidad y el coste económico de las diversas solicitudes.
7. Las resoluciones del comité de selección se publicarán en la página web de la SEProt antes del 15 de Octubre de 2006 y no podrán ser recurridas. El comité no mantendrá correspondencia sobre las mismas ni está obligado a devolver a los solicitantes la documentación facilitada con la solicitud. 8. Una vez realizada la actividad, y en el plazo máximo de un mes, deberá remitirse un informe a la Secretaría de la SEProt, [email protected]. 9. Aunque la presente convocatoria no supone obligación alguna de reedición, siempre que sus recursos lo permitan la SEProt seguirá promoviendo iniciativas similares, en cumplimiento de sus fines y destinadas a impulsar la formación de sus miembros.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
16
EuPA …. Concha Gil
La singladura de la Sociedad Española de Proteómica a través de su página web
Salvador Martínez Bartolomé y Jesús Vázquez
Aunque la SEProt se inauguró
oficialmente en abril del 2004, su andadura viene de cuando nos reunimos en Córdoba en febrero de 2003, de forma oficiosa, todos, o casi todos, los proteómicos del país. Desde entonces en la sociedad ha habido una actividad casi frenética, y rara ha sido la semana en la que no hayamos tenido que insertar un anuncio, un nuevo evento, una nota informativa. La actividad de la web refleja la actividad de la SEProt y resume la singladura de la sociedad desde sus comienzos.
Por la web hemos anunciado un
total de nueve cursos teóricos y prácticos y cinco seminarios científico-técnicos, todos ellos por supuesto relacionados con la Proteómica; hemos publicado once notas informativas de interés, y trece anuncios de otros tantos congresos han puesto su link al acceso del socio. La web también ha anunciado siete ofertas proteómicas de trabajo, en laboratorios de investigación y en unidades asociadas y servicios centrales, lo que habla por sí solo del creciente interés que están despertando estas tecnologías. El logo de la SEProt ha aparecido en casi todos estos eventos y con tanto tejemaneje va haciéndose popular entre los científicos españoles y no tan españoles.
Aunque los seminarios
científicos no han puesto el listón
precisamente bajo, llaman la atención los numerosos cursos impartidos, repartidos por toda la geografía nacional. De carácter teórico-práctico la mayoría de ellos, y a veces bioinformático, estos eventos reflejan el ímpetu y las ganas de muchos de los miembros de la sociedad por comunicar y formar, y transmitir el potencial de las nuevas tecnologías. Hay tal vez una cierta falta de coordinación y de uniformidad de conocimientos y contenidos, pero ello nos parece secundario en esta etapa de crecimiento y asentamiento de la sociedad, en la que podríamos decir que debería primar el bullicio y la publicidad sobre la pureza y homogeneidad de la información. Ya llegará el momento de establecer un sello de calidad SEProt y también de preparar entre todos un texto de referencia, a modo de pilar básico sobre el que seguir construyendo.
La web de la SEProt también ha
informado puntualmente de los movimientos de la EuPA, nuestra European Proteomics Association, y nunca mejor usado el posesivo, pues esta supersociedad se fraguó, a fin de cuentas, bajo auspicios cordobeses, durante nuestro primer congreso. Y un gran número de miembros de los comités euperos, son de nuestro país. Pero esta historia no nos corresponde narrarla, y se cuenta, y con mayor

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
17
propiedad, en otra sección de este boletín.
Un repaso a las estadísticas revela la buena aceptación y el alto uso que se hace de nuestro portal. Desde el 6 de septiembre del 2004, la web ha experimentado más de 11.000 accesos, con una media actual superior a 20 conexiones diarias. El récord de actividad se batió el 20 de septiembre del 2004, tal vez coincidiendo con la proximidad del primer congreso de la sociedad. Pero más curioso resulta encontrar que sólo el 75% de las visitas están realizadas desde territorio español, procediendo cerca del 10% del Reino Unido, Dinamarca, México, Alemania y Estados Unidos, en este orden (Figura 1). Les siguen de cerca Suiza, Francia, Argentina y Colombia. Datos que parecen reflejar la composición de los miembros principales de la EuPA, y el interés que nuestra iniciativa está despertando en los países americanos, estimulados probablemente por el carácter trasatlántico que tendrá el segundo congreso de nuestra sociedad.
Finalmente, queremos comentar que la canalización de actividades estrictamente científicas a través de la web y del foro asociado (el Foro de Proteómica constituido a través de RedIris, en la dirección www.rediris.es/list/info/proteomica.es.html), es todavía muy limitada. El foro ha sido muy poco utilizado, y aún garantizando que cualquier mensaje llegará, de forma segura, a todos los inscritos, la preferencia por el anuncio directo en la web es evidente, y no se ha constituido todavía un verdadero debate de temas científicos, técnicos o metodológicos. Lo que no deja de llamar la atención en una sociedad en la que proliferan los blogs y los círculos de chatting. En fin, los medios ya están ahí, a disposición del socio, y sólo es cuestión de usarlos y aprovechar su utilidad. Desde estas páginas os animamos a contribuir activamente en la web, el foro o el boletín, haciéndolos más vivos y en definitiva, más acordes con estos tiempos tan proteómicos que corren.
Figura 1.- Clasificación del número de accesos
a nuestro portal web según el país de procedencia

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
18
Algunos avances, novedades y consideraciones en el
Proyecto HUPO Fernando J. Corrales
En una reciente editorial publicada en la revista Nature a finales del año pasado se resalta uno de los problemas a los que nos enfrentamos en el área de la proteómica y que, de alguna manera, la HUPO (http://www.hupo.org) está contribuyendo a resolver de manera decisiva. Por una parte está claro que la disponibilidad de técnicas que permiten la identificación de proteínas de forma masiva ha generado una gran cantidad de datos que, en muchos casos, ha demostrado ser difícilmente reproducible. Además, las diferentes técnicas generan diversos tipos de información, lo que dificulta enormemente la comparación de los resultados obtenidos en distintos laboratorios. Finalmente, parece fuera de discusión que el sólo establecimiento de catálogos de largas listas de proteínas no va a suponer un gran avance en el conocimiento de nuestra biología. Teniendo en cuenta estas consideraciones, la HUPO ha decidido poner un poco de orden al caos generado por el trabajo masivo realizado en estos últimos años sin métodos consensuados de estandarización, normalización y filtrado. Entre las 7 iniciativas que auspicia la HUPO, creo que en el momento actual la PSI (Proteomics Standard Initiative. http://psidev.sourceforge.net) es clave y no sólo condiciona el avance de los demás proyectos del consorcio, si no que, en mi opinión, tendrá
necesariamente una repercusión general. El foro de discusión no queda restringido a la HUPO si no que se ha implicado a la comunidad internacional para asegurar la consecución de acuerdos de estandarización que, incluso en esta etapa inicial, están ya siendo considerados muy favorablemente por las revistas más relevantes en el área. Además de la iniciativa MIAPE (Minimum Information on A Proteomic Experiment) como criterios mínimos que deben cumplir los experimentos realizados, cabe resaltar el desarrollo de dos recursos bioinformáticos como PRIDE (http://www.ebi.ac.uk/pride) y la creación del consorcio IMEX (Internacional Molecular Exchange. http://imex.sourceforge.net) cuyo objetivo es compartir la información de las 5 bases de datos de interacción de proteínas más importantes y generar información en los formatos estandarizados en la HUPO. Los días 4 y 5 de mayo se celebró en Bilbao la reunión del grupo de HUPO cuyo objetivo es el estudio del proteoma hepático, HLPP. Asistieron representantes de todos los grupos integrantes de esta iniciativa, miembros de ProteoRed y SEProt y participaron los líderes de la PSI. La reunión fue sumamente interesante ya que además de tratar los temas centrales en el desarrollo del proyecto HLPP, el contenido de las discusiones reflejó claramente las necesidades de la proteómica a día de hoy que,

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
19
probablemente, sean un anticipo de las objetivos prioritarios a abordar en el futuro próximo. El grupo liderado por el Dr. Fuchu He demostró su capacidad de análisis, disponen de espectrómetros de masas de última generación que permiten identificar 1.000 proteínas al día. En base a esta capacidad, se han conseguido estudiar unos 800.000 péptidos que permiten identificar unas 12.000 proteínas hepáticas con un único péptido y unas 6.000 con más de dos. Por su parte, en el grupo de la Dra. Laura Beretta se consiguieron identificar unas 4.400 proteínas analizando en una LTQ las 191 fracciones obtenidas a partir de un tripsinizado total de hígado humano, mediante 2D-LC. Estos datos están siendo analizados con diversas herramientas bioinformáticas con el objetivo de conseguir una interpretación biológica. En este sentido, al cruzar el repertorio de proteínas identificado por el grupo de la Dra. Beretta con el definido en el proyecto enfocado en el estudio del plasma, HPPP, (Nat. Biotech, 2006) únicamente se detectaron 379 proteínas en común. Sin embargo, a pesar del ingente trabajo realizado, quedó patente la necesidad de uniformizar formatos que permitan el almacenaje, intercambio, validación e interpretación de la información, aspectos en los que se está colaborando de manera activa con la PSI. Por otro lado, es evidente que la identificación de unos pocos miles de proteínas no refleja ni la dimensión ni el dinamismo del proteoma, algo que en mi opinión es necesario tener en cuenta para evaluar el impacto de las alteraciones implicadas en el desarrollo de enfermedades. Además de que los desarrollos tecnológicos futuros nos permitan aumentar nuestro campo de análisis, ya se están realizando estudios cuyo fin es la caracterización de subproteomas. Sin embargo, se presentaron datos que indican que el la congelación de las
muestras biológicas tiene un efecto negativo en el rendimiento del fraccionamiento subcelular y posterior análisis proteómico. En el caso de las muestras humanas, que habitualmente se congelan inmediatamente después de la extracción, la necesidad de procesar el tejido fresco podría plantear un problema logístico. La recogida y almacenamiento de muestras son fundamentales ya que condicionan el resultado de los análisis posteriores y por lo tanto deben realizarse de acuerdo a protocolos normalizados. Por otra parte, más allá de la necesidad de combinar métodos que permitan resolver mezclas de proteínas y espectrometría de masas para lograr su identificación, parece claro que no hay un único flujo de trabajo que asegure un resultado definitivo, quizás porque los resultados que se derivan de una u otra estrategia son, al menos en parte, complementarios. En cualquier caso, se está trabajando intensamente en la elaboración de normas mínimas cuyo objetivo es la estandarización los protocolos de forma que, una vez elegido el procedimiento analítico a usar, los resultados sean reproducibles. Independientemente de los métodos analíticos utilizados, se planteó la acuciante necesidad de establecer métodos que permitan asignar un alto grado de fiabilidad a los datos derivados de los experimentos, con especial énfasis en aquellos que implican la identificación masiva de proteínas, porque será difícil progresar sobre una base que genera incertidumbre. Estas cuestiones junto con el diseño de los recursos bioinformáticos mencionados anteriormente fueron algunos de los temas de discusión más interesantes. En cuanto al futuro, la obtención de anticuerpos, el desarrollo de métodos cuantitativos que permitan detectar proteínas diferenciales con potencial utilidad biomédica, el análisis de modificaciones postraduccionales o las

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
20
interacciones entre proteínas, fueron los temas más destacados, en alguno de los
cuales ya se está trabajando.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
21
INFORMACIÓN DE
INTERÉS

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
22
Publicación del Número Especial de Proteomics
dedicado al “I Congreso de la Seprot”
Jesús V. Jorrín Novo
1Acaba de salir (18 de abril de 2006) “on line” el número especial de la revista Proteomics dedicado al “I Congreso de la SEprot” y aún sin recibir las copias impresas que enviaremos a los autores y a los que participaron en el Congreso, me dispongo a escribir este pequeño artículo en el que intento, más que valorar, contar lo que ha sido el proceso de negociación con Wiley a través del Dr. Krauss (“Senior Publishing Editor”) y, en menor medida, del Dr. M. Dunn (“Editor in Chief”) y la historia de estos diez últimos meses. Intentando resumir
1 El presente artículo fue escrito el 20 de abril. Más que corregirlo y actualizarlo, he preferido dejarlo en su versión original. Ya hemos recibido el número impreso en papel y será enviado a los autores y socios de la SEPprot.
todo en una frase, diría que: “bien está lo que bien acaba”, y que si, como creo, los contenidos reflejan el nivel científico de la proteómica en España, éste es, en términos generales, bueno, a la altura de la de otros países y sociedades con más tradición y, probablemente, medios. Creo que, e independientemente de la motivación e interés particular de cada grupo, éste era el objetivo prioritario, aunque habrá análisis que difieran del aquí expuesto.
Siempre creí que la publicación
en Proteomics era el colofón perfecto al Congreso, aunque su ejecución no ha estado exenta de un buen número de dificultades, la más importante relacionada con el cambio de política en Wiley respecto a la publicación de proceedings de Congresos y el hecho de que sacando 18-24 números regulares al año no necesitan de números especiales. Finalmente pienso que mi cabezonería, insistencia y vehemencia junto con el apoyo personal de M. Dunn y A. Krauss hicieron posible el seguir adelante, aunque en los múltiples correos y llamadas telefónicas siempre noté dudas o reticencias al respecto, o, al menos, no la respuesta entusiasta que me hubiese gustado.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
23
Nada hubiese sido posible sin el
envío de manuscritos, y el temor inicial fue despareciendo a medida que los íbamos recibiendo. En total, y desde mayo (si no recuerdo mal) y finales de julio (cuando acabó el proceso de la primera revisión) recibimos un total de 40 manuscritos, de los que finalmente fueron aceptados 31. Todos pasaron el proceso de revisión estándar de Proteomics y que nadie dude de que hubiesen sido publicados, de forma similar, en cualquier número regular. Los que han actuado de “referees” y los editores invitados, así lo pueden atestiguar. Alguno de ellos, y es una opinión personal, fueron rechazados injustamente, pero los evaluadores son los que deciden y su opinión, que no tiene por qué ser infalible, ha prevalecido. A otros les ha costado más recibir la aceptación definitiva y han requerido de hasta cuatro rondas de revisión, lo que, de alguna manera ha retrasado la salida del número. El envío de versiones revisadas acabó a finales de octubre (desde Proteomics se dio un periodo de tres meses) y su segunda revisión acabo a finales de noviembre. Algunos artículos necesitaron el envío de una tercera versión y un número mínimo de una cuarta. Esto nos llevó hasta finales de año y al incumplimiento de una de mis expectativas, el de la publicación en el año 2005, ¡iluso de mi¡, … primera gran disculpa.
El tema económico ha sido uno
de los motivos por los que hubiese desistido del intento, aunque
afortunadamente mi desánimo e impotencia apareció cuando el proceso era irreversible. Siempre he pensado que el problema económico es el menos importante y pudieron más las ganas de ver el número especial que las complicaciones legales, la burocracia,… El tema está aún sin resolver. Total que los costes de publicación, para cada uno de vosotros, ha sido similar el de un “regular paper”. En un momento pensé que podría abaratar los costes a través de subvenciones que no se consiguieron y la inclusión de publicidad, que no hubo. Segunda gran disculpa.
He sido la imagen, como “guest
editor”, y responsable de la organización del congreso de este número, aunque quiero resaltar que ha sido sólo una imagen. Es de justicia mencionar aquí y reconocer, con letras mayúsculas, el trabajo realizado tanto en el evento como en la edición de Juanjo, Toño y Ángela, como queda recogido en la editorial, en la que se puede reconocer, claramente, el estilo del primero.
El resto lo podéis deducir si
acudís a la página web de Proteomics e inspeccionáis el contenido, leéis la editorial y cada uno de los artículos. De nuevo resumiendo, ..a pesar de los errores y problemas, volvería a hacer lo mismo, ya que aún se me escapa una alternativa mejor. Del dinero no hablamos y si acabo en la cárcel, espero vengáis a verme.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
24
II Spanish Proteomics Society (SEProt)/I European Proteomics
Association (EuPA) Join Meeting.
The Spanish Proteomics Society (SEProt) and the European Proteomics Association (EuPA) will conjointly celebrate their II and I Congress, respectively, in Valencia, Spain, February 10-14, 2007.
The venue of the meeting will be the Auditorio Santiago Grisolía of the Museo de Ciencias "Príncipe Felipe" (http://www.fundacioncac.es/eng/fundacion/catedra/catedra.jsp), one of the most impressive buildings of the Arts and Science complex designed by Santiago Calatrava.
http://www.cbm.uam.es/seprot
Proteomics & Pathology
-from both sides of the Atlantic Ocean-

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
25
INVITED SPEAKERS John B. Fenn (Nobel Laureate 2002) Rolf Apweiler (UK) The challenge to move Proteomics: beyond parts list construction Carlos Bloch Jr. (Br) Secretome imaging of amphibian skin peptide Carlos Carmona (UY) SELDI-TOF analysis of early biomarkers of Fasciola hepatica infection in the ovineLila Castellanos-Serra (Cu) Looking deeper into the proteome: subcellular fractionation followed by reversed-phase protein chromatography at basic pH María de los Ángeles Chávez (Cu) Carlos Cerveñansky/Rosario Durán (UY) Ser/Thr protein kinases in mycobacteria: autophosphorylation and substrate identification by proteomics approaches Alberto B. Domont (Br) Proteomics in sepsis Jörg Durner (Ger) S-nitrosylation and its function in Arabidopsis thaliana Emilio Gelpí (E) From big analogical machines to little digital “black boxes”: thirty two years of progress in mass spectrometry and its role in Proteomics Denis Hochstrasser (CH) Clinical proteomics and biomarkers discovery Peter James (S) Using high-throughput, low coverage 2D-PAGE for cancer biomarkers to direct low-throughput, high coverage 2D-HPLC for cancer biology György Marko-Varga (S) Discovery of peptides and proteins within disease-surveillance strategies Silvia Moreno (Ar) A proteomic approach to protein kinase A signaling in yeast Gabriel Padrón (Cu) Novel methods for the selective isolation of peptides using LC-MS Deborah Penque (P) Proteomics in biomarker discovery of Cystic Fibrosis, a monogenic disease Jasna Peter-Katalinic (Ger) Fernando Revert (E) GPBP overexpression produces aberrant conformers of the Goodpasture antigen Pier Giorgio Righetti (I) Agatha Christie and the proteome: a detective storyCarlos Robello (UY) Proteome Analysis of the causative agent of Chagas disease: Trypanosoma cruzi Manuel J. Rodríguez-Ortega (E) “Surfome” analysis of group A Streptococcus: a new approach to characterize surface proteins and to identify vaccine candidates Peter Roepstorff (DK) Glycoproteomics, is it relevant in disease studies? Jean Rosenbaum (F) Proteomics for the discovery of new markers and mediators of liver carcinogenesis Michael Przybylski (Ger) Molecular approaches for immunotherapy and diagnosis of neurodegenerative diseases based on affinity-proteomics and high-resolution mass spectrometry Josep Villanueva (E) Tumor-specific serum peptidome patterns
Looking forward to meeting you in Valencia…!!
OTHER EVENTS - Poster Sessions - Selected short oral communications - 2. SEProt Poster Award - 1. SEProt Annual Best Paper Award - State-of-the art-exhibits and workshops - Opening reception and banquet - … and much more…
A Website is operative at http://Proteomics-Valencia2007.ibv.csic.es Inquiries should be addressed to: the Congress Secretariat ([email protected]) or to the Chair of the Organizing Committee ([email protected])

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
26
CONVOCATORIA DEL SEGUNDO PREMIO DE LA SOCIEDAD ESPAÑOLA DE
PROTEÓMICA
Valencia, Mayo del 2006
La Sociedad Española de Proteómica (SEProt) convoca la segunda edición del PREMIO SOCIEDAD ESPAÑOLA DE PROTEÓMICA destinado a reconocer la labor en el campo de la Proteómica de un científico que desarrolle su actividad en España. El premio, patrocinado por Bruker BioSciences Española S.A. (http://www.bruker.es), está dotado con 2000 € y una placa de cerámica conmemorativa, y será entregado por un representante de Bruker Española durante el 2. Congreso de la SEProt que se celebrará en el Auditorio "Santiago Grisolía" del Museo de Ciencias "Príncipe Felipe" de la Ciudad de las Artes y las Ciencias de Valencia conjuntamente con el 1. Congreso de la European Proteomics Association (EuPA) entre los días 10-14 de Febrero del 2005 (http://Proteomics-Valencia2007.ibv.csic.es). En la presenta convocatoria se otorgarán dos galardones. Una mitad del premio (1000 €, diploma acreditativo y la placa de cerámica conmemorativa) será para una publicación científica relacionada con cualquier desarrollo o aplicación de
la Proteómica. La otra mitad del premio (1000 €, diploma acreditativo) será para una contribución en forma de panel al 2º Congreso de la SEProt. Las decisiones de los jurados serán inapelables. Los Premios no podrán concederse al mismo científico dos veces y podrían quedar desierto si así lo decidiese el jurado.
La elección de la publicación científica merecedora del galardón será realizada por un jurado de expertos designado a tal efecto por el Presidente de la Sociedad Española de Proteómica. Solo podrán optar al Premio científicos españoles sean o no socios de la SEProt. La labor investigadora deberá haber sido realizada en España y haber sido publicada entre Enero del 2005 y Diciembre del 2006. Los candidatos deberán remitir tres copias del trabajo al Secretario de la SEProt (Jesús V. Jorrín, Dpto. Bioquímica y Biología Molecular, Edificio Severo Ochoa (C6), Campus de Rabanales, Universidad de Córdoba, 14014 Córdoba). Deberá, asimismo,
2º Premio SEProt

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
27
adjuntarse los datos personales y profesionales del candidato, y un breve resumen de las razones que, a juicio del candidato, debieran ser consideradas por el jurado. La fecha límite para la recepción de los trabajos será el 1 de Diciembre del 2006. La decisión se tomará antes del 15 de Diciembre del 2006 y será dada a conocer en el programa final del congreso y a través del portal electrónico de la SEProt. El galardonado será invitado a participar en el 2. Congreso de la SEProt exento del pago de la tasas de inscripción, y se comprometerá a pronunciar una conferencia de 10-15 minutos basada en el trabajo premiado en la sesión de entrega del premio.
La elección del ganador(a) del Premio SEProt a la contribución en formato panel presentadas al 2º Congreso de la SEProt se efectuará entre los primeros firmantes del estudio por un jurado designado a tal efecto por el Presidente de la Sociedad Española de Proteómica. Solo podrán optar al Premio científicos españoles sean o no socios de la SEProt. La labor investigadora considerada deberá haber sido realizada en España y ser inédita o, en todo caso, no haber sido publicada con anterioridad a Diciembre del 2006.
Juan J. Calvete
(Presidente de la SEProt)

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
28
ARTÍCULOS DE
OPINIÓN

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
29
Sobre la Identificación mediante Espectrometría de Masas de Sitios de Fosforilación y de Nitrosilación y otras Sutilezas Postraduccionales
Jesús Vázquez1
No pretendo andarme con
revisiones, que muchas y buenas las hay, y de autores de más renombre. Intento más bien adelantar algunos comentarios, a modo de reflexiones en un asunto espinoso como éste, inaugurando así la sección de opinión de nuestro boletín.
La relevancia fisiológica de las modificaciones postraduccionales ha sido comentada hasta la saciedad. Y llevada al término extremo da lugar a una interesante teoría, según la cual la diferencia entre el hombre y el gusano, especies tan parejas en cuanto al número de genes, se debe a la extensión y complejidad de las modificaciones que experimentan sus proteínas. Se trata, en verdad, de un argumento idóneo para justificar nuestros proyectos en los farragosos formularios de las convocatorias públicas. Sin embargo, no es la única teoría, habiéndose esgrimido con idéntico propósito la complejidad del interactoma, y, muy recientemente, del transcriptoma. Claro que si añadimos el proteoma, que a secas hemos de entender en la perspectiva de los cambios dinámicos, y el metaboloma, y obviamos la flagrante simplificación, tendríamos la teoría completa. Las proteínas interaccionan y
se modulan y modifican entre sí y también se unen a los promotores de los genes, que a su vez activan la producción de otras proteínas, las cuales modulan la proporción de metabolitos circulantes, y paremos de contar. Obviamente deberíamos prescindir de tantos omas y tal vez prestar más atención a la red global. Pero lo que aquí nos atañe es que el modificoma (otro hilarante palabro) es parte fundamental del entramado, y con ello ya decimos lo suficiente. Pensándolo bien, hay otra razón de peso. Aunque tarde o temprano acabará eclipsándola, la proteómica siempre ha sido la hermana pequeña de la genómica. Y en este sobrevivir ante quien tiene más madurez y tamaño, la más pequeña se aferra a sus habilidades únicas. Y resulta que estas modificaciones, al no estar codificadas en el genoma, son coto de caza exclusivo del proteómico. Hay autores que afirman que la espectrometría de masas es la técnica idónea para identificar modificaciones. Parte de razón no les falta, si consideramos que los instrumentos actuales alcanzan sin despeinarse la resolución mínima de una unidad de

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
30
masa atómica. La detección de cualquier cambio, por pequeño que sea, en la masa del péptido o de la proteína está por tanto al alcance de estos aparatos. Y las modificaciones isobáricas casi nunca ocurren. Sin embargo, la exactitud de masa es, paradójicamente, una de las dificultades principales de la técnica, cuando se aplica a este fin concreto. La culpa la tienen, una vez más, los motores de búsqueda con que nos hartamos de identificar péptidos en las bases de datos a partir de sus espectros de fragmentación. Estos programas están concebidos para procesar números y secuencias, pero no para interpretar los números antes de hablar de secuencias. Cuando la modificación altera la masa del precursor, no hay nada que hacer pues no cuadran los números; sólo caben apaños y remiendos sobre modificaciones supuestas a priori. Y puesto que la proteómica actual no se concibe sin estos programas, la cruda realidad es que desconocemos cuál es la proporción de péptidos modificados; los espectros que no encajan se ignoran, y listos. Este proceder induce a creer que la inmensa mayoría de los péptidos están intactos y que nuestras enzimas tienen una selectividad exquisita.
Otro detalle que se olvida es que las proteínas están tan vivas como los organismos de los que forman parte. Y reaccionan por los cuatro costados. La complejidad química de una cadena polipeptídica es difícil de imaginar, sobre todo cuando algo queda de su estructura espacial, y el microentorno de los grupos próximos estimula la reactividad local. El más vivo de todos, la cisteína, puede adormecerse mediante la pertinente alquilación; sin embargo, poco es sabido que esta alquilación favorece su oxidación, al estilo de su colega la metionina. Y la
tirosina, la histidina, la lisina están también ahí, formando parte de la orquesta, anhelosas de relacionarse con electrófilos errantes. Otros grupos menos considerados, como el triptófano o la prolina, se oxidan o hidroxilan más fácilmente de lo que se piensa, y los grupos ácidos están siempre a la expectativa… Lo realmente sorprendente es que no detectemos ni por asomo un intricado amasijo de reacciones secundarias.
Así que ni sabemos cómo
detectar los péptidos modificados, de manera eficaz y de una vez por todas, ni conocemos su proporción, y encima ignoramos por completo cuán artefactuales son los protocolos de rutina. Tal vez debiéramos hacer un esfuerzo de imaginación preventiva. Frente a la concepción de una proteómica químicamente limpia e impoluta donde no hay sino toneladas de péptidos intactos y perfectos, con trazas de péptidos modificados postraduccionalmente en sitios fisiológicos y relevantes, deberíamos contraponer una visión más underground, plagada de péptidos oxidados o acetilados, aductos de sodio y otros metales y reacciones callejeras y parásitas de todo tipo.
Pero la carencia de aplicaciones
informáticas adecuadas sólo contribuye en parte a la complejidad del asunto. Otro primerísimo problema es la falta de cobertura, o imposibilidad de conseguir información sobre la secuencia completa de las proteínas. Y claro, la modificación que buscamos tendrá buen cuidado de situarse precisamente en la región invisible; ya sabemos que cuanto más importante, más se dejará querer, pues si no ya la habría identificado otro colega.
Para complicar el tema, la falta
de cobertura tiene no una, sino varias

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
31
causas. La accesibilidad de las proteasas a sus sitios de corte es con frecuencia caprichosa e impredecible, sobre todo si nuestro protocolo incluye -¿cuándo no?- una etapa electroforética. Y ninguna de las dos técnicas de ionización es perfecta; aunque casi todo, no todo lo que vuela en el MALDI salta de la gota en el electrospray. Aunque los catálogos comerciales se empeñen en lo contrario, no todos los péptidos se retienen en fases inversas, y algunos péptidos son particularmente pegajosos -el inglés sticky es más adecuado-, y se van quedando por el camino, adheridos a eppendorfs, puntas de pipeta, agujas Hamilton o líneas cromatográficas. Para colmo, no todas las proteasas son tan selectas como la tripsina, y hoy por hoy hay escasas alternativas limpias y fiables.
Hablamos de reacciones
secundarias; afortunadamente, la más buscada y trascendente de las modificaciones, la fosforilación, no ocurre sin ayuda catalítica. Y sólo la sulfatación, tan infrecuente como poco conocida, se le iguala en lo que a masa nominal se refiere. Por ello, identificar un péptido fosforilado in vivo gracias a un buen espectro de fragmentación constituye una experiencia del todo gratificante. Porque supone una demostración tan inequívoca de que esa modificación está en ese sitio y en plena célula, que es innecesario recurrir a técnicas paralelas para confirmar el resultado. Por desgracia, demostrar que la modificación desempeña un papel fisiológico y no es fruto de la acción de una quinasa despistada es harina de otro costal y requiere otras herramientas.
Una propiedad popular de los
fosfopéptidos es la de perder el grupo fosfato anclado a los residuos de serina y treonina, como sombrero en el vendaval, cuando se excitan en la célula
de colisión. Cualidad que a mi juicio habría que añadir a la larga lista de propiedades específicas de los fosfopéptidos, y que en la práctica no son tales, como la afinidad por quelantes metálicos, el bajo punto isoeléctrico o las reacciones de eliminación en beta. Propiedades que han dado lugar a la publicación de miríadas de métodos, a cuál más milagroso, para caracterizar selectivamente estas modificaciones. Lo anecdótico es lo elegantemente bien que funcionan cuando se aplican a la caseína beta. En la práctica no es válido el argumento de que si un científico danés fue capaz de identificar centenares de sitios de fosforilación en un proteoma entero, entonces identificar sólo uno en una única proteína, aislada y predispuesta, debería ser tan fácil como pulsar un botón. Pues más difícil puede resultar lo segundo que lo primero, y tal vez el fosfoproteoma no refleje las modificaciones más abundantes, sino las menos problemáticas.
Porque aunque no haya duda de
su presencia cuando se ven, es decir, cuando se fragmentan, el mero hecho de ver los fosfopéptidos resulta un más difícil todavía. Estos péptidos tienen un fuerte carácter que un germano hablante llamaría zwitteriónico, con un grupo fosfato que se resiste a ser protonado aunque forcemos por lo bajo el espectro de pH. Y la afamada pérdida de fosfato se convierte entonces en un enemigo de las series b y de los motores de búsqueda. El resultado son péptidos escurridizos y antipáticos que ponen a prueba la mucha dosis de paciencia que se le supone a la ciencia.
Una de las técnicas más
populares para analizar estos procesos es el uso de fósforo radiactivo. Se suele razonar que una fosforilación muy clara y abundante, a tenor de la intensidad de la mancha autorradiográfica, implica

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
32
que el fosfopéptido debería detectarse por espectrometría de masas. Nada más lejos de la realidad. Puede resultar engorroso reconocer que los radioisótopos permiten una sensibilidad varios órdenes de magnitud superior. Pero esta afirmación, que supone casi siempre enzarzarse en discusiones bizantinas, es simple de refutar: basta con utilizar un contador de centelleo y hacer un cálculo exacto de los moles de fosfato. Experiencia que sirve para practicar, y nunca viene mal, con el evasivo concepto de zeptomol.
La nitrosilación es otra modificación interesante que también parece tener un rol notable en un sinnúmero de procesos bioquímicos. Pero tiene la peculiaridad de no ser enzimática, y se involucra en un entramado que se debe considerar bajo una perspectiva oxidativa y nitrosativa. Hay algo de inefable y apasionante en ese proceso dinámico y continuo, en el que los tioles y grupos redox presentes en el medio actúan como al unísono, donando y recibiendo oxígenos, nitrógenos y radicales, en mayor o menor medida de acuerdo al estrés celular, como si las células también sufrieran las inclemencias del tráfico. Siempre me ha costado entender cómo esta evanescente cascada de reacciones puede influir en procesos tan tangibles como la hipertensión o el envejecimiento. También me resisto a imaginar la preferencia por determinados grupos en ausencia de directrices catalíticas. Hay detalles evidentes que se escapan, y que tendrán que ver con la estructura tridimensional y con las interacciones y el entorno en el que se sitúan las proteínas aceptoras. Lo que sube el listón de dificultad técnica hasta lo insospechable.
Ya he comentado que las
cisteínas son especialmente reactivas. Y esta propiedad ha sido muy aprovechada para desarrollar métodos para cuantificarlas, determinar su estado de oxidación, la formación de puentes disulfuro. Estos métodos, basados en el uso de reactivos de grupo sobre proteínas aisladas, se han adaptado para pescar, de forma específica, proteínas nitrosiladas en muestras muy complejas. Se dice que algunos, como el biotin-switch, son particularmente eficaces. Pero la realidad es que la bibliografía es muy parca en lo que a demostrar la existencia de sitios de nitrosilación se refiere y más aún sobre su relevancia fisiológica, que sólo se conoce en un número de casos que podrían contarse con los dedos de la mano. ¿Qué fundamento hay, pues, para hablar de eficacia? ¿No estaremos abusando del término especificidad?
La mayoría de nosotros somos
conscientes de casi todos estos problemas. ¿Por qué hay, entonces, tanto empeño en analizar estos cambios? No volveré al tópico de que estas cosas son relevantes, o se publican muy bien, que por otra parte es cierto. Adelantaré a cambio la hipótesis, que por supuesto no puedo probar, de que lo que nos atrae de ellos es su desesperante dificultad. El reto que suponen y el gozo de encontrarlos. Lo que entre científicos no podía ser de otra manera.
1El autor agradece a Horacio
Serrano y Antonio Martínez-Ruiz sus útiles sugerencias y comentarios.
.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
33
TESIS DOCTORALES

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
34
Análisis de los mecanismos moleculares
asociados al desarrollo de EHNA y HCC en un
modelo de ratón deficiente en el gen MAT1A
mediante proteómica funcional.
Enrique Santamaría
Unidad de proteómica, Fundación para la Investigación
Médica Aplicada (FIMA), Pamplona-
En este estudio, hemos investigado la progresión de la esteatohepatitis no-alcohólica (EHNA) y hepatocarcinoma (HCC) en un ratón Knock-out deficiente en la síntesis de S-adenosilmetionina (MAT1A-/-) mediante técnicas de proteómica de primera generación. Durante el desarrollo de EHNA, se han identificado 117 proteínas en el hígado del ratón MAT1A-/-. Doce de ellas, cambian de expresión desde el nacimiento del ratón hasta la aparición de lesiones histológicas. Cuatro de estas proteínas (Cadena β de la ATPasa mitocondrial, subunidades I y II de la Citocromo C oxidasa y Prohibitina I) están implicadas en el mantenimiento de la función mitocondrial, lo que sugiere un fallo mitocondrial en los estadios tempranos de la enfermedad. Estas alteraciones, también se han demostrado tanto en el hígado de ratones ob/ob como en pacientes obesos, que presentan una predisposición a sufrir EHNA. Además, se ha observado la sobreexpresión de una isoforma acídica de la Apolipoproteína A-I (Apo A-I) en el suero del ratón MAT1A-/- meses antes de la aparición de lesiones histológicas. La caracterización de esta isoforma diferencial mediante espectrometría de masas en tándem ha revelado la oxidación específica de la metionina 85
y 216 a metionina sulfóxido. También se ha observado un enriquecimiento de una isoforma acídica de Apo A-I en el suero de pacientes con Hepatitis B que posteriormente han desarrollado HCC. Esta isoforma presenta la oxidación específica de la metionina 112 a metionina sulfóxido y la oxidación a formilkinurenina de los residuos de triptófano 50 y 108. En el análisis de los nódulos tumorales presentes en el hígado de diferentes ratones, se ha observado que más del 10% de las proteínas (p<0.05) cambian su expresión mientras que la variabilidad observada en otros grupos experimentales es del 1% (p<0.01). Estos resultados sugieren que la heterogeneidad del proteoma no sólo deriva del proceso estocástico asociado al crecimiento del tumor, sino que existen factores específicos de cada individuo que participan en la inestabilidad del proteoma, ejerciendo una presión selectiva sobre los focos preneoplásicos. Se han identificado 151 proteínas expresadas diferencialmente en los tumores MAT1A-/- y 27 de ellas son comunes, como mínimo, al 50% de los tumores analizados. 13 de ellas se encuentran alteradas en HCC humanos, y de estas 13, 7 se encuentran alteradas en hígado cirrótico, por lo que este panel de potenciales biomarcadores

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
35
puede ser útil no sólo en la diagnosis, sino además en la detección precoz de estadios pre-tumorales.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
36
La Proteómica Aplicada al Estudio de las Respuestas de las Plantas a Estreses Bióticos y
Abióticos
Mari Angeles Castillejo Sanchez Departamento de Bioquímica y Biología Molecular,
Universidad de Córdoba
Las plantas están expuestas a lo
largo de su ciclo de vida a condiciones externas adversas, lo que afecta a su desarrollo, reduciendo la productividad de numerosos cultivos. Mediante una aproximación de proteómica se han estudiado las bases moleculares de la respuesta de las plantas a estreses abióticos y bióticos, utilizando para ello los siguientes sistemas experimentales:
i) La planta modelo Medicago truncatula, en la que se han abordado estudios de respuesta a hongos fitopatógenos (Uromyces striatus, roya) y a plantas parásitas (Orobanche crenata, jopo). ii) La leguminosa Pisum sativum (guisante), en la que se ha
estudiado la respuesta a plantas parásitas (Orobanche crenata, jopo). iii) La oleaginosa Helianthus annuus (girasol), en la que se han llevado a cabo estudios de respuesta a sequía. Para ello, se ha comparado el
perfil proteico de variedades de Medicago truncatula, Pisum sativum y Helianthus annuus que presentan diferente grado de resistencia/tolerancia frente a dichos estreses, mediante una combinación de electroforesis bidimensional (2DE) y análisis por espectrometría de masas (MALDI-TOF). El análisis comparativo en cada uno de los sistemas estudiados ha revelado la existencia de diferencias en el patrón de expresión proteico de raíces y hojas entre genotipos. Así mismo se han observado cambios en el mapa proteico en respuesta a la inoculación con Uromyces striatus (Medicago truncatula) y Orobanche crenata (Medicago truncatula y Pisum Sativum) y como consecuencia del tratamiento de sequía (Helianthus annuus). Las proteínas expresadas diferencialmente entre genotipos y tratamientos se han identificado mediante espectrometría de masas y búsquedas en bases de datos de proteínas o DNA utilizando para ello motores de búsqueda (algoritmos).

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
37
Los resultados obtenidos validan la aplicación de la metodología utilizada para identificar proteínas y genes de respuesta a sequía, hongos fitopatógenos y plantas parásitas y caracterizar los mecanismos de resistencia y tolerancia frente a estos estreses. Entre las proteínas identificadas se encuentran tanto proteínas de defensa y de respuesta a estrés, como enzimas del metabolismo de carbohidratos y fotosíntesis, pudiéndose diferenciar entre mecanismos de resistencia constitutivos e inducidos, y respuestas generales y específicas de especie o estrés. Así, en el sistema Medicago truncatula-Uromyces striatus se observa un número reducido de proteínas diferencialmente expresadas. En relación a la respuesta a O. crenata se observan dos tendencias claras entre los genotipos estudiados: mientras que en los genotipos susceptibles de ambas plantas (Medicago truncatula y Pisum sativum) se observa una disminución general de las enzimas del metabolismo energético, en los genotipos resistentes se aprecia un incremento de proteínas de defensa frente a patógenos y proteínas de respuesta a estrés. En girasol, el tratamiento de sequía produce principalmente, una disminución de las enzimas de la fotosíntesis y del metabolismo de carbohidratos en el genotipo susceptible que no se aprecia en el resistente.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
38
Análisis del proteoma de suero humano para la identificación de proteínas con utilidad clínica en el
cáncer colorrectal: Estudio proteómico de las isoformas de clusterina
Ana María Rodríguez Piñeiro
Departamento de Bioquímica, Genética e Inmunología, Universidad de Vigo
El pasado 16 de diciembre de 2005
fue presentada la Tesis Doctoral “Análisis del proteoma de suero humano para la identificación de proteínas con utilidad clínica en el cáncer colorrectal: Estudio proteómico de las isoformas de clusterina”, defendida por Dña. Ana María Rodríguez Piñeiro y merecedora de la máxima calificación. Dicho trabajo fue realizado en el Departamento de Bioquímica, Genética e Inmunología de la Universidad de Vigo, bajo la dirección de los Dres. María Páez de la Cadena Tortosa y Francisco Javier Rodríguez Berrocal.
El cáncer colorrectal (CCR) constituye una de las neoplasias más frecuentes en la sociedad occidental, siendo la tercera en cuanto a su incidencia y la segunda por su mortalidad. Por ello, para conseguir un correcto diagnóstico, pronóstico y seguimiento de la patología es necesario buscar nuevos marcadores tumorales. Considerando que la proteómica es una de las áreas científicas más prometedores en esta búsqueda, en el trabajo presentado se han empleado diversas metodologías proteómicas y bioquímicas, con el fin de detectar marcadores tumorales séricos de utilidad para el CCR.
El amplio rango dinámico de las proteínas séricas es uno de los mayores inconvenientes del análisis del proteoma sérico completo. Por ello, se empleó un método de prefraccionamiento mediante cromatografía de afinidad a través de Concanavalina A. Como se observa en la figura 1, esta separación previa permite la obtención de una fracción constituida mayoritariamente por proteínas no glicosiladas u O-glicosiladas (FI), y de otra fracción enriquecida en N-glicoproteínas (FII). El análisis mediante electroforesis bidimensional de estas fracciones permite la detección de un número más elevado de proteínas séricas, obteniéndose mayor resolución con respecto al análisis del proteoma sérico en conjunto (figura 2). En particular, la fracción de N-glicoproteínas es una muestra adecuada para la búsqueda de marcadores tumorales, ya que se han descrito numerosas alteraciones en los procesos de N-glicosilación en la carcinogénesis. Una ventaja añadida al uso de esta fracción es que no contiene albúmina sérica, la proteína no glicosilada más abundante en suero, lo que permite obtener mejor resolución y mayor sensibilidad de detección en los mapas bidimensionales de N-glicoproteínas

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
39
Figura 1. Perfiles representativos de la separación de sueros controles y tumorales a través de Con A-Sefarosa. FI (fracción I): proteínas no retenidas por la lectina; FII (fracción II): proteínas unidas específicamente a la matriz de Con A; α-MM: α-metilmanósido; DO: densidad óptica.
Figura 2. Mapas representativos de la separación de un suero control y de las fracciones obtenidas mediante cromatografía de afinidad a través de Con A: FI (mayoritariamente proteínas no glicosiladas y O-glicoproteínas) y FII (enriquecida en N-glicoproteínas). En estas imágenes se encontraron 1.376 spots en suero, 841 en FI y 1.185 en FII. En los mapas se han señalado algunas proteínas de interés para la caracterización del contenido de cada muestra. Mr: masa molecular relativa; pI: punto isoeléctrico; AACT: α-1-antiquimotripsina; A2HS: α-2-HS-glicoproteína; LRG: α-2-glicoproteína rica en leucina; Ig: inmunoglobulina; Apo-AI: apolipoproteína A-I; RPB: proteína plasmática de unión a retinol.
En la búsqueda de marcadores tumorales utilizando mapas bidimensionales de proteínas existen tres aproximaciones posibles. La primera consiste en la detección de especies proteicas que presenten diferencias en sus niveles de expresión entre grupos de población. De este
modo, se detectaron 72 proteínas alteradas en relación a la patología, 37 con expresión incrementada y 35 con expresión disminuida (figura 3), de las cuales se identificaron aproximadamente un 50% mediante espectrometría de masas. Estas proteínas están implicadas en diversos

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
40
procesos fisiológicos, como la coagulación (antitrombina III, cadena B del factor de coagulación XIII, apolipoproteína H), la regulación de la homeostasis sanguínea (α-1-antiquimotripsina, haptoglobina), la respuesta inmunitaria (inmunoglobulinas, factor I del complemento), los fenómenos de inflamación y fase aguda (α-1-
microglobulina), y el metabolismo lipídico (α-2-glicoproteína de zinc, apolipoproteína A-I). Por otra parte, también se identificaron proteínas relacionadas con la apoptosis (clusterina), y la transducción de señales (α-2-glicoproteína rica en leucina), de mayor interés como potenciales candidatos a marcadores tumorales para el CCR.
Figura 3. Mapa estándar de la comparación de FII donde se destacan los 51 spots alterados significativamente en pacientes con CCR con respecto a individuos sanos (incrementos en color rojo y disminuciones en color azul).
Una segunda aproximación para la búsqueda de marcadores tumorales consiste en utilizar los patrones de expresión en los mapas bidimensionales como herramienta diagnóstica. Empleando el análisis de componentes principales (figura 4) y el análisis discriminante, se pudo establecer un patrón de expresión diferencial para cada uno de los grupos de población. Por lo tanto, en cualquiera de los dos tipos de análisis se podría incluir un individuo cuyo estado en relación al CCR (sano o paciente) fuera desconocido y clasificarlo de acuerdo a los patrones preestablecidos.
Una tercera aproximación a la búsqueda de marcadores tumorales,
complementaria a las anteriores, es la detección de cambios en la posición relativa de proteínas en mapas bidimensionales. Estos cambios son el reflejo de variaciones en el punto isoeléctrico y la masa molecular, que pueden ser debidas a los procesos cancerosos. En este estudio se aplicó un método morfométrico geométrico, denominado análisis de deformaciones relativas, que permite la detección de variaciones locales de posición, eliminando el efecto que puedan causar los errores técnicos en el patrón de separación global. El análisis empleado permitió detectar variaciones significativas en la posición de N-glicoproteínas séricas de pacientes con

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
41
CCR en relación a las de individuos sanos (figura 5). Esta técnica puede servir como herramienta diagnóstica, puesto que permite la discriminación entre individuos sanos y enfermos. Además, el método permitió detectar la contribución de cada proteína a la distinción de los grupos. Entre las que
presentaban mayor efecto se identificaron el angiotensinógeno, la α-1-microglobulina, la apolipoproteína A-I, y ciertas isoformas de haptoglobina.
Figura 4. Representaciones gráficas del análisis de componentes principales. Tanto en una representación 2D junto con el segundo componente (CP2’), como en una gráfica 3D junto con el segundo y el tercero (CP2’ y CP3’), se observa que la separación dada por el primer componentes principal (CP1’) agrupa a los pacientes con CCR, separándolos de los individuos control. En ambas gráficas destaca la mayor variabilidad del grupo control.
Figura 5. Representación de los valores de desviación relativa (RW3) como vectores (reescalados 10x para visualizar las diferencias) que definen el movimiento neto de las manchas proteicas.
Componente principal 2'
2,52,01,51,0,50,0-,5-1,0-1,5
Com
pone
nte
prin
cipa
l 1'
1,5
1,0
,5
0,0
-,5
-1,0
-1,5
GRUPO
2
1
GRUPO
Paciente
Control
23
-1,0
-,5
0,0
12
,5
1,0
1,5
01 -10 -2-1
CP 1’
CP 2’ CP 3’

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
42
Uno de los potenciales marcadores tumorales identificados en los estudios anteriores, de mayor interés por su implicación documentada en diversos procesos neoplásicos, es la clusterina. Esta proteína presenta formas tisulares citoplasmáticas y nucleares, siendo también secretada a fluidos biológicos como el suero. Su función exacta todavía es cuestión de controversia, aunque una de sus funciones más relevantes se relaciona con la apoptosis, ejerciendo funciones tanto pro- como antiapoptóticas.
En este trabajo se han estudiado las diversas formas séricas de la clusterina, comparando su distribución y niveles en el suero de individuos sanos y de pacientes con CCR. Se empleó el prefraccionamiento de suero a través de Concanavalina A, seguido de técnicas electroforéticas nativas y desnaturalizantes, tanto en una como en dos dimensiones. En condiciones nativas, en el suero total y en la fracción cromatográfica FII se detectaron el heterodímero de 70 a 85 kDa, según su grado de glicosilación, y diversas formas de alto peso molecular que podrían corresponder a dímeros o agreagados. La aplicación de un tratamiento desnaturalizante permitió detectar las subunidades de la proteína con una masa de 40 kDa, así como bandas de ∼75 a 80 kDa correspondientes a la holoproteína, total o parcialmente glicosilada. El hallazgo más destacable fue la detección, exclusivamente en pacientes con CCR, de una forma de clusterina eluida en la primera fracción cromatográfica (FI CLU), constituida por al menos 5 isoformas, con una masa nativa de 85 kDa y subunidades de 40 kDa. La desglicosilación de la FI CLU permitió comprobar que era una N-glicoforma, probablemente con una glicosilación aberrante. Por otra parte, en FII se detectó la disminución de expresión de 15 N-glicoformas, y el aumento de una.
El análisis de la composición glucídica de 5 de estas isoformas no permitió detectar variaciones significativas del patrón de glicosilación, aunque sí existen diferencias en los índices de sialilación y de fucosilación de isoformas específicas.
Una vez realizados los estudios proteómicos, se evaluó la utilidad clínica de la clusterina sérica para el CCR. En primer lugar, se desarrolló una técnica de ELISA para la cuantificación de los niveles séricos de clusterina total. La comparación de los valores en 31 sueros de individuos sanos y en 31 sueros de pacientes con CCR permitió detectar un incremento significativo de 1,6 veces en estos últimos. Para la valoración de clusterina en las fracciones cromatográficas, se desarrolló un método de inmunodetección por slot blot, aplicándolo a las muestras de 10 individuos sanos y de 10 pacientes (figura 6). Así, se corroboró el incremento significativo de clusterina sérica total en pacientes, y se observó una disminución de formas eluidas en FII que no alcanzaba la significación estadística. El resultado más destacable fue la elevación de más de 4 veces de los niveles de FI CLU en pacientes con CCR, que corrobora los resultados obtenidos en los estudios proteómicos.
En relación al valor clínico de la clusterina sérica, se determinaron los parámetros diagnósticos de la clusterina sérica total, encontrando dos posibles puntos de corte. Uno de ellos presentó una eficiencia superior al marcador CEA, de uso establecido en clínica. Sin embargo, sus bajos niveles de sensibilidad no permiten recomendar su utilización como marcador de diagnóstico.
Por último, el hallazgo clínico más importante fue la relación de la FI CLU

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
43
con la metástasis. Teniendo en cuenta los datos clínicos y anatomopatológicos, se comprobó estadísticamente que un valor igual o superior a un 4% de FI CLU en el suero preoperatorio, con respecto a la clusterina sérica total, permite la clasificación correcta de 9 de los 10 casos examinados en relación a la presencia de metástasis. Además, existe una correlación significativa entre los niveles de FI CLU y la distancia a la
que se localizan los nuevos tumores con respecto al primario. Por ello, esta forma de clusterina puede constituir un valioso marcador de pronóstico para pacientes con CCR.
De los resultados obtenidos en este trabajo se concluye que el estudio de isoformas específicas de clusterina puede ser más informativo en relación al CCR que el análisis de clusterina sérica total.
Figura 6. Niveles de clusterina en suero total y en las fracciones cromatográficas FI y FII en relación a la cantidad total de proteínas de la muestra. Los histogramas representan el promedio con la desviación estándar (barras) para los 10 individuos de cada grupo poblacional. UA: unidades arbitrarias; *: P < 0,05.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
44
Desarrollo de Nuevas Metodologías para
el Análisis del Proteoma
Daniel López-Ferrer
Laboratorio de Proteómica y Química de Proteínas
Centro de Biología Molecular Severo Ochoa
La tesis se encuadra dentro de las líneas de investigación abiertas en nuestro laboratorio en los últimos años. Entre otras, el desarrollo de técnicas de segunda generación para el análisis de proteomas. El objetivo era desarrollar métodos para realizar análisis a gran escala de proteínas basados en espectrometría de masas, especialmente utilizando trampas iónicas por su capacidad de trabajo como caballos de batalla. Como la premisa fundamental fue conseguir análisis robustos apostamos por una aproximación “Bottom-Up” para analizar proteomas, por las ventajas de sobra conocidas, que supone trabajar con péptidos en lugar de con proteínas.
Los desarrollos que se describen en
esta memoria engloban todos y cada uno de los pasos que se realizan en un experimento a gran escala en proteómica, desde la preparación de la muestra, el análisis de datos, estudios de expresión diferencial y finalmente el estudio de modificaciones post-traduccionales.
A lo largo de los capítulos se van enumerando los distintos desarrollos junto a las publicaciones a las que han dado lugar. En los dos primeros, a modo de introducción, se aporta primero el marco teórico en donde podríamos englobar la proteómica dentro del nuevo paradigma que supone la biología de sistemas y posteriormente, hacemos una revisión bibliográfica bastante detallada de la aportación de la espectrometría de masas a la proteómica1.
En cuanto se refiere a la parte experimental, entramos de lleno con la aplicación de un nuevo protocolo de preparación de muestra basado en la utilización de ultrasonidos focalizados durante el proceso de la hidrólisis enzimática de proteínas, esta combinación nos permite acelerar los tiempos de digestión de horas a minutos, con la ventaja que supone en las aplicaciones a gran escala2.
Otro de los avances que proponemos, enmarcado en el proceso de la prospección de datos, es el diseño de un modelo estadístico empírico que permite el análisis de los datos de forma semiautomática y al mismo tiempo de una manera rápida y robusta, con el objetivo de evaluar el error cometido en la identificación de péptidos3. Al mismo tiempo, proponemos, a partir de un estudio detallado del comportamiento de las distribuciones de puntuaciones del motor de búsqueda SEQUEST, un diseño eficiente de las condiciones de búsqueda con la finalidad de ahorrar tiempo y aumentar la sensibilidad de ésta.
Terminando con la parte de proteómica de segunda generación, demostramos la alta sensibilidad, precisión y solidez de la utilización de marcaje enzimático con H2
18O y espectrometría de masas de trampa lineal, para realizar estudios de expresión diferencial a partir de experimentos de dilución isotópica4, y como la utilización de los ultrasonidos focalizados en el proceso de marcaje acelera el proceso de días a minutos.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
45
Para concluir, se muestra además, el desarrollo llevado a cabo para la caracterización y monitorización directa de modificaciones postraduccionales de proteínas, tales como nitrosilación, en muestras biológicas complejas5,6.
Con todas sus limitaciones y
asumiendo el peligro de generalizar, creemos que esta tesis ha intentado, desde un punto de vista trandisciplinar, abarcar la mayor parte de los problemas que atañen el análisis proteómico. Poniendo de manifiesto, que el científico proteómico no es un especialista estricto, pero sí posee una visión global que le permite resolver problemas de forma eficiente aunando conocimientos de disciplinas muy diferentes.
Bibliografía
1. Canas B, López-Ferrer D, Ramos-Fernández A, Camafeita E, Calvo E. (2006). Mass spectrometry technologies for proteomics. Briefings in Functional Genomics and Proteomics. 4. 295-320
2. Lopez-Ferrer D, Capelo JL, Vazquez J. (2005). Ultra fast trypsin digestion of proteins by high intensity focused
ultrasound. J Proteome Res. 4. 1569-74.
3. Lopez-Ferrer D, Martinez-Bartolome S, Villar M, Campillos M, Martin-Maroto F, Vazquez J. (2004). Statistical model for large-scale peptide identification in databases from tandem mass spectra using SEQUEST. Anal Chem. 76. 6853-60
4. Lopez-Ferrer D, Ramos-Fernandez A, Martinez-Bartolome S, Garcia-Ruiz P, Vazquez J. (2006). Quantitative proteomics using 16O/18O labeling and linear ion trap mass spectrometry. Proteomics. 2006 6 Suppl 1:S4-S11.
5. Villar M, Ortega-Perez I, Were F, Cano E, Redondo JM, Vazquez J. (2006). Systematic characterization of phosphorylation sites in NFATc2 by linear ion trap mass spectrometry. Proteomics. 6 Suppl 1:S16-27.
6. Martinez-Ruiz A, Villanueva L, Gonzalez de Orduna C, Lopez-Ferrer D, Higueras MA, Tarin C, Rodriguez-Crespo I, Vazquez J, Lamas S. (2005). S-nitrosylation of Hsp90 promotes the inhibition of its ATPase and endothelial nitric oxide synthase regulatory activities. Proc Natl Acad Sci U S A. 102. 8525-30.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
46
LUDOPROTEÓMICA: EL RINCÓN DEL SOCIO

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
47
Poetómica
Jesús Vázquez
Ascensión de la proteína
Esta proteína aleve que en la membrana encalla tiene, sí, su isoeléctrico punto tan desplazado que más que migrar vuela, tal campo de batalla fuera el gel inocente, rabiosa hacia su cátodo.
Se sale la proteína y asesina mi gozo
sustentado en artículos de astronómico impacto donde ella sería mi invento y mi alborozo,
mi trampolín de fama, mi medio de contacto.
Y yo que ya pensaba medir su raudo vuelo, atraparla y contar sus enlaces peptídicos,
glosar su homología -tan alto era mi anhelo- y discutir sus roles, ¿utópicos? ¿Verídicos?
¡Qué suculento alijo de péptidos daría,
tan luenga y tan repleta de azufres y de bases! ¡Qué lista de fragmentos sin par, en sintonía con su papel en cáncer, telómeros, profases!
Sospechan que es sustrato de frágiles quinasas,
transloca si se altera la tirosina veinte... ¡Qué ciega apoteosis de espectros y de masas
sería demostrarlo en vivo y de repente!
Para colmo regula la NO sintasa en endotelio, dicen... ¿Qué no haría esta perla,
oh joya, no va más desmedido y sin tasa para el voraz proteómico? ¡Sólo es cuestión de verla!
Mas la proteina escapa, sólo queda un fantasma de manchas y de iones cercanos mas sin dueño.
Sólo queda un proyecto cuyo cántaro plasma en pedazos la historia repetida de un sueño.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
48
SERVICIOS Y
UNIDADES DE
PROTEOMICA

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
49
1. INTRODUCCIÓN
El Centro de Investigación Príncipe Felipe
(CIPF) nace en 2002 como la mayor apuesta realizada hasta el momento por la Generalitat Valenciana en el campo de la investigación biomédica. El CIPF es un Centro dedicado, fundamentalmente, a la investigación en los campos de la Medicina Regenerativa, y del descubrimiento de nuevos fármacos, teniendo carácter de Centro Asociado al Instituto de Salud Carlos III como Centro Nacional de Referencia para la Investigación en Biomedicina y en transplantes de Tejidos y Órganos. Integrado por un equipo de más de 260 investigadores y personal de apoyo cuyo trabajo está orientado hacia:
• La derivación de células troncales embrionarias.
• La diferenciación de células troncales en diversos tipos celulares y tejidos.
• La terapia celular. • El descubrimiento de nuevos
compuestos con actividad terapéutica, y su desarrollo hasta las fases preclínicas.
• El descubrimiento y validación de nuevas dianas terapéuticas.
El CIPF cuenta con un banco de líneas celulares troncales embrionarias; plataformas de citómica, proteómica, bioinformática, genómica, química médica; y un animalario para distintas especies. El Servicio de Proteómica del CIPF empezó a ponerse en marcha en diciembre de 2004 y a funcionar
como servicio al público en junio de 2005. Dada la reciente creación de este Servicio, y del centro en su conjunto, todavía hay aspectos que están por desarrollar, como son la página web, etc.
2. OBJETIVOS
Nuestros objetivos fundamentales son proporcionar un servicio de identificación de proteínas y, mediante el empleo de la espectrometría de masas, dar apoyo a distintos proyectos de investigación. Además, el Laboratorio de Proteómica está interesado en el desarrollo y/o puesta a punto de protocolos menos convencionales que puedan ser de utilidad a los investigadores.
SERVICIO DE PROTEÓMICA CENTRO DE INVESTIGACIÓN PRÍNCIPE FELIPE
Manuel M. Sánchez del Pino Departamento de Química Médica, Centro de Investigación
Príncipe Felipe, Avenida Autopista del Saler 16, 46013 Valencia

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
50
3. PERSONAL
Para cumplir los objetivos mencionados, el Laboratorio de Proteómica se ha dotado de una estructura con una dirección científica, a cargo de Manuel M. Sánchez del Pino, y una dirección técnica, llevada a cabo por Luz Valero. El resto del personal técnico está integrado por Carmen Aguado y Ester Dionís. La dirección científica se responsabiliza del funcionamiento general del Laboratorio de Proteómica. Debe decidir las metodologías y los servicios que, por su interés para el centro, deben implementarse. Los proyectos de investigación que se realicen en el Laboratorio de Proteómica, tanto propios como fruto de colaboraciones con grupos de investigación dentro y fuera del centro, serán seleccionados y dirigidos por el responsable científico. La dirección técnica se responsabiliza de los aspectos metodológicos y de funcionamiento diario del Laboratorio de Proteómica. Debe establecer los protocolos experimentales adecuados para optimizar el rendimiento de los instrumentos del Laboratorio. Asimismo, debe establecer los mecanismos de control necesarios que garanticen reproducibilidad y fiabilidad de los resultados y que permitan trazar las causas de posibles anomalías. También es tarea de la dirección técnica determinar el flujo de trabajo que conduzca a la mejor eficacia y aprovechamiento posible de los recursos. La función del personal técnico no es la de meros operadores de instrumentos o protocolos sino que son capaces de entender los fundamentos de su funcionamiento de los mismos. Por tanto, el análisis y interpretación técnica de los resultados es responsabilidad suya. Asimismo, dada su formación, están capacitados para participar activamente en proyectos de investigación, lo que sirve de motivación y estímulo. 4. SERVICIOS
En la actualidad ofrecemos distintos servicios de separación e identificación de proteínas:
• Separación de proteínas mediante
electroforesis bidimensional. • Electroforesis 2D diferencial
cuantitativa mediante metodología DIGE.
• Identificación de proteínas por huella peptídica mediante MALDI TOF-TOF.
• Identificación de proteínas en mezclas de baja complejidad mediante nanoLC MS/MS.
• Identificación de proteínas en mezclas complejas mediante 2DnanoLC MS/MS.
• Determinación de peso molecular mediante MALDI TOF o NanoESI QqTOF.
• Secuenciación de novo de péptidos mediante MALDI TOF-TOF o NanoESI QqTOF.
Otros servicios como identificación de
modificaciones postraduccionales, separación de proteínas mediante isoelectroenfoque en solución, análisis de complejos proteicos mediante electroforesis bidimensional Blue Native, etc. también pueden realizarse aunque no están tan estandarizados como los anteriores. 5. INSTRUMENTACIÓN
Para realizar los servicios mencionados contamos con los siguientes equipos: Electroforesis bidimensional y DIGE. Disponemos de los sistemas de GE Healthcare:
• Ettan IPGPHOR II IEF para la 1ª dimensión.
• Ettan DALTsix y Hoefer SE 600 Ruby para la 2ª dimensión.
• Typhoon™ 9400 para la detección de los marcadores fluorescentes.
• DeCyder™ para el análisis de los geles.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
51
Cromatografía. Disponemos del sistema cromatográfico de LC Packings compuesto de:
• Bomba nano HPLC UltiMate™ • Auto inyector Famos • Módulo MDLC Switchos • Micro colector de fracciones Probot
Procesamiento de muestras. Disponemos de los siguientes robots de Genomic Solutions:
• Investigator ProPic™ para la extracción de fragmentos de gel de poliacrilamida.
• Investigator ProPrep™ para la manipulación y digestión de los fragmentos de poliacrilamida y para la preparación de las placas de muestras para el espectrómetro de masas MALDI.
Espectrómetros de masas. Disponemos de los siguientes sistemas:
• 4700 MALDI-TOF/TOF de Applied Biosystems. Contiene el software necesario para realizar LC/MALDI.
• QSTAR XL MS/MS de Applied Biosystems. Espectrómetro híbrido
cuadrupolo-tiempo de vuelo. Asociado al espectrómetro hay un sistema de nanoHPLC Ultimate de LC Packings.
• LC/MS ZQ4000 de Waters/Micromass. Este sistema de cromatografía líquida convencional con detector de masas de simple cuadrupolo utilizado para el análisis rutinario de compuestos de síntesis de química orgánica y péptidos.
6. CONTACTO
Información del CIPF se puede encontrar en la dirección www.cipf.es. La página del Servicio de Proteómica está en proceso de construcción. Las direcciones de correo electrónico del personal del Servicio de Proteómica del CIPF son las siguientes:
• Manuel M. Sánchez del Pino: [email protected]
• Luz Valero: [email protected] • Carmen Aguado: [email protected] • Ester Dionís: [email protected]

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
52
La Unidad de Proteómica del Servicio
General de Investigación de Genómica y Proteómica de la Universidad del País Vasco comienza a funcionar en el año 2005 para dar respuesta a la demanda de este tipo de servicio por parte de grupos de investigación de la Universidad y otros centros de investigación y empresas del entorno.
En la actualidad somos cuatro las personas involucradas en el Servicio. Kerman Aloria es el técnico superior que se encarga del día a día del servicio. Kerman se licenció en Biología en la Universidad del País Vasco, hizo la tesis doctoral sobre plegamiento de tubulinas en la Universidad de Cantabria y se ha incorporado al Servicio de Proteómica después de realizar una estancia post-doctoral de cuatro años en la Universidad de Wisconsin-Madison, donde trabajó en proteínas implicadas en la homeostasis de hierro en mitocondrias. Miren Josu Omaetxebarria se licenció en Biología y Bioquímica en la Universidad del País Vasco, donde hizo la tesis doctoral sobre antígenos de Candida albicans. Tras realizar una estancia post-doctoral de algo más de dos años en la Universidad de Odense familiarizándose con las técnicas de espectrometría de masas aplicadas a proteómica, se ha incorporado a nuestro grupo con un contrato post-doctoral y su función es poner a punto nuevas técnicas para que puedan ser ofertadas por el Servicio. Asier Fullaondo se licenció y doctoró en la Universidad del País Vasco donde es profesor
del Departamento de Genética. En los últimos dos años ha realizado estancias en el laboratorio de Albert Heck en Utrech. Jesus M. Arizmendi también se licenció y doctoró en la Universidad del País Vasco, tras lo cual realizó una estancia post-doctoral en el laboratorio de John Walker en el MRC Laboratory of Molecular Biology de Cambridge. Es profesor del Departamento de Bioquímica y Biología Molecular y responsable de la Unidad de Proteómica del Servicio General de Genómica y Proteómica de la Universidad del País Vasco
El Servicio dispone de un sistema LC-
MS/MS de Waters-Micromass compuesto por un CapLC, un espectrómetro de masas Q-TOF micro y la plataforma informática MassLynx y ProteinLynx Global Server. El CapLC nos permite hacer separaciones cromatográficas de los péptidos con columnas capilares a flujos de 250 nl/min. El Q-TOF micro es capaz de realizar, entre otras aplicaciones, experimentos de MS/MS de
UNIDAD DE PROTEÓMICA DEL SERVICIO GENERAL DE INVESTIGACIÓN DE GENÓMICA Y PROTEÓMICA DE LA
UNIVERSIDAD DEL PAÍS VASCO Kerman Aloria, Miren Josu Omaetxebarria, Asier
Fullaondo, Jesus M. Arizmendi Facultad de Ciencia y Tecnología, Sarriena s/n, 48940 Leioa

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
53
forma automática en modo ‘Data Directed Analysis’, y gracias al sistema NanoLockSpray obtenemos datos con una exactitud de masa de 5 ppm. La plataforma informática, además de controlar tanto el sistema de HPLC como el espectrómetro de masas, procesa los datos crudos obtenidos, generando listas de picos que son utilizados en la búsqueda en distintas bases de datos por medio del motor de búsqueda del propio software y de Mascot. Además, el software tiene aplicaciones para la estimación de la masa molecular de moléculas múltiplemente cargadas, tales como proteínas.
Gracias a este equipamiento y personal, en
la actualidad se ofertan tres tipos de análisis:
• Determinación de masas molecular de proteínas purificadas
• Identificación de proteínas a partir de muestras purificadas
• Identificación de proteínas a partir de mezclas no muy complejas
Las tarifas y detalles del servicio se
pueden consultar en la siguiente página web www.ehu.es/vicinves/index_serviciosgenerales.htm o contactando con [email protected] o [email protected]
Durante el poco tiempo que llevamos funcionando hemos dado servicio a grupos de
investigadores de diferentes departamentos de la Universidad del País Vasco, así como a centros tecnológicos del País Vasco y grupos de Universidades próximas. Además, se están analizando muestras del proyecto HUPO de hígado liderado por el CIC bioGUNE.
De cara al futuro, pretendemos ir ampliando la lista de servicios ofertados. Concretamente, estamos poniendo a punto técnicas para el análisis de mezclas más complejas de proteínas y para el análisis de proteínas fosforiladas, y estamos interesados en técnicas de cuantificación sin el empleo de 2DE.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
54
La Unidad de Proteómica del CNIC se creó en 2003 como un servicio de proteómica general abierto a toda la comunidad científica, poniendo a disposición de cualquier laboratorio que lo solicite una amplia gama de servicios con sus correspondientes tarifas. La Unidad ofrece su experiencia no sólo en técnicas consolidadas en el área de la proteómica, sino también en nuevas técnicas aún no disponibles en otros laboratorios. Asimismo, la Unidad imparte periódicamente cursos prácticos para el aprendizaje y aplicación de dichas técnicas. 1. EQUIPAMIENTO DE LA UNIDAD DE PROTEÓMICA DEL CNIC
i) Sistema completo de electroforesis bidimensional (para geles de hasta 26 cm de longitud).
ii) Sistema completo para análisis diferencial cuantitativo de geles bidimensionales premarcados con fluoróforos (DIGE, GE Healthcare), con Typhoon 4100 y DeCyder v6.5 y módulo EDA.
iii) Estación robótica para el picado de manchas en geles (SpotPicker, GE Healthcare).
iv) Estación robótica para la digestión de proteínas en gel y muestreado de digeridos en placas de MALDI (Proteineer DP, Bruker).
v) Espectrómetro de masas MALDI-TOF/TOF (Ultraflex TOF/TOF, Bruker) equipado con módulo LIFT
para la aceleración de iones fragmento.
vi) Espectrómetro de masas ESI-trampa iónica cuadrupolar, compuesto por nHPLC de LC Packings (Ultimate nanoLC, FAMOS Micro Autosampler y Switchos Micro Column Switching Module) y trampa Bruker Esquire HCT.
vii) Espectrómetro de masas 4000-Q Trap, compuesto por nHPLC de LC Packings (Ultimate nanoLC Plus, FAMOS Micro Autosampler y Switchos Micro Column Switching Module) y triple cuadrupolo modificado de Applied Biosystems.
2. SOLICITUD DE LOS SERVICIOS DE PROTEÓMICA
En nuestra página web (http://www.cnic.es/proteomica) se describe la forma adecuada de solicitar los servicios, así como las condiciones de preparación de la muestra y las tarifas aplicadas. Ciertos servicios adicionales no descritos aquí, al igual que determinados proyectos que por sus particularidades pueden requerir una excesiva demanda de recursos y tiempo, deben ser previamente consultados con el personal de la Unidad. 3. SERVICIOS DE PROTEÓMICA
Con el nivel de equipamiento actual y la experiencia del personal de la Unidad, se
UNIDAD DE PROTEÓMICA. FUNDACIÓN CENTRO NACIONAL DE INVESTIGACIONES CARDIOVASCULARES
CARLOS III Juan Antonio López del Olmo, Emilio Camafeita,
Enrique Calvo Unidad de Proteómica. Fundación CNIC. Melchor Fernández
Almagro, 3. 28029 - Madrid

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
55
ofertan una amplia gama de servicios relacionados con la separación e identificación de proteínas. 3.1. Separación de extractos proteicos por electroforesis bidimensional
La electroforesis bidimensional es la herramienta más comúnmente utilizada para el análisis de mezclas complejas de proteínas extraídas de células, tejidos o cualquier tipo de muestra biológica. La separación se hace inicialmente en función del punto isoeléctrico de las proteínas y, en una segunda fase, de acuerdo con su masa molecular. 3.2. Análisis de la expresión diferencial de proteínas por electroforesis bidimensional con fluoróforos premarcados (2D-DIGE)
El análisis comparativo de geles bidimensionales clásicos presenta el problema de la variabilidad entre geles. La electroforesis diferencial en gel (DIGE, GE Healthcare) viene a paliar en buena medida esta deficiencia, mejorando la reproducibilidad y la capacidad de extraer información cuantitativa de los análisis. El sistema permite la cuantificación estadística de pequeñas variaciones en los niveles de expresión de proteínas entre distintas muestras, las cuales son posteriormente analizadas para su identificación y caracterización. 3.3. Recorte de proteínas en gel automatizado
La automatización del recorte de proteínas de geles es muy útil para los proyectos que implican el recorte sistemático de todas las proteínas de un gel, y necesario cuando las proteínas de interés vienen de
geles teñidos con agentes fluorescentes. 3.4. Digestión enzimática en gel automatizada
La automatización del proceso de digestión de proteínas en gel, así como la deposición de los digeridos resultantes en portamuestras apropiados para los espectrómetros de masas permite el procesamiento rápido de un elevado número de muestras y reduce notablemente la contaminación de las mismas con queratinas. 3.5. Identificación de proteínas por “huella peptídica de masa” mediante MALDI
Realizada la digestión enzimática, la determinación precisa de la masa molecular de los péptidos resultantes mediante espectrometría de masas MALDI genera la huella dactilar peptídica de la proteína. Para identificar dicha proteína esta huella se compara con las masas predichas por la digestión in silico de todas las secuencias incluidas en las bases de datos de proteínas.
3.6. Identificación y caracterización de proteínas por “huella peptídica de fragmentación” mediante MALDI
El módulo LIFT del espectrómetro de
masas MALDI-TOF/TOF ofrece la posibilidad de medir con gran eficacia los espectros de fragmentación de algunos de los

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
56
péptidos generados por la digestión enzimática. Esta huella peptídica de fragmentación puede emplearse en combinación con la huella peptídica de masa para aumentar notablemente la eficacia de la búsqueda en bases de datos, permitiendo incluso la identificación de proteínas de organismos cuyo genoma es aún desconocido. En ciertos casos, estos espectros de fragmentación permiten determinar sitios de modificaciones post-traduccionales u obtener secuencias parciales mediante secuenciación de novo. 3.7. Identificación de proteínas por nLC-MS/MS (ESI-Trampa iónica)
Esta configuración de cromatografía líquida acoplada a trampa iónica se emplea para la identificación de mezclas poco complejas de proteínas mediante el estudio de sus espectros de fragmentación. El resultado es una lista de masas de péptidos (huella peptídica de masa) y de sus correspondientes fragmentos (huella peptídica de fragmentación), de modo que la eficacia de la búsqueda en bases de datos es muy elevada. Los espectros MS/MS obtenidos permiten determinar la secuencia peptídica de novo y la localización de sitios de modificación post-traduccional. 3.8. Secuenciación de péptidos mediante nanoSpray off-line
Este abordaje contempla el análisis directo de una mezcla de péptidos en disolución, en ausencia de contaminantes, para su ionización y fragmentación en una fuente ESI. Ofrece la ventaja de poder efectuar rondas sucesivas de selección y fragmentación, MS(n), de modo que mediante la correcta interpretación de los espectros de fragmentación se puede llegar a determinar la secuencia peptídica de novo y la localización de sitios de modificación post-traduccional. 3.9. Identificación de proteínas en mezclas complejas por cromatografía
bidimensional acoplada on-line a ESI-Trampa iónica
En esta configuración, la totalidad de los péptidos trípticos procedentes de la digestión de una mezcla muy compleja de proteínas (p. ej., un extracto celular completo), son separados en función de su afinidad por diferentes fases cromatográficas colocadas en línea y fragmentados en tiempo real en el analizador de trampa iónica. Este abordaje, que resulta muy sensible, es apropiado cuando se pretende soslayar las limitaciones de la electroforesis bidimensional y para el análisis de ciertos subproteomas. 3.10. Identificación de modificaciones post-traduccionales en proteínas
Uno de los aspectos más importantes de la proteómica es la caracterización de modificaciones post-traduccionales (PTMs), responsables de efectos fisiológicos de enorme relevancia así como de la regulación fina de un gran número de procesos biológicos. Son numerosas la PTMs importantes descritas en la literatura (glicosilación, nitrosilación, acetilación, fosforilación, ubiquitinación, hidroxilación, etc.). El correcto análisis de los espectros de fragmentación inducida de los péptidos que contienen la modificación conduce a la determinación y localización exactas del residuo modificado. La Unidad dispone de un espectrómetro de masas híbrido (triple cuadrupolo-trampa iónica lineal), que ofrece notables ventajas para la determinación de ciertas modificaciones post-traduccionales como, por ejemplo, la fosforilación. Además, el filtrado selectivo de iones exclusivo del triple cuadrupolo posibilita el estudio de otras

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
57
modificaciones post-traduccionales de gran interés (p. ej., palmitoilación). 3.11. Cuantificación y análisis de la expresión de proteínas mediante el marcaje isotópico (ICAT) o isobárico (iTRAQ) diferencial
Mediante el marcaje diferencial estable se pueden etiquetar distintas mezclas de péptidos para poder obtener información cualitativa y además cuantificar relativamente las señales correspondientes a diferentes muestras. El marcaje isotópico (ICAT) se emplea para el análisis comparativo de dos estados distintos, centrándose en el estudio de los péptidos que contienen cisteínas. Por el contrario, mediante el marcaje isobárico (iTRAQ), podemos analizar y cuantificar señales correspondientes a cuatro estados diferentes; además, el análisis es extensible a todos los péptidos, ya que el marcaje se produce en el extremo amino después del proceso de digestión. Para hacer el análisis comparativo mediante iTRAQ es necesario disponer de un sistema capaz de estudiar el rango de bajo peso molecular (114-117 uma), como nuestro espectrómetro híbrido de triple cuadrupolo. 3.12. Proteómica de interacción proteína-ligando (chemical proteomics)
Una de las cuestiones novedosas que está abordando la Unidad es el desarrollo y
aplicación de métodos para el estudio de la interacción entre proteínas y sus ligandos. Estos métodos permiten determinar los sitios y el mecanismo de unión a proteínas, proporcionando una mejor comprensión del papel biológico de dichas proteínas. 3.13. Otros servicios y colaboraciones
Es posible afrontar otros problemas biológicos no contemplados en este cuadro general de servicios. La interacción continuada de los laboratorios implicados resulta fundamental para el desarrollo de protocolos que permitan alcanzar los objetivos del proyecto. 3.14. Cursos y entrenamiento en las técnicas de la proteómica
Con el objeto de formar personal cualificado capaz de sacar el máximo partido al equipamiento proteómico y de dar a conocer a los investigadores el alcance de dichas técnicas, la Unidad organiza periódicamente cursos que abarcan aspectos tanto teóricos como prácticos de la proteómica y sus técnicas asociadas. Así mismo, en colaboración con algunas casas comerciales organizamos cursos de entrenamiento en aspectos específicos de las distintas tecnologías proteómicas.
4. PUBLICACIONES 2005
1. Pucciarelli, M. G., Calvo, E., Sabet, C., Bierne, H., Cossart, P. and García-del Portillo, F. 2005. Identification of substrates of the Listeria monocytogenes sortases A and B by a non-gel proteomic analysis. Proteomics, In Press.
2. Calvo E., Escors D., López J. A., González J. M., Álvarez A., Arza E. and Enjuanes L. 2005. Phosphorylation and subcellular localisation of
Transmissible gastroenteritis coronavirus nucleocapsid protein in infected cells. J. Gen. Virol. 86: 2255-2267.
3. Calvo, E., Flores-Romero, P., López, J.A. and Navas, A. 2005. Identification of proteins expressing differences among isolates of Meloidogyne spp. (Nematoda: Meloidogynidae) by nano-liquid chromatography coupled to ion-trap mass spectrometry. J. Proteom. Res., 4:1017-1021.
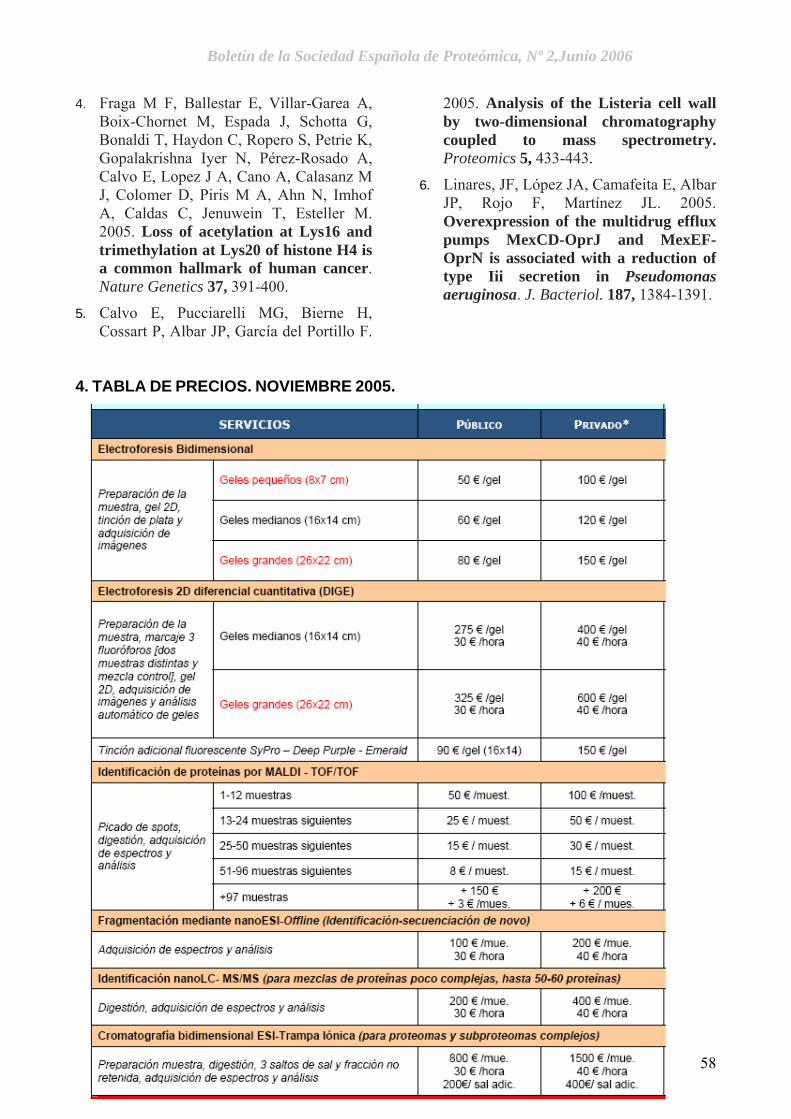
Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
58
4. Fraga M F, Ballestar E, Villar-Garea A, Boix-Chornet M, Espada J, Schotta G, Bonaldi T, Haydon C, Ropero S, Petrie K, Gopalakrishna Iyer N, Pérez-Rosado A, Calvo E, Lopez J A, Cano A, Calasanz M J, Colomer D, Piris M A, Ahn N, Imhof A, Caldas C, Jenuwein T, Esteller M. 2005. Loss of acetylation at Lys16 and trimethylation at Lys20 of histone H4 is a common hallmark of human cancer. Nature Genetics 37, 391-400.
5. Calvo E, Pucciarelli MG, Bierne H, Cossart P, Albar JP, García del Portillo F.
2005. Analysis of the Listeria cell wall by two-dimensional chromatography coupled to mass spectrometry. Proteomics 5, 433-443.
6. Linares, JF, López JA, Camafeita E, Albar JP, Rojo F, Martínez JL. 2005. Overexpression of the multidrug efflux pumps MexCD-OprJ and MexEF-OprN is associated with a reduction of type Iii secretion in Pseudomonas aeruginosa. J. Bacteriol. 187, 1384-1391.
4. TABLA DE PRECIOS. NOVIEMBRE 2005.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
59
6. PERSONAS DE CONTACTO
JEFE UNIDAD
Juan Antonio López del Olmo 91 – 453 12 00 Ext. 1200 [email protected]
Electroforesis 2D / DIGE
Juan Antonio López del Olmo 91 – 453 12 00 Ext. 1200 [email protected]
Espectrometría MALDI-TOF/TOF
Emilio Camafeita 91 – 453 12 00 Ext. 1201 [email protected]
Espectrometría ESI-Trampa / Triple Cuadrupolo
Enrique Calvo 91 – 453 12 00 Ext. 1202 [email protected]
Dirección Postal
Unidad de Proteómica Fundación CNIC Melchor Fernández Almagro, 3 28029 - Madrid Teléfono: 91 – 453 12 00 Fax: 91 – 453 12 45 http://www.cnic.es/proteomica

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
60
1. HISTORIA
La Unidad de Proteómica de la
Universidad Complutense -Parque Científico de Madrid, ubicada en la Facultad de Farmacia de dicha Universidad, se creó, EN EL 2001, como una ampliación del Centro de Asistencia a la Investigación de Secuenciación de DNA, dando origen al Centro de Genómica y Proteómica. Así se consiguió consolidar la primera Unidad de Proteómica integrada de los Centros de Apoyo a la Investigación de una Universidad española. Este Centro ha contado desde el principio con un importante apoyo de la Cátedra de Merck-Sharp and Dohme, cuyo director es el Prof. César Nombela., y cuya misión es la de promocionar la docencia y la investigación en Genómica y Proteómica en la UCM.
Durante el año 2002, los
servicios que desarrolla la Unidad de Proteómica de la UCM quedaron integrados dentro del Parque Científico de Madrid. (http://www.fpcm.es/). En el año 2005 hemos entrado a formar parte de ProteoRed, una red nacional de servicios de Proteómica, financiada por
Genoma España, dentro del nodo 1, localizado en Madrid. (http://sci.cnb.uam.es/proteored/pages/index.html).
El objetivo principal de la
Unidad de Proteómica es el de fomentar la investigación científica y el desarrollo tecnológico en la Comunidad de Madrid en el área de la Proteómica y poner a disposición de los investigadores que lo requieran un conjunto de tecnologías y equipamiento asistidos por personal cualificado. 2. EQUIPAMIENTO
2.1. Preparación de muestras
• Digestor Multiple probe 215
liquid handler de GE. • Robot para la excisión de
manchas proteicas de GE • Sistema de microdisección
mediante captura por láser. 2.2. Separación de proteínas
• Sistema de electroforesis
monodimensional de Biorad
UNIDAD DE PROTEÓMICA DE LA UNIVERSIDAD COMPLUTENSE DE MADRID- PARQUE CIENTÍFICO DE
MADRID Mª Luisa Hernáez y Concha Gil
Unidad de Proteómica, Departamento de Microbiología, Facultad de Farmacia, Plaza de Ramón y Cajal S/N, 28040
Madrid

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
61
• Electroforesis Bidimensionale: Ettan Dalt-six e IPGphor II de GE.
2.3. Análisis de imagen
• Escáner de fluorescencia
Typhoon 9400 de GE. • Image master 2D-Platinum • Software de análisis diferencial
DECYDER
2.4. Separación de proteínas
• Sistema de cromatografía
líquida capilar CapLC acoplado on-line al espectrómetro de masas Q-TOFde Waters.
• Sistema de cromatografía líqida integrado por HPLC nano Ultimate, inyector automático FAMOS, unidad para cambio automático de microcolumnas , denominado Switchos II y microcolector de fracciones PROBOT de LC-Packings.
2.5. Espectrometría de masas
• Espectrómetro de masas MALDI-TOF modelo Voyager DE-STR de Applied Biosystems.
2.6. Espectrómetros de masas en tandem
• Espectrómetro de masas Q-
TOF2 de Micromass.
• Espectrómetro de masas MALDI-TOF/TOF 4700 de Applied Byosistems.
2.7. Herramientas bioinformáticas
• Global Protein Server • ProteinLynx
2.8. Interacciones biomoleculares
• Biacore3000 3. SERVICIOS OFERTADOS
Determinación de masas moleculares.
Identificación de proteínas pos huella peptídica
Identificación de proteínas por huella peptídica y fragmentación
Secuenciación de péptidos Electroforésis monodimensional Electroforésis bidimensional Análisis de imagen Proteómica de expresión
diferencial: tecnología DIGE
4. PRÓXIMOS SERVICIOS
Cromatografía multidimensional Microdisección mediante
captura con láser Interacciones biomoleculares Proteómica de expresión
diferencial o Cuantificación con
marcaje isotópico iTRAQ

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
62
5. CURSOS IMPARTIDOS EN EL CENTRO
• Proteómica: Electroforesis bidimensional y análisis de proteínas por espectrometría de masas. Organización: Departamento de Microbiología II (Facultad de Farmacia - UCM), Unidad de Proteómica, Cátedra de Genómica y Proteómica y Aula Científica. Ediciones 2005,2004, 2003.
• Curso de Genómica y Proteómica (100 horas) Escuela Complutense de Verano Ediciones 2004, 2003, 2002.
• Curso de Proteómica (100 horas).Escuela Complutense de Verano Ediciones 2005
• Participación en cursos de Doctorado
6. USUARIOS
El Centro de Proteómica da servicio actualmente a aproximadamente 75 grupos de Investigación, principalmente pertenecientes a la Comunidad Autónoma de Madrid, tanto centros de investigación públicos como privados.
Tecnologías de identificación y cuantificación de expresión diferencial
DIGE iTRAQ
Extranjero 8% (6)
Resto de España 20% (15)
Madrid 72% (55)

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
63
Distribución por situación geográfica de usuarios de los servicios ofertados por la Unidad de Proteómica UCM-PCM 7. PERSONAL DEL CENTRO
El personal de la Unidad de Proteómica tiene dos objetivos fundamentales: dar servicio de proteómica a la comunidad científica y desarrollar y poner a punto nuevas metodologías que se están desarrollando en dicho campo.
Responsable de la Unidad de Proteómica UCM-PCM
o Concha Gil García e-mail: [email protected]
Coordinador de Proteómica del Parque Científico de Madrid
o Fernando Vivanco e-mail: [email protected]
Responsable Técnico o María Luisa Hernáez Sánchez e-
mail: [email protected] Personal Técnico
o Lola Gutiérrez Blázquez e-mail: [email protected]
o Montserrat Martínez Gomariz e-mail: [email protected]
Personal Técnico de ProteoRed o Pilar Ximénez de Embún
Cadarso e-mail:[email protected]
o Antonio Serna Sanz e-mail: [email protected]
Beca FINNOVA o Maria del Carmen Ruiz
Unidad de Proteómica e-mail: [email protected]

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
64
http://www.ucm.es/info/gyp/proteomica/
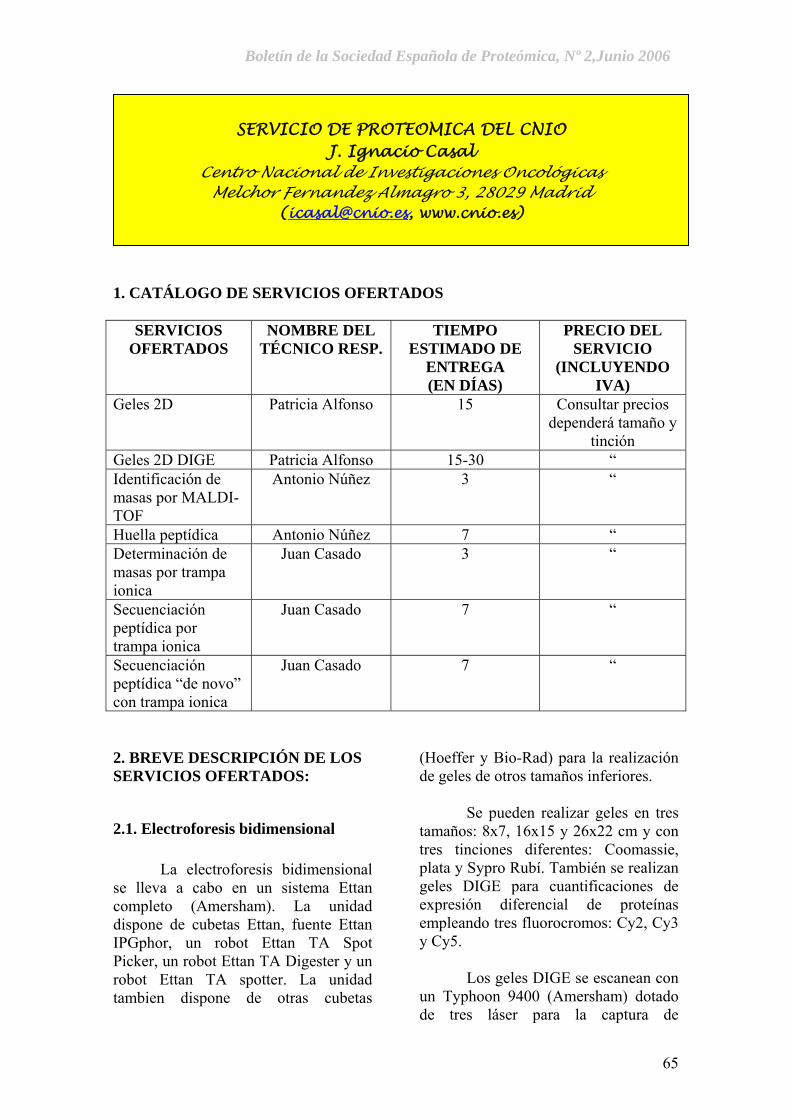
Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
65
1. CATÁLOGO DE SERVICIOS OFERTADOS
SERVICIOS OFERTADOS
NOMBRE DEL TÉCNICO RESP.
TIEMPO ESTIMADO DE
ENTREGA (EN DÍAS)
PRECIO DEL SERVICIO
(INCLUYENDO IVA)
Geles 2D Patricia Alfonso 15 Consultar precios dependerá tamaño y
tinción Geles 2D DIGE Patricia Alfonso 15-30 “ Identificación de masas por MALDI-TOF
Antonio Núñez 3 “
Huella peptídica Antonio Núñez 7 “ Determinación de masas por trampa ionica
Juan Casado 3 “
Secuenciación peptídica por trampa ionica
Juan Casado 7 “
Secuenciación peptídica “de novo” con trampa ionica
Juan Casado 7 “
2. BREVE DESCRIPCIÓN DE LOS SERVICIOS OFERTADOS:
2.1. Electroforesis bidimensional
La electroforesis bidimensional se lleva a cabo en un sistema Ettan completo (Amersham). La unidad dispone de cubetas Ettan, fuente Ettan IPGphor, un robot Ettan TA Spot Picker, un robot Ettan TA Digester y un robot Ettan TA spotter. La unidad tambien dispone de otras cubetas
(Hoeffer y Bio-Rad) para la realización de geles de otros tamaños inferiores.
Se pueden realizar geles en tres tamaños: 8x7, 16x15 y 26x22 cm y con tres tinciones diferentes: Coomassie, plata y Sypro Rubí. También se realizan geles DIGE para cuantificaciones de expresión diferencial de proteínas empleando tres fluorocromos: Cy2, Cy3 y Cy5.
Los geles DIGE se escanean con un Typhoon 9400 (Amersham) dotado de tres láser para la captura de
SERVICIO DE PROTEOMICA DEL CNIO J. Ignacio Casal
Centro Nacional de Investigaciones Oncológicas Melchor Fernandez Almagro 3, 28029 Madrid
([email protected], www.cnio.es)

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
66
imágenes. Las imágenes se analizan con el software DeCyder (Amersham). El recorte de spots se lleva a cabo con el Ettan TA Spot Picker y la digestión tríptica automática con el Ettan TA Digester.
2.2. Identificación de masas por MALDI-TOF
La unidad dispone de un MALDI-TOF-TOF 4800 (Applied Biosystems) para el análisis de masas y de identificación de proteínas por huella peptidica.
2.3. Identificación de proteínas mediante huella peptídica
La unidad realiza un servicio de identificación de proteínas por huella peptídica siguiendo la digestión tríptica de los spots seleccionados en los geles 2D o 1D. Se puede realizar huella peptídica a partir de cualquiera de las tinciones que se ofrecen, Coomassie, plata o Sypro. A partir de la huella se realiza una búsqueda on-line con los algoritmos MASCOT y/o ProFound en las bases de datos SwissProt o NCBI y se seleccionan los targets que superan el score adecuado y cubres un porcentaje mínimo de proteína del 30%. Toda esta información se envía al usuario por correo electrónico en un formato definido. En breve se pondrá a punto un servicio de LC MALDI para identificación de proteínas en muestras complejas. 2.4. Determinación de masa exacta por trampa iónica
Bajo condiciones desnaturalizantes, las proteínas producen una serie de especies correspondientes a diferentes estados de carga. La deconvolución de todos los
picos pertenecientes a las mismas moléculas conduce al cálculo preciso de la masa molecular. 2.5. Identificación de proteínas por secuenciación peptídica utilizando LC/MS-MS (trampa iónica)
Tras digestión de la proteína con una proteasa apropiada (p.e. tripsina), se puede hacer una carrera cromatográfica en una columna de fase reversa C18 en condiciones nano o micro-HPLC, conectada on-line a una fuente micro o nano-ESI y al espectrómetro de masas de trampa iónica. Por fragmentación por colisión de los péptidos y análisis de las series de iones resultantes se obtiene la secuencia del péptido correspondiente mediante interrogación utilizando el algoritmo SEQUEST en bases de datos NCBI o SwissProt.
En el año 2004 se adquirió un equipo de espectrometría de masas por trampa iónica con fuente de ionización por electrospray y acoplado a un sistema de cromatografía líquida con nano-HPLC. Concretamente se adquirió el equipo de trampa iónica lineal LTQ de alta precisión acoplado a un nano-HPLC Surveyor equipado con inyector automático de muestra, todo ello de la marca Thermo Finnigan. Este equipo es capaz de suministrar espectros MS/MS y aplicaciones LC/MS/..../MSn. Asimismo, lleva incorporado un paquete de software Bioworks 3.1 que incorpora la última versión de software TurboSEQUEST, para identificación rápida de proteínas en bases de datos indexadas, incluye web server profesional, O´Reilly y software de cuantificación ICAT y el programa DeNovox para la secuenciación manual de péptidos de origen desconocido. Para mejorar la sensibilidad del instrumento se ha adquirido recientemente (2005) un nano-HPLC Ultimate 3000 (LC-Packings) dotado de la capacidad de realizar cromatografía

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
67
multidimensional, para la realización de la técnica MudPIT.
2.6. Secuenciación peptídica “de novo”
Similar al método anterior pero aplicado a proteínas obtenidas de
organismos cuya base de datos genómica no ha sido completada, es inexistente, existen datos conflictivos o mutaciones no descritas. En este caso se puede deducir la secuencia aplicando un software adecuado (p.e “DeNovox”).

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
68
1. INFORMACIÓN GENERAL Responsable: David Andreu Personal: Cristina Chiva: técnica superior especialista Eva Borrás: técnica superior especialista Mireia Segura: técnica superior especialista Manuel Rodríguez: técnico especialista Página web: www.upf.es/cexs/sct
Contacto: e-mail: [email protected]
Teléfono: 93 5422825
Fax: 93 5422802
2. SERVICIOS OFERTADOS
- Separaciones de proteínas por electroforesis 2D (geles de 7 y 24 cm).
- Análisis de imagen de geles 2D utilizando tinción con plata y Coomassie coloidal.
- Digestión, tanto automática como manual, de bandas de gel específicas de proteínas y posterior identificación de éstas por MALDI-TOF y búsqueda en bases de datos.
- Identificación de proteínas por digestión, separación por nanoLC y análisis MS/MS en nanoES-Q-TOF.
- Secuenciación de Edman de proteínas y péptidos.
- Síntesis de péptidos (escala 1-100 mg). Marcaje en posiciones específicas (biotina, fluoróforos, etc), residuos modificados (fosforilación, sulfatación, lipidación) y conjugación a proteínas.
3. INSTRUMENTACIÓN
3.1. Electroforesis bidimensional de proteínas
El servicio dispone de un sistema de electroforesis 2D para la separación de proteínas que incluye:
- Ettan IPGphor Isoelectric Focusing, Amersham Biosciences. Permite separar las proteínas según el punto
SERVICIO CIENTÍFICO-TÉCNICO DE PROTEÓMICA UNIVERSIDAD POMPEU FABRA/PARC DE RECERCA
BIOMÈDICA DE BARCELONA David Andreu
Departamento de Ciencias Experimentales y de la Salud. Unidad de Proteómica y Química de Proteínas. Universidad
Pompeu Fabra. Dr. Aiguader 80, 08003 Barcelona

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
69
isoeléctrico (pI) en tiras de diferentes gradientes de pH inmovilizados, en la primera dimensión. Puede procesar hasta a 12 tiras, de 7 ó 24 cm, simultáneamente.
- Ettan Dalt II, Amersham Biosciences. Es un sistema de electroforesis vertical en que las proteínas se separan de acuerdo con la masa molecular, en la segunda dimensión. Se pueden hacer, simultáneamente, hasta 12 geles de 26x20 cm .
- Mini-PROTEAN 3, Bio-Rad, para la realización de la segunda dimensión en geles de 8x7 cm.
- Accesorios para la preparación, fijación, tinción y conservación de geles.
- ImageScanner, Amersham Biosciences, para la detección de imagenes de geles.
- ImageMaster gel analysis software, Amersham Biosciences, para el procesado de resultados de experimentos de electroforesis 2D.
3.2. Digestión de bandas
Las bandas consideradas de interés se recortan de los geles 2D y se procede a la digestión proteolítica (in-gel digest) de la proteína, como etapa previa a su análisis e identificación por espectrometría de masas. La digestión, extracción de péptidos resultantes y
desalado de los mismos, puede realizarse individualizadamente, o de modo más habitual en dispositivos de procesado simultáneo tales como el kit de digestión Montáge (Millipore) o la estación robotizada Multiprobe (Perkin Elmer)
3.3. Espectrómetro de masas MALDI-TOF mod. Voyager STR, Applied Biosystems
El espectrómetro de masas
MALDI-TOF está equipado con fuente

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
70
MALDI que utiliza un láser de nitrógeno que opera a 337 nm y un detector de tiempo de vuelo. Permite la determinación de masa molecular de péptidos y proteínas con elevada exactitud (0.001% en modo reflectrón con calibrado interno), resolución (>10 000 de modo rutinario). Este equipo además es muy sensible, y puede detectar niveles de fmol de péptido y también permite la determinación de
pesos moleculares de proteínas, y biopolímeros en general, con excelente precisión y sensibilidad. También permite la identificación de proteínas (separadas en geles mono o bidimensionales) mediante digestión tríptica. La identificación se realiza mediante las pertinentes herramientas informáticas de búsqueda en base de datos (Mascot) disponible en modo residente en el servicio.
3.4. Espectrómetro de masas mod. Q-Star Pulsar MDS Sciex/Applied Biosystems
El espectrómetro de masas mod. Q-Star Pulsar está equipado con diversos dispositivos de introducción de muestra (ionspray, nanospray, o-MALDI); analizador de masa compuesto por cuadrupolo, cámara de colisión y tubo de vuelo (TOF). Adquisición de espectros en modo normal (MS) o tándem (MS/MS), que posibilita la secuenciación. Dispone también de un sistema nano-LC mod. Ultimate (LC Packing), con inyector automático, conectado a la fuente iónica, lo cual permite el análisis de
mezclas complejas de péptidos (p. ej., procedentes de digestión).
La principal utilización de los recursos anteriores es la identificación de proteínas (separadas en geles mono o bidimensionales) mediante digestión tríptica y análisis MALDI-TOF del digerido. Alternativamente, la identificación puede realizarse –con mayor fiabilidad- por MS/MS de alguno(s) de los péptidos trípticos. La identificación se realiza mediante las pertinentes herramientas informáticas de búsqueda en base de datos (Mascot, Protein Prospector, etc.), disponibles en modo residente en el servicio.
3.5. Secuenciador de Edman, Procise cLC 491, Applied Biosystems
Permite el análisis y la identificación de proteínas, incluidas posibles modificaciones

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
71
posttraduccionales, utilizando la química de Edman. Este sistema rompe secuencialmente y de una manera totalmente automática, los aminoácidos del extremo amino terminal de la proteína y analiza por HPLC capilar los derivados PTH-aminoácido, resultado
de la escisión. El sistema capilar de cromatografía y el especial diseño del equipo permiten analizar proteínas a nivel subpicomolar.
3.6. Sintetizador de péptidos mod. 433 Applied Biosystems
El sintetizador de péptidos 433A, está constituido por un modulo de reacción y un sistema de válvulas dispensadoras de reactivos y disolventes, el sistema permite la síntesis en fase sólida de péptidos y pequeñas proteínas en un proceso repetitivo y totalmente automatizado de acoplamiento de los aminoácidos des de el extremo C-terminal, el cual está permanentemente ancorado a un suporte polimérico. Se trata de un equipo de gran versatilidad que puede trabajar con
las dos químicas más generales (Boc y Fmoc), con cualquiera estrategia de activación y a un gran número de escalas. Permite la obtención de rutina de péptidos sintéticos de hasta 25-30 aminoácidos, en forma libre o modificados en posiciones específicas (biotinilación, lipidación, glicosilación, fosforilación, etc). Con previa consulta puede estudiarse la síntesis de secuencias de talla superior o con otras modificaciones (cíclicos, aminoácidos no naturales, etc.)
3.7. Sintetizador múltiple de péptidos SyroII (MultiSyntech)
Permite la preparación simultánea de varias (hasta 48) secuencias peptídicas, de tallas hasta

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
72
20-25 aminoácidos, en forma rápida y eficaz.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
73
1. HISTORIA
En el año 1993 se creó el Servicio de “Química de Proteínas” del CBMSO. La trayectoria del laboratorio, actualmente Servicio de “Proteómica”, ha sido paralela a la evolución de la comunidad científica en el estudio de las proteínas y tiene como objetivos:
• Ofrecer apoyo y asesoramiento
científico-técnico al usuario-investigador:
o Métodos apropiados para la preparación de la muestra.
o Diseño del flujo de trabajo para cada objetivo.
• Aplicar todas las técnicas
disponibles en el laboratorio para el análisis de proteínas.
La oferta del Servicio va dirigida a
usuarios del CBMSO, otros centros públicos y entidades privadas (los baremos aplicados varían con el tipo de trabajo y el origen del usuario). 2. PERSONAL
El personal adscrito al Servicio y sus funciones actuales son las siguientes:
• Alberto Jorge García:
Responsable de la recepción de muestras, digestión en gel, análisis MALDI-TOF e interpretación de resultados.
Laboral fijo (CSIC), Técnico de Investigación y Laboratorio.
• Yolanda Núñez:
Responsable del análisis de
muestras y la interpretación de resultados mediante LC-ESI-MS/MS.
Funcionaria (CSIC), Técnico Especialista de Grado Medio
• Dra. Mercedes del Valle:
Responsable del análisis de MPT. Titulado Superior Especializado
(ProteoRed). • Dra. Esperanza Morato: Responsable de la confección de
geles bidimensionales. Titulado Superior Especializado
(ProteoRed). • Dra. Anabel Marina:
Responsable del Servicio de
Proteómica, coordinación y supervisión de todos los análisis así como de las tareas administrativas relacionada con el Servicio.
SERVICIO DE PROTEÓMICA DEL CENTRO DE BIOLOGÍA MOLECULAR “SEVERO OCHOA”
Anabel Marina Ramírez Lab. 001, CBMSO, Edificio de Biológicas, Facultad de
Ciencias, U.A.M., Cantoblanco, 28049, Madrid

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
74
Funcionaria (CSIC). Titulado Superior Especializado. 3. EQUIPAMIENTO
- Espectrómetro de masas de trampa iónica modelo LCQ Classic de Thermo Finnigan, equipado con sistemas de ionización de nanospray y de microspray de fabricación propia acoplado a un HPLC Agilent 1100.
- Espectrómetro de masas tipo MALDI-TOF, modelo Autoflex de Bruker, equipado con reflector y sistema para PSD. - Equipamiento completo para la confección de geles bidimensionales Amhersam-Pharmacia (IPG-Phor, cubetas 2ª dimensión y fuente de alimentación).
- HPLCs (2) modelo Smart de Pharmacia, con colector automático de fracciones. - Robot digestor/preparador de muestras para MALDI-TOF Autoflex (pertenece al Servicio de Proteómica del nodo U.A.M. de la red cardiovascular). - “Softwares” para el análisis de datos.
4. TÉCNICAS-OFERTA DE ANÁLISIS
Básicamente la oferta de nuestro
laboratorio se puede agrupar en dos dependiendo de la procedencia de la muestra a analizar:
• Análisis mediante
Espectrometría de Masas.
• Identificación
sistemática de proteínas presentes en las bases de datos a partir de geles.
*Análisis mediante Espectrometría de masas
1. Análisis empleando MALDI-TOF
• De muestras preparadas por el usuario (incluyendo portamuestras y matriz).
• De muestras preparadas por el Servicio.
2. Análisis empleando LC-ESI-IT
de fracciones de péptidos provenientes de HPLC preparadas por el usuario.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
75
*Servicio de Proteómica propiamente dicho
1. Confección de geles bidimensionales.
2. Tinción fluorescente (SYPRO-
Ruby) y adquisición de imagen (Typhoon).
3. Digestión tríptica en gel SDS-
PAGE, mono ó bidimensional, preparado por el usuario.
4. Análisis mediante MALDI-TOF: Obtención del mapa peptídico e identificación mediante huella peptídica (PMF).
5. Análisis mediante LC-ESI-IT:
Basándonos en el mapa de MALDI, si es bueno pero se sospecha de mezcla de proteínas se realiza Exclusión Dinámica (DE) y búsqueda en las bases de datos (DB) mediante TurboSequest; si el mapa de MALDI es pobre ó interesan determinadas péptidos se aplica la Monitorización de iones sencillos (SIM) que, a pesar de ser una aproximación más lenta es superior en prestaciones a equipos de nueva generación como el MALDI-TOF-TOF.
5. PAGINA WEB (http://www.cbm.uam.es/servicios/proteomica.htm
SDS-PAGE
Bandas de proteínas
μ-desalado
MALDI-TOF
Secuenciación “de novo”. MPT
2D-IEF-SDS-PAGE Tinción
Proteínas Péptidos sintéticos
Péptidos MHC Digestión “in situ”
On-line LC-MS
“Mapa de péptidos”
Identificación “numérica” de
proteína/s en BD“Aislamiento y Fragmentación
ESI-trampa iónica
Datos de peso moleculary de secuencia
Off-line NanoESI

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
76
La página web del Servicio está en continuo proceso de actualización ya que pretende ser una herramienta donde el usuario pueda hacer todo tipo de gestiones y consultas relacionadas con todo lo concerniente a nuestro laboratorio.
La primera etapa del procedimiento de trabajo es:
• Recepción y control de muestras
Hemos creado una hoja excel (con acceso directo desde nuestra web) a disposición de cualquier usuario que contiene la relación de los mismos, fechas de recepción de muestras, estatus del trabajo, etc. Cualquier incidencia ó avería se hace constar en esta hoja así como la fecha de resolución de las mismas. La segunda etapa del procedimiento consiste en el:
• Análisis. • Resultados.
Los resultados se presentan en formato “power-point” y se entregan vía e-mail al usuario.
6. GRADO DE CONSECUCIÓN
DE LOS TRABAJOS • Análisis mediante
Espectrometría de Masas de muestras preparadas por el usuario empleando MALDI-TOF.
Durante el año 2004 se analizaron
2169 muestras. Dado el alto número de muestras
que se solicitan, estas se analizan en modo automático con un grado de consecución muy positivo.
• Análisis mediante
Espectrometría de Masas de muestras preparadas en el servicio empleando MALDI-TOF.
Este tipo de análisis se aplica
básicamente al control de calidad de los péptidos sintéticos que se obtienen en el Servicio de Síntesis de Péptidos y es
Consultar página web www2.cbm.uam.es/proteomica
Ubicación, e-mail, personal
Oferta de Análisis
Infraestructura
+
Entrega de RESULTADOS en formato power point
vía e-mail
Solicitud de Servicio Detalle del trabajo a
realizar
Entrega de muestras
Estatus de muestras y análisis en tiempo
Consulta “on-line”
ANÁLISIS Interpretación

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
77
crítico para determinar que la síntesis ha funcionado correctamente.
• Análisis mediante
Espectrometría de Masas LC-MS-IT de fracciones de HPLC preparadas por el usuario.
El grado de consecución es muy
positivo teniendo en cuenta la complejidad de las muestras y los problemas intrínsecos a la secuencia de los péptidos MHC.
El modo más usual de trabajo es el de alta sensibilidad (SIM) obteniéndose espectros de fragmentación con todas las muestras analizadas.
Durante el año 2004 se analizaron 111 muestras.
• Confección de geles bidimensionales
La demanda de este servicio por
nuestro centro es muy baja. • Identificación de proteínas
mediante “huella peptídica” (PMF).
Este es uno de los trabajos más
demandados por el usuario. Para muestras que se visualizan
mediante tinción de coomassie se obtuvieron espectros de masas con más del 90% de las bandas/spots analizados. Los motivos del alto porcentaje de éxito son principalmente dos:
1. La cantidad de proteína
es suficiente (el límite de sensibilidad del coomassie es de 0,1 µg).
2. La tinción es compatible con el proceso de digestión y análisis posterior MS.
Durante el año 2004 se analizaron 184 muestras
Si la muestra procede de un gel teñido con plata los resultados son totalmente impredecibles por los siguientes motivos:
1. La plata es un agente oxidante
muy fuerte que modifica todas las proteínas en distinto grado haciéndolas resistentes a digestión.
2. La cantidad de proteína es baja y
no se puede cuantificar (problema intrínseco a la tinción) por lo que la intensidad no se correlaciona con la cantidad y es variable gel a gel.
Por este motivo el Servicio ha
invertido mucho tiempo y esfuerzo empleando todos los protocolos de digestión y tinción “compatibles” disponibles en la bibliografía. A pesar de ello el porcentaje de éxito fue muy bajo por lo que desde el pasado mes de mayo del 2004 implementamos un nuevo método de tinción fluorescente empleando SYPRO Ruby. La sensibilidad de este fluoróforo es igual o superior a la de la plata “compatible” obteniéndose unos resultados excelentes por:
1. Su total compatibilidad con los
métodos de digestión y análisis MS.
2. 3. Su excelente rango dinámico:
tres órdenes de magnitud. De los spots/bandas digeridos hasta
la fecha hemos obtenido espectros de masas MALDI-TOF en un 80% de las muestras, teniendo en cuenta que esta tinción se emplea cuando se trabaja a alta sensibilidad, por lo que la cantidad de proteína es baja (femtomoles).

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
78
● Identificación de proteínas a partir de espectros MS/MS de péptidos mediante LC-MS/MS ESI-IT.
Durante el año 2004 se analizaron 32 muestras.
Cuando una banda/spot proteíco
no se identifica mediante la técnica anteriormente descrita, PMF, a pesar de haberse obtenido un buen espectro MS, los motivos más comunes son los siguientes:
1. Mezcla de proteínas. 2. Proteína no presente en
bases de datos.
En el primero de los casos, siempre y cuando las proteínas contenidas en la mezcla estén presentes en las bases de datos, el grado de consecución de resultados es muy bueno. Identificándose así qué proteínas componen la mezcla.
En el segundo de los casos el grado de consecución de resultados depende del grado de homología que tenga la proteína con otras proteínas presentes en las bases de datos. Si este grado es medio-alto se identifica sin problemas mediante identidad de
secuencias cortas (péptidos), si es medio-bajo (caso de organismos como plantas, hongos…) el porcentaje de identificaciones mediante homología es bajo.
En cualquiera de los casos SIEMPRE se obtienen espectros de fragmentación MS/MS de buena calidad. ● Caracterización de péptidos a partir de espectros MS/MS mediante LC-ESI-IT en modo SIM.
En numerosas ocasiones el usuario tiene interés en caracterizar un péptido en particular de una proteína. En ese caso se emplea el modo de trabajo a alta sensibilidad que monitoriza el/los péptidos de interés. La sensibilidad de este método es extrema y siempre se obtienen espectros de fragmentación, como mínimo, de uno de los cuatro péptidos que se monitorizan (máximo permitido por la velocidad de barrido del analizador).
De este modo hemos
identificado muchos péptidos (a partir de los cuales se identifica la proteína) no detectados mediante espectrometría de masas MALDI-TOF por su baja proporción relativa en la mezcla inicial.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
79
PLATAFORMA DE PROTEÓMICA, PARC CIENTIFIC DE BARCELONA
Eliandre de Oliveira, David Bellido, Antonia Odena, Stéphanie Boussert
Parc Científic de Barcelona, C/ Josep Samitier 1-5, 08028 - Barcelona
1. PERSONAL
Eliandre de Oliveira Responsable del laboratorio, codirectora de tesis de una estudiante de doctorado. (Contrato Torres-Quevedo).
David Bellido Español Técnico Superior. Involucrado en el desarrollo de métodos en proteómica, responsable de la electroforesis 2D y del ESI-Q-TOF.
Antonia Odena Técnica Superior. Involucrada en el desarrollo de métodos en proteómica, responsable del MALDI-TOF-TOF y de la secuenciación de Edman. (Contrato Proteored).
Stéphanie Boussert Estudiante de doctorado. Su proyecto de tesis se basa en el desarrollo de superficies de afinidad para aplicaciones en espectrometría de masas.
2. HISTORIA
El Parc Científic de Barcelona es una estructura creada por la Universitat de Barcelona que incorpora grupos de investigación públicos y empresas en un único espacio equipado con una amplia oferta tecnológica.
Ubicado en el campus Diagonal,
acoge veinte empresas, tres centros de investigación y la Bioincubadora CIDEM-PCB en un edificio modular de 20.000 m2 equipado con potentes plataformas para la I+D+I entre las que se incluye la Plataforma mixta de Proteómica. Esta plataforma ha sido creada con el objetivo de potenciar el campo de la proteómica.
La Plataforma de Proteómica ofrece tecnología e infraestructura completa y de última generación para estudios proteómicos . El servicio se ofrece des del año 2002 tanto a la comunidad científica como a las empresas privadas. 3. TECNOLOGÍA Y SERVICIOS
Las técnicas disponibles permiten hacer estudios de expresión proteica electroforesis bidimensional y/o cromatografía líquida bidimensional acoplada on-line y/o off-line a espectrometría de masas.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
80
Identificación con alto redimiendo de proteínas a través del proceso de digestión enzimática y análisis de huella peptídica (PMF) y secuenciación de péptidos por MALDI-TOF/TOF (PMF + MS/MS) y/o capLC-Q-TOF. La plataforma dispone de un colector de micro fracciones que permite la separación de los péptidos provenientes de las digestiones enzimáticas, que pueden ser directamente fraccionados y colectados en una placa de MALDI, para ser analizados en el MALDI-TOF/TOF. Esto aporta la ventaja de poder analizar muestras más complejas de forma más rápida, ya que los péptidos provenientes de la digestión enzimática podrán ser separados por cromatografía líquida en una o en dos dimensiones (por fase reversa, o cromatografía de intercambio iónico seguido de fase reversa) y posteriormente analizados en el MALDI-TOF/TOF (capLC-Probot-MALDI-TOF/TOF).
Se utiliza secuenciación de novo MSMS de péptidos provenientes de digestión proteica para identificar secuencias no descritas por homología de secuencia.
La plataforma también dispone de un secuenciador de proteínas por degradación de Edman de alta sensibilidad. Es de gran utilidad en la identificación de proteínas que necesitan una secuenciación más exacta, como puede ser el caso de las proteínas que sufren modificaciones postraduccionales (PTM) en la región N-terminal, o en péptidos que no son identificados por espectrometría de masas.
Localización de PTMs como fosforilaciones y acetilaciones.
Además, en los laboratorios del PCB se dispone de salas de cultivos
celulares, microscopios, sala de espectroscopia, congeladores, cámaras frías, ultra-centrífugas, sala de experimentación animal, etc. Cabe destacar la presencia de un servicio de síntesis de péptidos ubicado junto a la plataforma de proteómica. Este servicio puede ser útil si se aplica, juntamente con los ICATs la tecnología AQUA en los estudios proteómicos cuantitativos.
Distribución de los servicios en 2004
Interno57%
Público15% Privado
28%
4. I+D
La Plataforma de Proteómica realiza labor de investigación y desarrollo en colaboración con diferentes grupos de investigación. Algunos de los proyectos son: Extracción e identificación de proteínas de cerebro humano relacionadas con la enfermedad de Alzheimer por medio de sistemas afinidad específica (superficies de oro químicamente modificadas y funcionalizadas con proteína �-amiloide) y identificación de proteínas de cerebro humano relacionadas con esquizofrenia (por medio de afinidad con metales). Las proteínas son digeridas enzimaticamente y los péptidos obtenidos analizados por nanoLC-nanoESI-MSMS y/o MALDI-MSMS.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
81
Caracterización de subproteomas del tomate Expresión diferencial de proteínas en hígados expuestos a bebidas alcohólicas.) Identificación del proteoma de liquido cerebro-espinal de fetos de Gallus gallus. Identificación de expresión diferencial de proteínas en bacterias resistentes a antibióticos. Identificación de proteínas fosforiladas en Arabidopsis. 5. EQUIPOS Electroforesis IPGphor de Amersham Biosciences Ettan Dalt II de Amersham Biosciences Gel Caster Six de Amersham Biosciences Sistema Hoefer 250/260 Migthy Small II de Amersham Biosciences Análisis de imágenes Scanner para digitalizar geles ImageScanner de Amersham Biosciences Softwares ImageMaster 2D y ImageMaster Database de Amersham Biosciences Escisión y digestión de spots Spot Picker de Amersham Biosciences Digestor automático Progest de PerkinElmer Espectrometría de masas MALDI-TOF/TOF 4700 Proteomics Analyzer de Applied Biosystems HPLC para nanoflujo capLC de Waters ESI-Q-TOF Q-Tof Global de Micromass Colector de microfracciones/placa de MALDI – Robot Probot de LC Packings
Análisis de datos Estación de trabajo para análisis de datos de forma automática con GPS de Applied Biosystems o con Global Server de Waters-Micromass. Licencias de softwares para secuenciación de novo: PepSeq de Micromass y DeNovo tool de Applied Biosystems. Licencias de softwares ProteinProspector y Mascot para búsquedas en bases de datos. Secuenciador de Edman Procise cLC 492 de Applied Biosystems.
6. PUBLICACIONES Santos, C.; Mateus, M.L.; dos Santos, A.P.; Moreira, R.; de Oliveira, E. and Gomes, P. (2005) Cyclization-activated prodrugs. Synthesis, reactivity and toxicity of dipeptide esters of paracetamol. Bioorganic & Medicinal Chemistry Letters, 15: 1595–1598 Tarragó, T.; Sabidó, E.; Kogan, M.J.; de Oliviera, E. and Giralt, E. (2005) Primary structure, recombinant expression and homology modelling of human brain prolyl oligopeptidase, an important therapeutic target in the treatment of neuropsychiatric diseases. Journal of Peptide Science, 11:283-287 Martí, S.; Sánchez-Cespedes, J.; Oliveira, E.; Bellido, D.; Giralt, E. and Vila, J. (In Press) Proteomic analysis of a fraction enriched in cell envelope proteins of Acinetobacter baumannii. Journal of Proteomics Irar, S.; de Oliveira, E.; Pagès, M. and Goday, A. (In Press) Towards the identification of LEA phosphoproteoma

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
82
in Arabidopsis by two-dimensional gel electrophoresis and mass spectrometry. Journal of Proteomics Cruz, L.J.; Insua, M.M.; Pérez Baz, J.; Trujillo, M.; Rodriguez-Mias, R.A.; Oliveira, E.; Giralt, E.; Albericio, F. and Cañedo, L.M. (In Press) IB-01212, A
NEW CYTOTOXIC CYCLODEPSIPEPTIDE ISOLATED FROM THE MARINE FUNGUS Chlonostachys.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
83
UNIDAD DE PROTEÓMICA, GENÓMICA Y BIOINFORMÁTICA DEL CENTRO DE INVESTIGACIÓN MÉDICA APLICADA
Fernando J. Corrales Unidad de Proteómica. Fundación para la Investigación
Médica Aplicada (FIMA). Pío XII, 55. 31008 Pamplona
La Unidad de Proteómica Genómica y Bioinformática del Centro de Investigación Médica Aplicada (CIMA) y Universidad de Navarra (UNAV), se creó en el año 2001 con el objetivo de contribuir al desarrollo de proyectos de investigación aportando nuevas soluciones experimentales en el contexto de las nuevas tecnologías. El CIMA es un centro de investigación cuya prioridad es potenciar una investigación básica de alto nivel para generar información fundamental que permita ampliar nuestro conocimiento de los mecanismos moleculares asociados al desarrollo de enfermedades en cuatro áreas de interés: hepatología, fisiopatología cardiovascular, neurociencias y oncología. Es claro que la posibilidad de analizar perfiles de expresión génica a escala global, así como el estudio de proteomas, abre nuevas fronteras en la investigación biomédica, tanto en la generación de nuevos conceptos en biología, como en la identificación de nuevas dianas diagnósticas y terapéuticas o en la disección del mecanismo de acción de fármacos. Sin embargo, el nivel de inversión necesario y el alto grado de especialización que son necesarios para desarrollar y utilizar estas técnicas nos hizo considerar que este proyecto requería la coordinación de un grupo de investigación pluridisciplinar que permitiera establecer una plataforma en base al desarrollo e integración de nuevas tecnologías y a la elaboración de herramientas que facilitasen la interpretación e intercambio de los resultados obtenidos. Con esta idea, comenzamos, con más entusiasmo que
experiencia, nuestra andadura en el área de la proteómica y de la bioinformática desde los más elementales rudimentos en estas exigentes y cambiantes disciplinas.
En lo que a la proteómica se
refiere, comenzamos nuestro trabajo de puesta a punto con un sistema de electroforesis bidimensional (2DE) clásico, un robot digestor con capacidad para procesar 192 muestras y un MALDI TOF que permitía la identificación de proteínas mediante la aproximación de la huella peptídica.
Tabla1. Infraestructura disponible en la Unidad de Proteómica del CIMA.
InstrumentaciónHPLCAKTA FPLCRotoforMódulo de isoelectroenfoque Protean (x2)IPGphorCubetas Protean II XL (x3)Cubetas miniprotean (x3)Ettan DaltsixMódulo de transferencia mini Densitómetro GS-70 TyphoonRobot digestor (192 muestras)Espectrómetro de masas MALDI TOFEspectrómetro de masas Q TOF MicroCromatógrafo líquido capilar
SoftwarePDQuest 7.1(2 licencias)DeCyderProteinLynx Global Server 2.05BioLynx NTProteinLynxMaxEnt electrospray NTMaxEnt IIIGarban II (Desarrollado en CIMA)

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
84
Nuestro primer objetivo fue poner a punto el conjunto de estrategias experimentales que nos permitieran abordar estudios de proteómica diferencial y, para ello, enfocamos nuestra investigación en la identificación de proteínas que potencialmente estuvieran implicadas en la patogénesis de enfermedades hepáticas. Esta etapa inicial de puesta en marcha se pudo culminar con éxito gracias a la inestimable labor del Dr. Manuel Sánchez del Pino, Pino, que actualmente es el responsable del laboratorio de proteómica del Centro de Investigación Príncipe Felipe (CIPF) en Valencia. A finales de 2002 adquirimos un QTOF MICRO, un capLC y un segundo sistema de electroforesis bidimensional y, con ello, dábamos por cerrada la primera fase de dotación de infraestructura, hasta 2005. En estas fechas se incorporaron a la unidad Carmentxu Miqueo, nuestra técnico de laboratorio y Carolina Gómara, actualmente en el CICBiogune, en Bilbao. Con ellas, y con el generoso esfuerzo de nuestros becarios, el ya doctor Enrique Santamaría, Jokin Fernández y Javier Muñoz, conseguimos consolidar un
procedimiento estándar para analizar de forma genérica proteomas de sistemas biológicos: 2DE e identificación de proteínas mediante MALDI TOF MS o nanoLC ESI-MS/MS. Esta fue inicialmente la base de nuestro servicio que comenzó, de forma estructurada, a principios de 2004. Poco después se incorporaba al laboratorio la Dra. Laura Sesma, cuyo conocimiento y aportación han sido esenciales en el desarrollo y coordinación del trabajo que se realiza en la unidad. Finalmente, este año se ha incorporado María I. Mora, la especialista de la plataforma DIGE que estará operativa en nuestro laboratorio en el mes de enero de 2006.
En la actualidad el equipamiento y los experimentos que se incluyen en nuestro servicio están resumidos en las tablas 1 y 2 respectivamente. Consideramos que el servicio debe restringirse a aquellas técnicas que se puedan “estandarizar” y que, por lo tanto son fácilmente tarifables. Sin embargo, realizamos un buen número de trabajos en los que parte de la tarea está claramente incluida en el servicio, pero requieren una dedicación adicional difícil de cuantificar que generalmente hace referencia al análisis de los
resultados, ajuste de los protocolos de análisis para obtener resultados óptimos, o incluso desarrollo de nuevos métodos. En este caso establecemos, en la medida de nuestras posibilidades, colaboraciones científicas con los grupos interesados. En nuestra opinión, sean cuales sean las circunstancias, sevicio o colaboración, consideramos esencial mantener un dialogo continuo con los investigadores para diseñar los proyectos y resolver los problemas que se plantean de la manera más eficaz y que mejor se ajuste a las necesidades específicas de cada usuario.
207
985
437
3970
0100200300400500600700800900
1000
Geles 2D PMF NanoLC-ESI/MS/MS
Mezclas(ipp)
Otrasmoléculas*
Enero-Octubre 2005
Figura 1. Análisis realizados en la Unidad de Proteómica del CIMA en el periodo enero-octubre de 2005

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
85
Tabla 2. Resumen de lo servicios ofrecidos en la Unidad de Proteómica del CIMA.
Además, entendemos que es esencial realizar una labor de formación y difusión a través de la organización de cursos, que asegure la correcta utilización de los servicios. En nuestro caso hemos realizado dos cursos prácticos de formación en 2005 que han facilitado enormemente la interacción de los investigadores con el servicio. Para reflejar el índice de actividad de nuestra unidad, se muestran el la figura 1 los trabajos realizados de enero a octubre de 2005. Toda la
información referida en este documento, así como la forma de contactar y solicitar servicios en nuestra unidad puede realizarse a través de la sección destinada a las unidades de apoyo en la página web de CIMA: http://www.cima.es/index_fima.html. Nuestro laboratorio forma parte del Instituto Nacional de Proteómica (ProteoRed: http://www.proteored.org/pages/index.html), desde cuyo portal web es posible también acceder a nuestra unidad.
ServiciosSeparación y purificación Fraccionamiento/purificación (AKTA FPLC)RotoforElectroforesis bidimensional DIGE Enriquecimiento en fosfoproteínas/fosfopéptidosConcentración y desalado (Zip-Tip)
Identificación de proteínasMALDI TOF MSNanoLC ESI-MS/MS (individuales y mezclas de proteínas)Western blot
Caracterización de proteínas (nanoESI-MS/MS en colaboración).

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
86
UNIDAD DE SERVICIOS DE PROTEOMICA DEL CIC bioGUNE Nere Alkorta, Eva Rodriguez, Vladimir Montero,
Stephanie Richer y Felix Elortza. CIC bioGUNE, Parque Tecnológico de Bizkaia, Edificio 801 A,
48160 Derio.
1. INTRODUCCIÓN
El Centro de Investigación Cooperativa bioGUNE es un centro de nueva creación con un compromiso con la investigación de calidad, que desarrolla su actividad en las principales áreas de la biotecnología aplicada a la salud: genómica funcional, metabolómica, biología celular y células madre, bioinformática y proteómica.
La Unidad de Servicios de Proteómica es una unidad joven, inaugurada en el CIC bioGUNE a principios del año 2004. Esta unidad se centra en el análisis mediante espectrometría de masas de proteínas y metabolitos. Este Servicio se ofrece a los diferentes grupos/laboratorios de investigación del CIC bioGUNE, empresas y centros adscritos al CIC bioGUNE, así como cualquier otro potencial usuario público o privado. El grupo que hemos formado es multidisciplinar y aunque joven, tiene una sólida experiencia en el campo de la proteómica y la metabolómica. 2. EQUIPAMIENTO
El CIC bioGUNE cuenta con dos sistemas diferentes para la
separación de proteínas y su análisis diferencial posterior. El sistema para electroforesis diferencial en gel (DIGE-2D) consta del scanner Typhoon TRIO, su correspondiente robot picador y el programa para el análisis de imágenes DeCyder (General Electric). El sistema Proteome Lab PF 2D (Beckman Coulter) permite separar proteínas mediante cromatografía líquida (cromato-isoenfoque y fase reversa) de manera automática y también cuenta con un software específico para el análisis diferencial de muestras. En cuanto al análisis mediante espectrometría de masas, contamos con los siguientes sistemas:
-La Plataforma MALDI consta de dos máquinas. El Proteineer dp de Bruker, es el robot que permite realizar automáticamente las digestiones de bandas/spots de geles y sus posterior preparación de muestras para MALDI. Para el análisis de las muestras mediante MALDI, contamos con un Ultraflex TOF/TOF. Este espectrómetro de masas permite, además de identificar proteínas mediante la técnica conocida como “huella peptídica”, la fragmentación de los péptidos seleccionados. Con ello se pueden secuenciar los mismos, permitiendo el análisis estructural del péptido en sí y hace que la identificación de la proteína en cuestión sea más fiable.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
87
Digestor Preparador Proteineer dp
Ultraflex TOF/TOF -El sistema de nano-cromatografía líquida acoplada “on-line” al espectrómetro de masa hibrido cuadrupolo tiempo de vuelo. Este sistema cuenta con el cromatógrafo nano Acquity, que como cuyo nombre indica permite trabajar con flujos reales (sin split) a escala nano (nano litros por minuto). Este cromatógrafo esta acoplado al Q-TOF Premier, espectrómetro de masas de ultima generación de la casa Waters/Micromass. Este equipo consta con los últimos avances de hardware y software para obtener la máxima sensibilidad y prestaciones, tanto para la identificación de proteínas como para la caracterización de modificaciones post-traduccionales. El sistema en su conjunto (cromatografía y espectrómetro) está controlado por un software integrado. El procesamiento y análisis de datos se realiza mediante el software ProteinLynx Global Server.
Nano Acquity acoplado al Q-TOF Premier -En este laboratorio también tenemos un sistema de cromatografía líquida Acquity conectado “on-line” mediante electro spray aun espectrómetro de tiempo de vuelo, LCT Premier (Waters/Micromass). Este equipo (como el anteriormente citado Q-TOF Premier) tiene un sistema “Lock-spray” que permite realizar experimentos con gran exactitud de masa en las mediciones (< 3 ppm) y su diseño facilita una alta sensibilidad y resolución. La gran resolución del nuevo sistema cromatográfico, la exactitud de masa en las mediciones y el software especifico dedicado al tratamiento estadístico de los datos, hacen que este sistema sea ideal para obtener perfiles metabólicos así como para la identificación de nuevos bio-marcadores.
Acquity acoplado al LCT Premier 3. PERSONAL Nere Alkorta es Técnico de laboratorio y Licenciada en Ciencias Químicas por la Universidad del País Vasco/Euskal

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
88
Herriko Unibertsitatea (UPV/EHU). Antes de incorporarse al grupo en Septiembre del 2004 ha trabajado en laboratorios de diferentes instituciones como INASMET y la Facultad de Farmacia de Vitoria/Gasteiz. Hoy en día es la encargada de la plataforma MALDI, incluyendo el manejo robot digestor preparador y el Ultraflex TOF/TOF. Vladimir Montero es Licenciado en ciencias Físicas y se doctoró en la Universidad de Río de Janeiro (Brasil). Su tesis esta directamente vinculada al tema de espectrometría de masa por tiempo de vuelo (TOF). Su experiencia abarca muchos de los aspectos relevantes en el uso y diseño de espectrómetros de masa del tipo TOF o TOF/TOF, el desarrollo de interfaces para el control y adquisición de datos en espectrómetros TOF y la preparación de muestras MALDI. Además de su experiencia en el diseño de estos aparatos, tiene también conocimientos de análisis e identificación de proteínas mediante MALDI-TOF. Su última etapa investigadora la ha llevado a cabo en la Universidad de Manitota (Canadá) desarrollando un proyecto sobre una celda de disociación inducida por electrones para los Q-TOF en colaboración con SCIEX. Vladimir se incorporará al equipo en enero del 2006 para reforzar la plataforma MALDI. Eva Rodriguez es Licenciada en Biología por la Universidad de Santiago de Compostela y de Bioquímica por la Universidad de Salamanca. Tras trabajar durante casi dos años en el Neuroscience Research Centre (Merck Sharp and Dohme, Reino Unido) se trasladó al Michael Barber Center for Mass Spectrometry (Manchester, Reino Unido) para realizar el doctorado sobre fosfoproteomas de células cancerosas. Durante su doctorado se ha especializado en el uso de diversas
metodologías propias de un laboratorio de proteómica como los geles bidimensionales, y técnicas de espectrometría de masas como el análisis de proteínas mediante MALDI-TOF y cromatografía a escala nano acoplada on-line a quadrupolo tiempo de vuelo (nLC-QTOF). Eva es la responsable del sistema nano UPLC-QTOF.
Uno de los objetivos del CIC bioGUNE es aunar esfuerzos, con el fin de desarrollar nuevas metodologías para el estudio proteómico y metabolómico, encauzados al descubrimiento de nuevos bio-marcadores de diagnóstico mediante técnicas de espectometría de masas. Con ese fin, la compañía OWL Genomics trabaja conjuntamente con la Unidad del Servicio de Proteómica en la persona de Stephanie Richer, investigadora experta en metabolómica y responsable del sistema LC-TOF y del posterior análisis estadístico de los datos obtenidos. Stephanie Richer, se licenció en ciencias Químicas en la Université Claude Bernard, Lyon (Francia) donde realizo su doctorado en Química Analítica. Durante su estancia postdoctoral en la National University of Ireland, Galway (Irlanda) trabajó con técnicas como GC-MS y LC-MS. Tras ese periodo, se incorporó en el equipo del European Centre for High-End Mass Spectrometry, que Waters/Micromass tiene en Almere (Holanda). En Waters trabajó durante dos años como técnico especialista de aplicaciones para pequeña molécula, centrándose en el uso de instrumentos del tipo TOF para estudios de metabolómica y metabonómica. Felix Elortza se licenció en Biología (especialidad bioquímica) en la UPV/EHU y allí realizó su tesis doctoral (2001). Durante esos años pudo combinar la labor investigadora con la docente y ejerció como profesor en la

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
89
misma universidad dando clases de Bioquímica General y Biología Molecular (Facultad de Ciencias de Leioa y Facultad de Químicas de San Sebastián). Tras doctorarse realizó su estancia postdoctoral bajo la supervisión del Dr. Ole N. Jensen en el “Protein Research Group”, Southern Denmark
University, Odense (Dinamarca) (2001-2003), conocido laboratorio de espectrometría de masas que fue fundado por el profesor Peter Roepstorff. Felix Elortza es el responsable de la Unidad de Servicios de Proteómica del CIC bioGUNE desde enero del 2004.
Nere, Felix, Eva y Stephanie. Vladimir se incorporará en breve.
4. EL SERVICIO La unidad de Servicios de proteómica tiene una página web en la que se da información de los detalles que servicio ofrece, como por ejemplo: que tipo de muestras se procesan, como preparar las muestras y consejos para evitar los problemas más típicos, precios etc. El pedido se realiza “on-line” y una vez que el cliente tiene su código, el sistema lo reconoce la próxima vez y de esa manera no hay necesidad de escribir todos los datos de facturación las
siguientes veces que se accede a el. Además, el sistema informático le permite al cliente saber en que fase del proceso se encuentra su pedido (petición accedida, muestras recibidas, en proceso de análisis o terminadas). Para cualquier tipo de consulta, podéis llamar al teléfono 94 406 1315 o también al 94 4061329 o bien visitar nuestra página web en http://www.cicbiogune.es/servicios/main.asp

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
90
GRUPO DE PROTEOMICA DE ALIMENTOS MARINOS
José Manuel Gallardo, Carmen Piñeiro, Mónica Carrera, Ignacio Ortea y Lorena Barros.
Dep. Tecnología de Alimentos. IIM (CSIC). Eduardo Cabello 6. 36208. Vigo.
En 1989, el entonces grupo de
Química de Productos Marinos, formaba parte de un proyecto europeo de investigación en el que, por medio de diversas técnicas electroforéticas (IEF, SDS-PAGE, CE), aplicadas al análisis de proteínas de músculo crudo, tratado con urea, con tripsina y/o bromuro de cianógeno, se trataba de recaracterizar proteínas específicas de diferentes especies marinas. A partir de 1995, este equipo de trabajo comienza a desarrollar técnicas de electroforesis bidimensional, dirigida al estudio de proteínas de merluzas comerciales, que permitió la separación de un gran número de proteínas características de estas especies y la comparación cualitativa y cuantitativa, mediante el empleo de programas de análisis de imágenes. De esta forma, el equipo se ha ido adentrando de manera gradual y más profunda en la proteómica, convirtiéndose parte del mismo en lo que hoy constituye el equipo de Proteómica de Productos Marinos.
Las limitaciones de la
electroforesis bidimensional y la necesidad de identificar y caracterizar diversas proteínas de origen marino, muchas de las cuales no están presentes
en las bases de datos, motivó
que se pensase en la espectrometría de masas, como una opción para la identificación de proteínas, para lo cual era imprescindible disponer de un espectrómetro de masas. A partir de esa fecha y gracias a la formación en técnicas de espectrometría de masas, adquirida en el Laboratorio de Química de Proteínas del Centro de Biología Molecular Severo Ochoa, se solicitó el primer proyecto de proteómica al Plan Nacional, en el Área de Tecnología de Alimentos, cuyo primer fruto fue una tesis doctoral leída en 2001, en la que se caracterizaban y secuenciaban “de novo” diversas proteínas de merluza.
El grupo de proteómica del
Instituto de Investigaciones Marinas de Vigo (CSIC) está formado en la actualidad por el Dr.José Manuel Gallardo( Profesor de Investigación del CSIC), Dra Carmen Piñeiro (Titulada Técnica Especialista), Mónica Carrera (Becaria de Postgrado I3P), Ignacio Ortea(Becario FPU) y Lorena Barros( Ayudante de Laboratorio contratada). La vocación fundamental del grupo es la investigación, sin descartar, en un futuro, la realización de determinados servicios, cuando los recursos humanos lo permitan.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
91
La financiación con la que cuenta el grupo proviene en la actualidad de dos proyectos concedidos por el Plan Nacional y otro por la Comunidad Autónoma. En dichos proyectos se colabora con diversos organismos e instituciones como el CBMSO, la Universidad Complutense, la Universidad de Santiago de Compostela y el Centro Tecnológico del Mar (CETMAR). Además, se ha recibido cofinanciación de la Dirección General de I+D de la Xunta de Galicia, de fondos FEDER y del propio CSIC, para la adquisición de un equipo de espectrometría de masas con trampa iónica y triple cuadrupolo, acoplado a un HPLC.
Las líneas de investigación en
las que se trabaja están encaminadas fundamentalmente al estudio de péptidos y/o proteínas procedentes de productos de la pesca cuya presencia y/o ausencia esté relacionada con la calidad y la seguridad alimentaria de los mismos y son las siguientes:
-Caracterización de péptidos
específicos de merluza y crustáceos, con
el fin de elaborar a partir de ellos kits de diagnóstico rápido, para la identificación en lonja de productos elaborados a partir de estas especies.
-Definición de proteínas indicadoras de vida útil en especies refrigeradas como el rodaballo de cultivo, la sardina y el jurel.
-Identificación de proteínas alergénicas termoestables en productos marinos.
-Identificación de proteinas indicadoras de estrés ambiental en especies de cultivo.
Desde el punto de vista
metodológico, estamos intentando implantar nuevas tecnologías proteómicas, como la cromatografía líquida bidimensional, para intentar obviar en parte la separación de proteínas en gel.
Gracias a estas líneas de investigación, se están realizando dos tesis doctorales y se han publicado diversos trabajos, que nos han permitido ser considerados como un equipo de investigación pionero en la aplicación de técnicas de proteómica al análisis de alimentos de origen marino y de la acuicultura.
La implantación en nuestro
grupo de toda esta serie de metodologías, ha supuesto un avance fundamental para el conocimiento de las proteínas de origen marino, poco estudiadas todavía, que nos va a permitir abordar en el futuro diversos temas de investigación, relacionados principalmente con la toxicología y nutrición y con los efectos de las dietas sobre los niveles de expresión de proteínas en diferentes especies acuícolas.
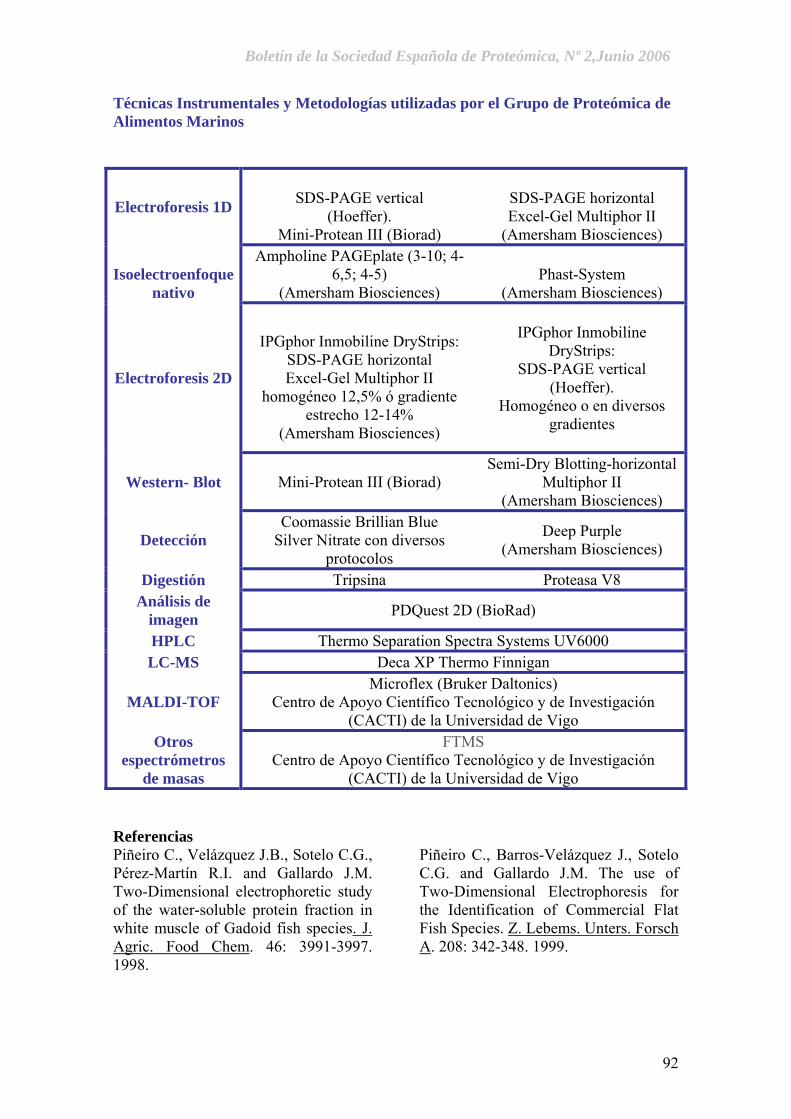
Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
92
Técnicas Instrumentales y Metodologías utilizadas por el Grupo de Proteómica de Alimentos Marinos
Electroforesis 1D
SDS-PAGE vertical
(Hoeffer). Mini-Protean III (Biorad)
SDS-PAGE horizontal Excel-Gel Multiphor II
(Amersham Biosciences)
Isoelectroenfoque nativo
Ampholine PAGEplate (3-10; 4-6,5; 4-5)
(Amersham Biosciences)
Phast-System
(Amersham Biosciences)
Electroforesis 2D
IPGphor Inmobiline DryStrips:
SDS-PAGE horizontal Excel-Gel Multiphor II
homogéneo 12,5% ó gradiente estrecho 12-14%
(Amersham Biosciences)
IPGphor Inmobiline
DryStrips: SDS-PAGE vertical
(Hoeffer). Homogéneo o en diversos
gradientes
Western- Blot Mini-Protean III (Biorad) Semi-Dry Blotting-horizontal
Multiphor II (Amersham Biosciences)
Detección Coomassie Brillian Blue
Silver Nitrate con diversos protocolos
Deep Purple (Amersham Biosciences)
Digestión Tripsina Proteasa V8 Análisis de
imagen PDQuest 2D (BioRad)
HPLC Thermo Separation Spectra Systems UV6000 LC-MS Deca XP Thermo Finnigan
MALDI-TOF Microflex (Bruker Daltonics)
Centro de Apoyo Científico Tecnológico y de Investigación (CACTI) de la Universidad de Vigo
Otros espectrómetros
de masas
FTMS Centro de Apoyo Científico Tecnológico y de Investigación
(CACTI) de la Universidad de Vigo
Referencias Piñeiro C., Velázquez J.B., Sotelo C.G., Pérez-Martín R.I. and Gallardo J.M. Two-Dimensional electrophoretic study of the water-soluble protein fraction in white muscle of Gadoid fish species. J. Agric. Food Chem. 46: 3991-3997. 1998.
Piñeiro C., Barros-Velázquez J., Sotelo C.G. and Gallardo J.M. The use of Two-Dimensional Electrophoresis for the Identification of Commercial Flat Fish Species. Z. Lebems. Unters. Forsch A. 208: 342-348. 1999.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
93
Piñeiro C., Vázquez J., Marina A.I., Barros-Velázquez J. and Gallardo J.M. Characterization and partial sequencing of species-specific sarcoplasmic polypeptides from commercial hake species by mass spectrometry following 2-DE analysis. Electrophoresis 22(8): 1545-1552. 2001
Piñeiro C.; Barros-Velázquez J; Vázquez J; Figueras A. and Gallardo J.M. Proteomics as a tool for the investigation of seafood and other marine products (Review). J. Proteome Research 2 (2): 127-135. 2003.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
94
UNIDAD DE PROTEÓMICA DE LA UNIVERSIDAD DE CÓRDOBA Samuel Ogueta, Consuelo Gómez, Raquel González, M.
Ángeles Castillejo Servicios Centrales de Apoyo a la Investigación (S.C.A.I.), Campus
Rabanales, Edificio Ramón y Cajal, 14071 Córdoba http://www.uco.es/servicios/scai/unidades/molecular/proteomica.htm
El creciente interés por parte de
diferentes grupos de investigación en la Comunidad de Andalucía en desarrollar proyectos con base proteómica, junto con una fuerte apuesta en el campo del estudio del proteoma por parte de la Universidad de Córdoba, dieron como fruto el nacimiento de la primera Unidad de Proteómica de Andalucía hace, apenas, un año (marzo de 2005), dentro de los Servicios Centrales de Apoyo a la Investigación (S.C.A.I.) de la propia Universidad de Córdoba (UCO).
Coincidiendo con su fundación, la Unidad de Proteómica de SCAI-UCO quedó englobada dentro de la Red Nacional de Servicios de Proteómica (ProteoRed) como grupo principal dentro del Nodo 6. Además, es la Unidad de Proteómica de la Plataforma Andaluza de Genómica, Proteómica y Bioinformática (PAGPB), creada por la Junta de Andalucía para un desarrollo competitivo en la región de estas tres especialidades. Esta apuesta de la Junta de Andalucía se ha traducido en el equipamiento de la Unidad con instrumental de última generación y en

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
95
la creación de contratos de personal técnico.
El trabajo desarrollado en la
Unidad de Proteómica se reparte entre el servicio a usuarios con la metodología ya establecida y la puesta a punto de nuevas metodologías de utilidad para nuestros usuarios.
En la actualidad, la Unidad consta
de las siguientes personas:
- Dr. Samuel Ogueta. Técnico Superior ProteoRed. Responsable y Coordinador de la Unidad. E-mail: [email protected]
- Dra. Consuelo Gómez. Técnico Superior. E-mail: [email protected]
- Dra. M. Ángeles Castillejo. Técnico PAGPB. E-mail: [email protected]
- Raquel Gónzalez. Técnico de Laboratorio PAGPB. E-mail: [email protected]
El equipamiento con el que contamos es el siguiente:
- Sistemas completos de electroforesis mono y bidimensional, para tamaños de gel de 7, 11 y 18 y 24cm, de BIO-RAD.
- Densitómetro GX-800 (BIO-RAD) para geles teñidos con cualquiera de los protocolos de tinción visible (Coomassie, plata, imidazol-zinc)
- Lector de fluorescencia FX (BIO-RAD) para geles, equipado con tres laceres.
- Programas PDQuest 7.0 y Quantity One 1.0 para análisis de imágenes.
- Estación de picado de geles ProPic, de Genomic Solutions
- Robot para digestión automática ProGest, de Genomic Solutions
- Robot para deposición de muestras digeridas sobre placas MALDI ProMS, de Genomic Solutions
- Cromatógrafo líquido HPLC Agilent 1100 con microrrecolector de fracciones.
- Colector de fracciones sobre placas MALDI o placas microtiter Probot (LCPackings) .
- Espectrómetro de masas MALDI-TOF/TOF, 4700 Promeomics Analyzer (Applied Biosystems)
- Espectrómetro de masas Electrospray-Trampa Lineal,

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
96
LTQ (Thermo Electron), acoplado a nanoHPLC Ultimate 3000 (LCPackings)
- Espectrómetro de masas Electrospray-Triple Cuadrupolo
/ Trampa Lineal, 4000 Q-Trap (Applied Biosystems), acoplado a nanoHPLC Switchos II/Ultimate Plus (LCPackings).
Hasta la fecha, los servicios ofertados por nuestra Unidad engloban tanto la Proteómica de Expresión, como la Proteomica de Interacción y la Proteómica de Segunda Generación. Como paso primordial antes de acometer un análisis, mantenemos una reunión con el cliente con el fin de adaptar toda la metodología existente a su necesidad concreta. Ofrecemos nuestros servicios a toda la comunidad científica, tanto pública, como privada, con una tabla de tarifas ajustada a la procedencia del usuario. La lista actual de nuestros servicios es la siguiente:
- Resolución de proteínas
mediante electroforesis mono y bidimensional.
- Identificación de proteínas a partir de gel mediante huella peptídica.
- Identificación de proteínas en solución mediante tándem nanoHPLC-Masas (LC-MS). Dependiendo de la complejidad de la muestra a analizar, se realiza cromatografía monodimensional (mezclas de hasta 30 proteínas) o cromatografía bidimensional (mezclas con más de 30 proteínas).
- Determinación de masas moleculares mediante MALDI-TOF.
- Separación de proteínas/péptidos mediante HPLC.
- Secuenciación de péptidos.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
97
Tenemos previsto ampliar la oferta de servicios en el presente año con los siguientes:
- Análisis de modificaciones en
proteínas (mutaciones,
modificaciones postraduccionales, etc.).
- Estudios de expresión diferencial con tecnología DIGE e iTRAQ.
- Proteómica Clínica.
Proteómica Clínica Tecnología iTRAQ
La acogida en este primer año de
funcionamiento la podemos clasificar como muy positiva, porque tenemos usuarios de la propia Universidad de Córdoba, de organismos y universidades de Andalucía, universidades del resto de España y centros internacionales.
En total, hemos analizado más de 1000 muestras con un porcentaje de éxito superior al 95%.
Además de ofrecer servicios, la Unidad de Proteómica SCAI-UCO, participa activamente en la difusión de la Proteómica mediante seminarios, cursos de doctorado y cursos especializados:
- Curso de Doctorado “Bioquímica de Proteínas y Proteómica”, años 2005 y 2006.
- Curso de la Universidad de Verano Corduba 2005 “Biotecnología: Marcadores moleculares, ingeniería genética y transformación genómica funcional e hibridación in situ, PCR cuantitativa a tiempo real y mezclas de DNA y proteómica”
- Curso Práctico sobre Proteómica 2006. Un paso adelante en el análisis de proteomas: iTRAQ, modificaciones Postraduccionales (nitrosilación).
- Seminario “Proteómica: generalidades, técnicas y aplicaciones en investigación”, Centro de la Grasa, Sevilla, Octubre 2005.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
98
Instrucciones a los autores
Las contribuciones al Boletín de la SEProt deberán ajustarse a uno de los formatos descritos a continuación. No es necesario ser socio de la SEProt para publicar en el Boletín. En todos los casos los textos deberán estar en formato MS-Word y el material gráfico en formatos .jpg, .tif, .gif, o .ppt. Caso de incluirse cuadros de texto o tablas, el formato preferido de éstos será pdf. La resolución de las figuras o fotografías deberá ser de al menos 300 dpi, y su tamaño deberá ser el real de publicación. Las contribuciones deberán ser enviadas por correo electrónico a uno de los editores, Jesús Jorrín ([email protected]), Juan J. Calvete ([email protected]) o Jesús Vázquez ([email protected])
Excepto las notas técnicas y los artículos de empresas, la publicación en el Boletín es gratuita.
CARTAS AL EDITOR
Las Cartas al Editor se publicarán en la sección "Correspondencia" y su longitud no deberá exceder de aproximadamente 400 palabras. El contenido de las Cartas es libre pero se recomienda que verse sobre algún tema de interés amplio, o que esté relacionado con algún artículo o carta publicada en un número anterior del Boletín. Las Cartas al Editor serán publicadas a discreción de los editores. La definición de "Cartas al Editor" es amplia. Una modalidad de carta puede ser una reseña de libro, artículo (propio o ajeno), Tesis Doctoral, reunión científica pasada o futura, e incluso una viñeta cómica sobre temas científicos o de política científica. ARTÍCULOS DE OPINIÓN
Los artículos de opinión no deberán sobrepasar las 3000 palabras y tratarán sobre temas científicos, preferentemente relacionados con la Proteómica, o de política científica de actualidad y amplio interés. Si bien los artículos y colaboraciones publicados en el Boletín reflejan la opinión de sus autores y no necesariamente la opinión de la dirección de la SEProt, el criterio general que se seguirá para la aceptación de un artículo en el Boletín será que su contenido no esté en desacuerdo con las ideas defendidas por la SEProt y reflejadas en sus Estatutos, que pueden consultarse en este Boletín así como en el portal electrónico de la Sociedad (http://www.cbm.uam.es/seprot). No obstante, se aceptarán artículos que discutan o critiquen de forma científica y correcta las ideas dominantes en la SEProt. ARTÍCULOS CIENTÍFICOS
Los artículos científicos tendrán una extensión máxima de alrededor de 5000 palabras, en español (preferentemente) o en inglés. El formato será libre aunque se recomienda un estilo de divulgación, ya que se pretende que puedan ser leídos por científicos no expertos en el tema. Caso de que un artículo contenga figuras ya publicadas, será responsabilidad exclusiva del autor obtener los permisos necesarios para reproducirlas en el Boletín de la SEProt. También la denominación de artículo científico será considerada en un sentido amplio. Se incluirán en este apartado Notas Técnicas y Protocolos de Laboratorio siempre y cuando éstos incluyan la suficiente información para ser reproducidos por otros. El contenido de los artículos será revisado por un especialista en la materia, quien aconsejará sobre la conveniencia o no de su publicación en el Boletín.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
99
Impresos de
Inscripción a la
SEProt

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
100
Sede social: Instituto de Biomedicina de Valencia, C.S.I.C. c/Jaime Roig 11, 46010 Valencia. Tel.: 96 339 1778; Fax: 96 369 0800 C.I.F. G97465629
Nº Registro Nacional de Asociaciones: 584180
http://www.cbm.uam.es/seprot
BOLETÍN DE INSCRIPCIÓN
(Socio ordinario/Socio Estudiante)2 Enviar el formulario relleno, incluida la orden de pago al banco, por e-mail o FAX. En caso de talón bancario, enviarlo por correo. El envío se hará al Secretario de la SEProt: Dr. Jesús V. Jorrín Novo. Edificio Severo Ochoa (C6), Campus Rabanales, Universidad de Córdoba 14071 Córdoba Tfno 957 218574, 618 808041 FAX 957 218592 E-mail:[email protected] 1. Datos personales y profesionales Nombre y apellidos: DNI-NIF/Pasaporte: Situación profesional: Estudiante de doctorado [ ], Postdoctoral [ ], Plantilla (especificar) [ ]: Grupo, Departamento, Institución: Dirección: Otra dirección de contacto: Teléfono: FAX: Correo electrónico: Área de investigación/interés:

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
101
2. Tipo de Socio3 [ ] Socio Ordinario [ ] Socio Estudiante 3. Presentado por los siguientes Socios de la SEProt: 2. Cuota de inscripción [ ] Cuota ordinaria de 40 Euros [ ] Cuota de estudiante de doctorado (adjuntar una carta del director): 20 Euros 3. Forma de Pago [ ] Mediante domiciliación bancaria (utilice el impreso de la página siguiente)4 [ ] Mediante talón bancario a nombre de la Sociedad Española de Proteómica Lugar y fecha:
Firmado:
3 Artículo 25 de los Estatutos de la SEProt. b) Socios Ordinarios: aquellos que ingresen después de la constitución de la SEProt. Podrán ser Socios Ordinarios los científicos con interés en el campo de la Proteómica. Las propuestas deberán ser hechas, en impreso facilitado por la Sociedad, por dos Socios Ordinarios o uno de Honor, y se enviarán, en los plazos que se establezcan, al Secretario de la Sociedad, quién les dará trámite. e) Socios Estudiantes: Pre-doctorales que deseen acogerse, por un máximo de cuatro años, a un régimen de cuota reducida. La solicitud de incorporación como Socio Estudiante deberá ser presentada por un Socio Ordinario y será resuelta directamente por la Junta Directiva. Los Socios Estudiantes podrán asistir a todas las sesiones de la Asamblea General con voz y voto. Todos los Socios, excepto los de Honor y los que se encuentran en situación de jubilados, deberán abonar una cuota anual que se fijará para cada tipo de Socio por la Asamblea General a propuesta de la Junta Directiva. 4 Enviar una copia al banco

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
102
ORDEN DE PAGO DE LA CUOTA ANUAL DE SOCIO DE LA
“SOCIEDAD ESPAÑOLA DE PROTEÓMICA (SEProt)”
Autorizo a cargar la cuota anual de socio de la “Sociedad Española de Proteómica “ a mi cuenta Banco: Oficina/Sucursal: Dirección postal: Código de cuenta (banco/sucursal/DC/Cuenta (10 dígitos)
, a de de 200
Firmado:

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
103
Sede social: Instituto de Biomedicina de Valencia, C.S.I.C. c/Jaime Roig 11, 46010 Valencia. Tel.: 96 339 1778; Fax: 96 369 0800 C.I.F. G97465629
Nº Registro Nacional de Asociaciones: 584180
http://www.cbm.uam.es/seprot
BOLETÍN DE INSCRIPCIÓN
(Socio Patrocinador)5,2 Enviar el formulario relleno, incluida la orden de pago al banco, por e-mail o FAX. En caso de talón bancario, enviarlo por correo. El envío se hará al Secretario de la SEProt: Dr. Jesús V. Jorrín Novo. Edificio Severo Ochoa (C6), Campus Rabanales, Universidad de Córdoba 14071 Córdoba Tfno 957 218574, 618 808041 FAX 957 218592 E-mail:[email protected] Empresa/Institución: Razón Social: Teléfono: FAX: Correo Electrónico: Página Web: Persona de Contacto: Ámbito de actuación/Área de interés: Autorizo a incluir el logo de la compañía/institución y un link a su correspondiente página web en la web de la SEProt. 5Artículo 25d de la SEProt: Socios Patrocinadores: los institutos, organizaciones, asociaciones y empresas que pretendan incrementar el desarrollo de la SEProt proporcionando medios y fondos para su actividad. Los socios Patrocinadores deberán ser aceptados por la Asamblea General a propuesta de la Junta Directiva. Tendrán derecho a voto en las Asambleas, pero no podrán ser elegibles para cargos directivos. 2 La Cuota es de 600 Euros anuales. Se incluirá en la página web de la SEProt el logo de la compañía/institución y un link a la correspondiente página web. Se incluirá el logo en los boletines que edite la sociedad.

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
104
[ ] SI [ ] NO 3. Forma de Pago [ ] Mediante domiciliación bancaria (utilice el impreso de la página siguiente)♣ [ ] Mediante talón bancario a nombre de la Sociedad Española de Proteómica Lugar, fecha y firma:
♣ Enviar una copia al banco

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
105
DIRECTORIO DE MIEMBROS DE LA SOCIEDAD ESPAÑOLA
DE PROTEÓMICA (ABRIL’06)

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
106
ABIÁN MOÑUX, JOAQUÍN CONSEJO SUPERIOR DE INVESTIGACIONES CIENTÍFICAS INSTITUTO DE INVESTIGACIONES BIOMÉDICAS DE BARCELO Tel: 933638311 Fax: 933633801 Email: [email protected] ALBAR RAMÍREZ, JUAN PABLO CONSEJO SUPERIOR DE INVESTIGACIONES CIENTÍFICAS INSTITUTO NACIONAL DE BIOTECNOLOGÍA Tel: 915854696 Fax: 915854506 Email: [email protected] ANDREU MARTÍNEZ, DAVID UNIVERSIDAD POMPEU FABRA CIENCIAS EXPERIMENTALES Y DE LA SALUD Tel: 935422934 Fax: 935422802 Email: [email protected] ARIZMENDI BASTARRIKA, JESÚS M. UNIVERSIDAD DEL PAÍS VASCO DPTO. DE BIOQUÍMICA Y BIOLOGÍA MOLECULAR Tel: 946012615 Fax: 946013500 Email: [email protected] BAÑÓN MANEUS, ELISENDA HOSPITAL CLÍNICO DE BARCELONA UNITAT DE TRASPLANTEMENT RENAL
LAB. HORMONOLOGÍA Tel: 636939599 Email: [email protected] BARBOSA TORRALBO, JOSÉ UNIVERSIDAD DE BARCELONA DPTO. DE QUÍMICA ANALÍTICA Tel: 934021279 Fax: 934021233 Email: [email protected] BÁRCENA RUIZ, JOSÉ ANTONIO UNIVERSIDAD DE CÓRDOBA DPTO. DE BIOQUÍMICA Y BIOLOGÍA MOLECULAR Tel: 957218590 Fax: 957218592 Email: [email protected] BARRANCO MEDINA, SERGIO CENTRO SUPERIOR DE INVESTIGACIONES CIENTÍFICAS ESTACIÓN EXPERIMENTAL DEL ZAIDÍN Tel: 958181600 Fax: 958129600 Email: [email protected] BARROS VELÁZQUEZ, JORGE UNIVERSIDAD DE SANTIAGO DE COMPOSTELA DPTO. DE QUÍMICA ANALÍTICA, NUTRICIÓN Y BROMATOLOGÍA Tel: 600942264 Fax: 986649266 Email: [email protected]

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
107
BLANCO GARCÍA, FRANCISCO J. HOSPITAL JUAN CANALEJO UNID. INV. OSTEOARTICULAR Tel: 981178272 Fax: 981178273 Email: [email protected] BLÁZQUEZ GUTIÉRREZ, M. DOLORES CAI-GENÓMICA Y PROTEÓMICA (UCM-PCM) UNIDAD DE PROTEÓMICA Tel: 913941613 Fax: 913941745 Email: [email protected] BODAS, BERNABÉ THERMO ELECTRON Tel: 914845957 Fax: 916615572 Email: [email protected] BORRÁS RAMÍREZ, EVA UNIVERSIDAD POMPEU FABRA CIENCIAS EXPERIMENTALES Y DE LA SALUD Tel: 935422825 Fax: 935422802 Email: [email protected] BRU MARTÍNEZ, ROQUE UNIVERSIDAD DE ALICANTE DPTO. DE AGROQUÍMICA Y BIOQUÍMICA Tel: 965903400 (3113) Fax: 965903880 Email: [email protected] CABEZÓN SORIANO, VIRGINIA UNIVERSIDAD COMPLUTENSE DE MADIRD DPTO. DE MICROBIOLOGÍA II Tel: 626446806 Fax: 913941745 Email: [email protected] CALVETE CHORNET, JUAN JOSÉ INSTITUTO DE BIOMEDICINA DE VALENCIA
DPTO. DE GENÓMICA Y PROTEÓMICA Tel: 96 339 1775 Fax: 96 369 0800 Email: [email protected] CANALS SURIS, FRANCESC INSTITUT DE RECERCA DEL HOSPITAL UNIVERSITARI VALL D'HEBRON LABORATORIO DE PROTEÓMICA Tel: 934894175 Fax: 932746026 Email: [email protected] CARRASCAL PÉREZ, MONTSERRAT CONSEJO SUPERIOR DE INVESTIGACIONES CIENTÍFICAS INSTITUTO INVESTIGACIONES BIOMÉDICAS DE BARCELONA Tel: 933638311 Fax: 933638301 Email: [email protected] CARRERA MOURIÑO, MÓNICA CONSEJO SUPERIOR DE INVESTIGACIONES CIENTÍFICAS DPTO. DE QUÍMICA DE PRODUCTOS MARINOS Tel: 986231930 Email: [email protected] CASADO VELA, JUAN CENTRO NACIONAL INVESTIGACIONES ONCOLÓGICAS UNIDAD DE TECNOLOGÍA DE PROTEÍNAS Tel: 912246900 Fax: 912246980 Email: [email protected] CASAL ÁLVAREZ, IGNACIO CNIO UNIDAD DE TECNOLOGÍA DE PROTEÍNAS Tel: 912246900 Fax: 912246972 Email: [email protected]

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
108
CASTILLEJO SÁNCHEZ, M. ÁNGELES UNIVERSIDAD DE CÓRDOBA DPTO. DE BIOQUÍMICA Y BIOLOGÍA MOLECULAR Tel: 957218574 Fax: 957218592 Email: [email protected] CHICANO GÁLVEZ, EDUARDO UNIVERSIDAD DE CÓRDOBA DPTO. DE BIOQUÍMICA Y BIOLOGÍA MOLECUAR Email: [email protected] CHIVA RODRÍGUEZ, CRISTINA UNIVERSIDAD POMPEU FABRA CIENCIAS EXPERIMENTALES Y DE LA SALUD Tel: 935422825 Fax: 935422802 Email: [email protected] COLLANTES DE TERÁN ESCRIBANO, LAURA UNIVERSIDAD DE SEVILLA DPTO. DE BIOQUÍMICA Tel: 954559902 Fax: 954233765 Email: [email protected] CORRALES IZQUIERDO, FERNANDO J. UNIVERSIDAD DE NAVARRA CENTRO DE INVESTIGACIÓN MÉDICA APLICADA Tel: 948194700 (4006) Fax: 948194717 Email: [email protected] CURTO RUBIO, MIGUEL UNIVERSIDAD DE CÓRDOBA DPTO. DE BIOQUÍMICA Y BIOLOGÍA MOLECULAR Tel: 957218574 Fax: 957218592 Email: [email protected] DE OLIVEIRA CACHEADO, ELIANDRE PARC CIENTÍFIC DE BARCELONA
PLATAFORMA DE PROTEÓMICA Tel: 934034653 Fax: 934037206 Email: [email protected] DÍEZ MASA, JOSÉ CARLOS CENTRO SUPERIOR DE INVESTIGACIONES CIENTÍFICAS INSTITUTO DE QUÍMICA ORGÁNICA GENERAL DPTO DE ANÁLISIS INSTRUMENTAL Y QUÍMICA AMBIENTAL Tel: 915622900 Fax: 915644853 Email: [email protected] DURÁN RUIZ, MARÍA DEL CARMEN UNIVERSITY COLLEGE OF LONDON LUDWIG INSTITUTE FOR CANCER RESEARCH Tel: 0044207679220 (0034609314576) Fax: 004420715081 Email: [email protected] ECHEVARRÍA ZOMEÑO SIRA UNIVERSIDAD DE CÓRDOBA ETSIAM Tel.: 957218439 Fax: 957218439 Email: [email protected] ELORTZA BASTERRIKA, FÉLIX CIC-BIOGUNE PARQUE TECNOLÓGICO DE BIZKAIA Tel: 944061315 Fax: 944061301 Email: [email protected] ESPUÑA NOVELL, GEMMA UNIVERSIDAD POMPEU FABRA CIENCIAS EXPERIMENTALES Y DE LA SALUD Tel: 935422825 Fax: 935422802 Email: [email protected] ESTANYOL, JOSEP MARÍA UNIVERSIDAD DE BARCELONA DPTO. DE BIOLOGÍA CELULAR

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
109
Tel: 934034798 Fax: 934021907 Email: [email protected] FERNÁNDEZ ACERO, FRANCISCO JAVIER UNIVERSIDAD DE CÁDIZ DPTO. DE MICROBIOLOGÍA Tel: 956016424 Fax: 956016181 Email: [email protected] FERNÁNDEZ LÓPEZ, ALMUDENA UNIVERSIDAD DE CÓRDOBA DPTO. DE MICROBIOLOGÍA Tel: 957212517 Fax: 957212517 Email: [email protected] FERNÁNDEZ IRIGOYEN, JOAQUÍN CENTRO PARA LA INVESTIGACION MEDICA APLICADA DPTO. DE TERAPIA GÉNICA Y HEPATOLOGÍA Tel: 948194700 (4005) Fax: 948194717 Email: [email protected] FERNÁNDEZ-ARENAS HERVÁS, ELENA UNIVERSIDAD COMPLUTENSE DE MADRID DPTO. DE MICROBIOLOGÍA II Tel: 913941614 Fax: 913941745 Email: [email protected] FLORIANO PARDAL, BELÉN UNIVERSIDAD PABLO OLAVIDE DPTO. DE CIENCIAS AMBIENTALES Tel: 954349053 Fax: 954349376 Email: [email protected] FORNÉ I FERRER, IGNASI CENTRO SUPERIOR DE INVESTIGACIONES CIENTÍFICAS INSTITUTO DE INVESTIGACIONES BIOMEDICAS DE BARCELONA
Tel: 933638311 Fax: 933638301 Email: [email protected] FUENTES GARCÍA, MANUEL CONSEJO SUPERIOR DE INVESTIGACIONES CIENTÍFICAS INSTITUTO DE CATÁLISIS Tel: 915854809 Fax: 915854760 Email: [email protected] GALLARDO ABUIN, JOSÉ MANUEL CENTRO SUPERIOR DE INVESTIGACIONES CIENTÍFICAS INSTITUTO DE INVESTIGACIONES MARINAS DPTO. QUÍMICA DE PRODUCTOS MARINOS Tel: 986231930 Fax: 986292762 Email: [email protected] GARCÍA ALONSO, ÁNGEL UNIVERSIDAD DE SANTIAGO DE COMPOSTELA VICERRECTORADO DE INVESTIGACIÓN Tel.: 981 563100 Fax: 981 547077 Email: GIL GARCÍA, CONCHA UNIVERSIDAD COMPLUTENSE DE MADRID DEPARTAMENTO DE MICROBIOLOGÍA II Tel: 913941748 Fax: 913941745 Email: [email protected] GÓMEZ ARIZA, JOSÉ LUIS UNIVERSIDAD DE HUELVA DPTO. DE QUÍMICA Y CIENCIA DE LOS MATERIALES Tel: 959019968 Fax: 959019942 Email: [email protected]

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
110
GÓMEZ DÍAZ,CONSUELO UNIVERSIDAD DE CÓRDOBA SERVICIO CENTRAL DE APOYO A LA INVESTIGACIÓN Tel: 957218984 Fax: 957218733 Email: [email protected] GÓMEZ MARTÍ, JOSEP SARTORIUS S.A. Tel: 932802623 Fax: 932801581 Email: [email protected] GONZÁLEZ FERNÁNDEZ, RAQUEL UNIVERSIDAD DE CÓRDOBA DEPARTAMENTO DE BIOQUÍMICA Y BIOLOGÍA MOLECULAR Tel: 957218686 Fax: 957218592 Email: [email protected] GUTIÉRREZ GIL, JUAN FRANCISCO UNIVERSIDAD DE SEVILLA DPTO. DE BIOQUÍMICA, BROMATOLOGÍA, TOXICOLOGÍA Y MEDICINA LEGAL Tel: 954559902 Email: [email protected] HERNAÉZ SÁNCHEZ, M. LUISA CAI-GENÓMICA Y PROTEÓMICA (UCM-PCM) UNIDAD DE PROTEÓMICA Tel: 913941613 Fax: 913941748 Email: [email protected] HERNÁNDEZ BARBADO, ROSA UNIVERSIDAD COMPLUTENSE DE MADRID DPTO. DE MICROBIOLOGÍA II Tel: 913941755 (696916955) Fax: 913941745 Email: [email protected] HUERTAS ROMERA, Mª JOSÉ UNIVERSIDAD DE CÓRDOBA DEPARTAMENTO DE BIOQUÍMICA Y BIOLOGÍA MOLECULAR
Tel: 957218318 Fax: 957218588 Email: [email protected] INSENSER NIETO, MARÍA UNIVERSIDAD COMPLUTENSE DE MADRID DPTO. DE MICROBIOLOGÍA II Tel: 913941834 Fax: 913941745 Email: [email protected] JOFRÉ I FRADERA, ANNA INSETITUT DE RECERCA I TECNOLOGIA AGROALIMENTARIA CENTRE DE TECNOLOGIA DE LA CARN Tel: 972630052 Fax: 972630373 Email: [email protected] JORGE CERRUDO, INMACULADA UNIVERSIDAD DE CÓRDOBA DPTO. DE BIOQUÍMICA Y BIOLOGÍA MOLECULAR Tel: 957218574 Fax: 957218592 Email: [email protected] JORGE GARCÍA, ALBERTO CSIC-UAM CENTRO DE BIOLOGÍA MOLECULAR SEVERO OCHOA Tel: 914978364 Fax: 914978087 Email: [email protected] JORRÍN NOVO, JESÚS V. UNIVERSIDAD DE CÓRDOBA DPTO. DE BIOQUÍMICA Y BIOLOGÍA MOLECULAR Tel: 957218574 Fax: 957218592 Email: [email protected] JOVER COBOS, MARÍA UNIVERSIDAD DE SEVILLA FACULTAD DE FARMACIA Tel: 657038931

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
111
Email: [email protected] JUÁREZ GÓMEZ, PAULA CENTRO SUPERIOR DE INVESTIGACIONES CIENTÍFICAS INSTITUTO DE BIOMEDICINA DE VALENCIA Tel: 963391778 Fax: 963690800 Email: [email protected] KAYALI SAYADI, NOUR UNIVERSIDAD COMPLUTENSEDE MADRID SERVICIO ESPECTROMETRÍA DE MASAS Tel: 913944160 Fax: 913944160 Email: [email protected] LARA LÓPEZ, MARÍA DEL PILAR APPLIED BIOSYSTEMS PROTEOMICS Tel: 615508606 Email: [email protected] LLAMAS MORENO, MARÍA JOSÉ CONSEJO SUPERIOR DE INVESTIGACIONES CIENTÍFICAS INSTITUTO DE AGRICULTURA SOSTENIBLE Tel: 957499242 Fax: 957499252 Email: [email protected] LÓPEZ, DANIEL CSIC-UAM CENTRO DE BIOLOGÍA MOLECULAR "SEVERO OCHOA" Tel: 914978364 Fax: 914978087 Email: [email protected] MALDONADO ALCONADA, ANA MARÍA UNIVERSIDAD DE CÓRDOBA DPTO. DE BIOQUÍMICA VEGETAL Y AGRÍCOLA Tel: 957218574
Fax: 957218592 Email: [email protected] MARCILLA DÍAZ, ANTONIO UNIVERSIDAD DE VALENCIA FACULTAD DE FARMACIA Tel.: 963543141 Tel.: 963544769 Email: [email protected] MARÍN MARCHANTE, MIGUEL CENTRO SUPERIOR DE INVESTIGACIONES CIENTÍFICAS INSTITUTO DE BIOMEDICINA DE VALENCIA Tel: 617030771 Fax: 963690800 Email: [email protected] MARINA RAMÍREZ, ANA ISABEL CSIC-UAM CENTRO DE BIOLOGÍA MOLECULAR SEVERO OCHOA Tel: 914978364 Fax: 914978087 Email: [email protected] MARTÍNEZ LÓPEZ, RAQUEL UNIVERSIDAD COMPLUTENSE DE MADRID DPTO. DE MICROBIOLOGIA II Tel: 913941755 Fax: 913941745 Email: [email protected] MARTÍNEZ ZORZANO, VICENTA SOLEDAD UNIVERSIDAD DE VIGO DPTO. DE BIOQUÍMICA, GENÉTICA E INMUNOLOGÍA Tel: 985812572 Fax: 986812556 Email: [email protected] MARTÍNEZ GOMARIZ, MONTSERRAT CAI-GENÓMICA Y PROTEÓMICA UNIDAD DE PROTEÓMICA Tel: 913941613 Fax: 913941745

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
112
Email: [email protected] MARTÍNEZ RUIZ, ANTONIO CENTRO NACIONAL DE INVESTIGACIONES CARDIOVASCULARES Tel: 918061880 Email: [email protected] MARTÍNEZ-SOLANO GONZÁLEZ, LAURA UNIVERSIDAD COMPLUTENSE DE MADRID DPTO. DE MICROBIOLOGÍA II Tel: 913941755 Fax: 913941745 Email: [email protected] MOLERO MARTÍN-PORTUGUÉS, GLORIA UNIVERSIDAD COMPLUTENSE DE MADRID DPTO. DE MICROBIOLOGÍA II Tel: 913941834 Fax: 913941745 Email: [email protected] MONTEOLIVA DÍAZ, LUCÍA UNIVERSIDAD COMPLUTENSE DE MADRID DPTO. DE MICROBIOLOGÍA Tel: 913941755 Fax: 913941745 Email: [email protected] MORENO LÓPEZ, ÁNGELA UNIVERSIDAD DE CÓRDOBA DPTO. DE GENÉTICA Tel: 957218730 Fax: 957218730 Email: [email protected] MORO RODRÍGUEZ, ERNESTO UNIVERSIDAD REY JUAN CARLOS DPTO. DE CIENCIAS DE LA SALUD Tel: 914888865 Fax: 914888831 Email: [email protected] MUÑOZ PERALTA, JAVIER
CENTRO DE INVESTIGACIÓN MÉDICA APLICADA DPTO. DE NEUROCIENCIAS Tel: 948194700 (4005) Fax: 948194717 Email: [email protected] NAVARRO ÁLVAREZ, PEDRO JOSÉ UNIVERSIDAD AUTONÓMICA DE MADRID CENTRO DE BIOLOGÍA MOLECULAR SEVERO OCHOA Tel.: 91 4978364 Email: [email protected] NAVAS IGLESIAS, NATALIA ÁFRICA UNIVERSIDAD DE GRANADA FACULTAD DE CIENCIAS Tel.: 958 243388 Fax: 958 243328 Email: [email protected] NAVAS SÁNCHEZ, ALFONSO G. CONSEJO SUPERIOR DE INVESTIGACIONES CIENTÍFICAS MUSEO NACIONAL DE CIENCIAS NATURALES Tel: 914111328 Email: [email protected] NOMBELA CANO, CÉSAR UNIVERSIDAD COMPLUTENSE DE MADRID DPTO. DE MICROBIOLOGÍA Tel: 913942084 Fax: 913941745 Email: [email protected] NÚÑEZ MORENO, YOLANDA CSIC-UAM CENTRO DE BIOLOGÍA MOLECULAR SEVERO OCHOA Tel: 914978364 Fax: 914978087 Email: [email protected] OGUETA, SAMUEL UNIVERSIDAD DE CÓRDOBA

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
113
SERVICIO CENTRAL DE APOYO A LA INVESTIGACIÓN Tel: 957218984 Fax: 957218733 Email: [email protected] OLEAGA PÉREZ, ANA CONSEJO SUPERIOR DE INVESTIGACIONES CIENTÍFICAS INSTITUTO DE RECURSOS NATURALES Y AGROBIOLOGÍA Tel: 923219606 Fax: 923219609 Email: [email protected] PÁEZ DE LA CADENA TORTOSA, MARÍA UNIVERSIDAD DE VIGO DPTO. DE BIOQUÍMICA, GENÉTICA E INMUNOLOGÍA Tel: 986812575 Fax: 986812556 Email: [email protected] PALOMARES RIUS, JUAN EMILIO UNIVERSIDAD DE CÓRDOBA DPTO. DE BIOQUÍMICA Y BIOLOGÍA MOLECULAR Tel: 957218574 Email: [email protected] PÉREZ SÁNCHEZ, RICARDO CONSEJO SUPERIOR DE INVESTIGACIONES CIENTÍFICAS INSTITUTO DE RECURSOS NATURALES Y AGROBIOLOGÍA Tel: 923219606 Fax: 923219609 Email: [email protected] PIÑEIRO GONZÁLEZ, CARMEN CONSEJO SUPERIOR DE INVESTIGACIONES CIENTÍFICAS DPTO. DE QUÍMICA DE PRODUCTOS MARINOS Tel: 986231930 Fax: 986292762 Email: [email protected]
PRADOS ROSALES, RAFAEL CARLOS UNIVERSIDAD DE CÓRDOBA DEPARTAMENTO DE GENÉTICA Tel: 957218981 Fax: 9572172 Email: [email protected] QUESADA MORAGA, ENRIQUE UNIVERSIDAD DE CÓRDOBA DPTO. C.R.A.F. UNIDAD DE ENTOMOLOGÍA AGRÍCOLA Tel: 957218475 Fax: 957218440 Email: [email protected] RAMÍREZ BOO, MARÍA UNIVERSIDAD DE CÓRDOBA DPTO. DE GENÉTICA Tel: 957218730 Email: [email protected] RAMOS FERNÁNDEZ, ANTONIO UNIVERSIDAD AUTONÓMICA DE MADRID CENTRO DE BIOLOGÍA MOLECULAR SEVERO OCHOA Tel.: 91 4978364 Email: [email protected] REQUEJO AGUILAR, RAQUEL UNIVERSIDAD DE CÓRDOBA DPTO. DE BIOQUÍMICA Y BIOLOGÍA MOLECULAR Tel: 957218686 Email: [email protected] RODA NOGUERA, ORIOL UNIVERSIDAD POMPEU FABRA CIENCIAS EXPERIMENTALES Y DE LA SALUD Tel: 935422825 Fax: 935422802 Email: [email protected] RODRÍGUEZ BERROCAL, FCO. JAVIER UNIVERSIDAD DE VIGO

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
114
DPTO. DE BIOQUÍMICA, GENÉTICA E INMUNOLOGÍA Email: [email protected] RODRÍGUEZ ORTEGA, MANUEL JOSÉ CHIRON VACCINES DPTO. DE BIOQUÍMICA Y BIOLOGÍA MOLECULAR Tel: 00390577245061 Fax: 00390577243564 Email: [email protected] RODRÍGUEZ PIÑEIRO, ANA MARÍA UNIVERSIDAD DE VIGO DPTO. DE BIOQUÍMICA, GENÉTICA E INMUNOLOGÍA Tel: 986813841 Fax: 986812556 Email: [email protected] ROMERO RUIZ, ANTONIO UNIVERSIDAD DE CÓRDOBA DPTO. DE BIOQUÍMICA Y BIOLOGÍA MOLECULAR Tel: 957218082 Fax: 957218688 Email: [email protected] RUIZ ROMERO, CRISTINA HOSPITAL JUAN CANALEJO UNIV. INV. OSTEOARTICULAR Tel: 981178272 Email: [email protected] SÁNCHEZ DEL PINO, MANUEL M. FUNDACIÓN VALENCIANA DE INVESTIGACIONES BIOMÉDICAS DPTO. DE QUÍMICA MÉDICA Tel: 963289680 (1301) Email: [email protected] SÁNCHEZ RUIZ, YOVAN UNIVERSITÀ DEGLI STUDI DI MILANO MASS SPECTROMETRY UNIT Tel: 390226432387 Email: [email protected]
SANCHO LÓPEZ, JAIME CENTRO SUPERIOR DE INVESTIGACIONES CIENTÍFICAS INSTITUTO DE PARASITOLOGÍA Y BIOMEDICINA DPTO. DE BIOLOGÍA CELULAR E INMUNOLOGÍA Tel: 958181664 Fax: 958181632 Email: [email protected] SANTAMARÍA MARTÍNEZ, ENRIQUE CENTRO PARA LA INVESTIGACIÓN MÉDICA APLICADA DPTO. DE TERAPIA GÉNICA Y HEPATOLOGÍA Tel: 948194700 (4005) Fax: 948194717 Email: [email protected] SANZ NEBOT, M.VICTORIA UNIVERSIDAD DE BARCELONA DPTO. DE QUÍMICA ANALÍTICA Tel: 934039116 Fax: 934021233 Email: [email protected] SANZ SANZ, LIBIA CENTRO SUPERIOR DE INVESTIGACIONES CIENTÍFICAS INSTITUTO DE BIOMEDICINA DE VALENCIA Tel: 963391778 Fax: 963690800 Email: [email protected] SEGURA ARGULLÓ, MIREIA UNIVERSIDAD POMPEU FABRA PARQUE DE INVESTIGACIÓN BIOMÉDICA DE BARCELONA Tel.: 935422825 Fax: 935422802 Email: [email protected] SERRANO RIVERA, HORACIO UNIVERSIDAD AUTÓNOMA DE MADRID FACULTAD DE CIENCIAS Tel.: 91 4313910/001 787 881-1982

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
115
Fax: 91 4974799 Email: [email protected]
[email protected] SESMA AGURRE, LAURA CENTRO DE INVESTIGACIONES MÉDICAS APLICADAS UNIDAD DE PROTEÓMICA, GENÓMICA Y BIOINFORMÁTICA Tel: 948194700 (5012-5013) Fax: 948194717 Email: [email protected] TENA ALDAVE, MANUEL UNIVERSIDAD DE CÓRDOBA DPTO. DE BIOQUÍMICA VEGETAL Y AGRÍCOLA Tel: 957218574
Fax: 957218592 Email: [email protected] VALLEDOR GONZÁLEZ, LUIS UNIVERSIDA DE OVIEDO DPTO. DE BIOLOGÍA DE ORGANISMOS Y SISTEMAS Tel.: 985 104834 Email: [email protected] VÁZQUEZ COBOS, JESÚS CENTRO DE BIOLOGÍA MOLECULAR SEVERO OCHOA LABORATORIO DE QUÍMICA DE PROTEÍNAS Y PROTEÓMICA Tel: 914978276 Fax: 914978087 Email: [email protected]

Boletín de la Sociedad Española de Proteómica, Nº 2,Junio 2006
116
Y, como siempre, nuestro agradecimiento a los Socios Patrocinadores de la SEProt