biología - variabilidad y adaptación de los seres vivos · 1 introducciÓn la asignatura...
TRANSCRIPT

1
INTRODUCCIÓN La asignatura Variabilidad y adaptación de los seres vivos da continuidad a los conceptos básicos de la teoría evolutiva que se introducen en el curso. La ciencia de la vida y tiene como principal propósito que el futuro docente adquiera una plataforma conceptual básica en torno a la teoría de Darwin, que le permita comprender el proceso de la evolución biológica para orientar su aprendizaje en los alumnos de la escuela secundaria. El curso se inicia con el análisis de algunos fragmentos de los trabajos originales de Darwin y Wallace. Con esta actividad se pretende que el estudiante normalista elabore sus propias interpretaciones, las exponga y contraste, tanto con las de sus compañeros, como son las que se incluyen en algunos libros de texto. Además se retoma y profundiza el estudio de la selección natural y los conceptos básicos que se asocian a ésta, como son la variabilidad, la supervivencia y la adaptación. La selección natural, piedra angular del pensamiento darviniano se presenta en dos fases: en la primera se abordan aspectos acerca de la variabilidad y su origen. En la segunda se establece que el cambio en las especies es el producto de la adaptación y supervivencia diferenciada en la variedad de ambientes que pueden ocupar. El desarrollo del tema inicia con una actividad en la que se recuperan las ideas previas de los estudiantes acerca de la variabilidad y su origen, para pasar después a un análisis de los capítulos “La variación en la naturaleza”, “La lucha por la existencia” y “Selección natural o la supervivencia de los más aptos”, del libro El origen de las especies, de Darwin. Así mismo, se analiza el concepto moderno de “selección natural” en términos genéticos y se ilustra con ejemplos como la anemia falciforme, la resistencia a la malaria, el mecanismo industrial y la resistencia de los insectos. Una vez que los estudiantes identifican el esquema evolutivo propuesto por Darwin, se discute la teoría sintética de la evolución y se aclara que aun cuando existen otras propuestas, ésta se conserva como la base de toda explicación para entender el cambio en los organismos vivos y su diversidad. La última parte del curso se dedica a la identificación de las ideas equivocadas y explicaciones alternativas acerca de la evolución, que son frecuentes en el pensamiento de los alumnos en la escuela secundaria. En esta parte también se presentan algunas propuestas para favorecer la reestructuración de conocimientos y promover el cambio conceptual. también se incluyen sugerencias para desarrollar el tema de la evolución biológica en el salón de clases y los recursos didácticos que se pueden aprovechar para apoyar y mejorar la enseñanza y el aprendizaje.
RELACIÓN CON OTRAS ASIGNATURAS El presente curso se relaciona horizontal y verticalmente con otras asignaturas que se han estudiado en los semestres anteriores y las que se cursan de manera simultánea con éste. Las relaciones directas se establecen con la asignatura Introducción a la Enseñanza de: Biología que los estudiantes cursaron en el segundo semestre y con la asignatura denominada Procesos vitales: estructura y funciones de los seres vivos, que se imparten en este semestre. Durante el desarrollo de la asignatura Introducción a la Enseñanza de: Biología, cursada En el segundo semestre, los estudiantes reconocieron la importancia científica y pedagógica de los conceptos de alto poder explicativo. En este semestre se retoman dichos conceptos, indispensables en las respuestas a las interrogantes del “qué”, “cómo” y “porqué” de los seres vivos. Más adelante, en los semestres cuarto, quinto y sexto se atenderán con mayor detalle y profundidad estos temas, de manera especial en los cursos particulares de cada disciplina y en la asignatura Biología I, II y III. Otra relación directa se establece con Procesos Vitales: Estructura y Funciones de los Seres Vivos, que inicia su estudio tomando como referencia el cuerpo humano, para después identificar el paralelismo evolutivo, estructural y funcional de la vida.

2
ORIENTACIONES DIDÁCTICAS GENERALES A continuación se enuncian algunas líneas de trabajo que sería conveniente desarrollar a lo largo del curso: 1. Lograr que el conocimiento de los fines y el contenido de este programa sea compartido por
docentes y estudiantes. Será provechoso que al iniciarse el curso, el docente y el grupo analicen conjuntamente el programa, para dejar claros sus propósitos formativos, la secuencia de sus componentes y el tipo de trabajo que se espera de cada quien. Durante el curso, cuando sea necesario, deberá regresarse a la lectura del programa para precisar porqué y para qué trabajar determinados contenidos y actividades.
2. Aprovechar los conocimientos y experiencias previas del alumnado a fin de incorporarlos al
proceso de planeación de la enseñanza y evaluación de los logros del aprendizaje. 3. Fomentar la convicción de la existencia de múltiples manifestaciones naturales que ofrecen
diversas oportunidades para el aprendizaje, para lo cual es necesario que los propios normalistas recuperen y ejerciten sus habilidades de observación y exploración, tanto en su propio cuerpo como en su entorno inmediato. También es importante que reconozcan los recursos y materiales de apoyo didáctico que por su disponibilidad en la región pueden aprovecharse mejor.
4. Asegurar una lectura comprensiva de la bibliografía básica y vincular las ideas que en ella se
presentan, con las actividades que se realicen en clase y con las actividades de observación y práctica que realizan los estudiantes en las escuelas secundarias. Debe evitarse el riesgo común de que el material de lectura sea visto como algo separado del trabajo aplicado, que se lee por obligación y está sujeto a formas poco eficaces de control y asumir, en cambio, que la mejor forma de demostrar una buena lectura es incorporar su contenido al análisis, la discusión y la actividad práctica.
Si el maestro advierte que algunos alumnos muestran dificultades en el manejo de la bibliografía, puede promover la formación de círculos de estudio que funcionen temporal o continuamente, solicitando la colaboración de los alumnos más adelantados.
5. Incluir, en el programa del trabajo del grupo, actividades en los cuales los estudiantes lleven a la
práctica las observaciones y la indagación propuesta para los alumnos de la escuela secundaria en temas especialmente relevantes, los programas, el libro para el maestro y los libros de texto. Ello permitirá a los futuros maestros experimentar situaciones que vivirán sus alumnos, revisar con detenimiento los materiales didácticos para su aprovechamiento eficaz y poder anticipar algunos retos y dificultades pedagógicos que enfrentarán en su vida profesional.
6. Promover sistemáticamente la observación y el acercamiento de los estudiantes normalistas con
los adolescentes de la escuela secundaria, a propósito del conocimiento de la naturaleza y el aprendizaje de la biología. Una oportunidad de hacerlo sistemáticamente la ofrece la asignatura Observación y Práctica Docente I; sin embargo, se deberá alentar a los estudiantes a buscar y aprovechar todas las ocasiones informales para hacerlo, será con grupos escolares a los que tengan acceso o con adolescentes de su entorno familiar y de residencia. La familiarización con las formas de percepción y reflexión de los adolescentes, de sus reacciones ante estímulos cognitivos que poseen un propósito claro, permitirá a los estudiantes desarrollar su sensibilidad y su capacidad de empatía hacia la perspectiva desde la cual los adolescentes miran y tratan de dar sentido al mundo que les rodea.
7. Realizar actividades complementarias de estudio para fortalecer la formación disciplinaria básica
de la biología. El maestro y los estudiantes deberán estar atentos a la detección oportuna de deficiencias y vacíos que pueden existir en la formación individual. En esos casos, el docente habrá orientar el estudio y consulta de la bibliografía pertinente y accesible en el acervo de la biblioteca de la escuela.
Así mismo, es recomendable aprovechar los audios cintas, el material video grabado y los programas de informática educativa disponibles en la biblioteca de la escuela normal y en los Centros de Maestros.

3
8. Establecer un adecuado equilibrio entre el trabajo de los alumnos, tanto individual como en equipo. Es claro que númerosas actividades de aprendizaje deben realizarse individualmente, en tanto que otras se benefician con el esfuerzo de un grupo de trabajo. En este último caso, deben observarse ciertas normas mínimas que aseguren la eficacia de esta modalidad de organización didáctica: la planeación clara del trabajo, la distribución equitativa de las tareas y el carácter realmente colectivo del análisis, la discusión, la elaboración del resultado final del trabajo y la evaluación. Estas normas son útiles porque evitarán una frecuente deformación del trabajo de equipo, que fracciona temas de aprendizaje, no permite a los estudiantes visualizar los contenidos en su conjunto y oculta desequilibrios injustos en el esfuerzo realizado por cada alumno. Se sugiere establecer como criterio que los equipos se integren con cinco alumnos como máximo..
9. Propiciar la redacción de notas de lectura, registros de observación, así como el diseño y
elaboración de actividades y materiales didácticos para el desarrollo de los temas que integran los programas de biología en la escuela secundaria. En este sentido es conveniente que cada estudiante integre a lo largo del curso una carpeta personal con los productos del aprendizaje, útil para el ordenamiento y la clasificación de su trabajo, para consultar durante los siguientes semestres, en su futuro trabajo profesional y, eventualmente, como elemento para la evaluación.
SUGERENCIAS PARA LA EVALUACIÓN Los criterios y procedimientos que se definan para evaluar habilidades, valores, actitudes y conocimientos adquiridos por los estudiantes durante el estudio de los temas del curso, deben ser congruentes con los propósitos y las orientaciones didácticas señaladas. Es necesario tener en cuenta que la evaluación, entendida como proceso permanente, no sólo permite identificar los avances y las dificultades en el aprendizaje de los estudiantes, sino también aporta información que el maestro puede aprovechar para tomar decisiones que contribuyan a mejorar sus formas de enseñanza. Para que los estudiantes tomen conciencia de los compromisos y tareas que le corresponde asumir, es conveniente que al iniciar el curso acuerden con el maestro los criterios y procedimientos que se aplicarán para evaluar. De esta manera tendrán los elementos básicos para reconocer aquellos campos específicos en los que requiere fortalecer su formación profesional. Las características de este curso y el tipo de actividades a realizar requieren de prácticas de evaluación diversas que evidencien los conocimientos que se adquieren de las actitudes, las habilidades y los valores que los alumnos manifiestan ante el trabajo individual y colectivo, hacia los adolescentes y hacia la naturaleza. Para evaluar, deben observarse y registrarse sistemáticamente las actitudes, las habilidades y los valores que manifieste cada alumno durante el curso, para hacer comparaciones e identificar sus avances. También debe aprovecharse su participación en la clase, los textos escritos y las indagaciones que realicen. En este caso, la evaluación no requiere de acciones ni productos distintos de los generados en los procesos mismos de enseñar y aprender. Cuando se considere necesario que los alumnos muestren sus niveles de logro por medio de un desempeño destinado especialmente a la evaluación, los instrumentos seleccionados deben: plantear retos para que los estudiantes apliquen su capacidad de análisis, interpretación, juicio crítico, comprensión, relación, síntesis, argumentación y toma de decisiones; y proporcionar información sobre rasgos como los que se enuncian enseguida. • El interés que muestran los estudiantes por acercarse al conocimiento científico. • La comprensión de las intenciones educativas de la enseñanza de la biología en la escuela
secundaria, a partir del análisis de los contenidos propuestos en los programas de estudio de este nivel.
• La habilidad para vincular las elaboraciones teóricas con el análisis de las situaciones educativas relacionadas con la enseñanza y el aprendizaje de la biología.

4
• La capacidad para diseñar, mediante el conocimiento y uso eficaz de los libros de texto y otros recursos educativos y del medio, estrategias didácticas que estimulen en los adolescentes las habilidades y actitudes propias de la indagación y del pensamiento científicos.
Para lograr lo anterior, se sugiere tomar como base las recomendaciones de evaluación de los libros para el maestro de Biología, Física y Química. Una combinación de éstas podrá ayudar a utilizar los instrumentos adecuados para cada situación que se necesite evaluar.
PROPÓSITOS GENERALES El curso de variabilidad y adaptación de los seres vivos corresponde al séptimo semestre del Plan de Estudios de la Licenciatura en Educación Secundaria y tiene como propósitos generales que los alumnos normalistas: 1. Que el futuro docente adquiera una plataforma conceptual con la importancia de la evolución. 2. Comprender el esquema evolutivo de Darwin tomando en cuenta la selección natural y otras
teorías sobre el cambio evolutivo. 3. Identifique las ideas y los errores conceptuales más comunes relativos a la evolución, para que
adquieran las capacidades y elaboren propuestas didácticas para fomentar habilidades, aptitudes, valores y la apropiación de conocimientos pertinentes en los alumnos de educación secundaria acerca de la evolución, selección natural, variabilidad, supervivencia, adaptación y otras teorías sobre el cambio evolutivo..
ORGANIZACIÓN POR BLOQUES El programa del curso esta organizado en tres bloques temáticos. Los bloques, sus propósitos y actividades sugeridas son las siguientes.
BLOQUE I
IMPORTANCIA DE LA EVOLUCIÓN TEMAS: Tema 1. Darwin y Wallace. Tema 2. La teoría sintética de la evolución. Este bloque se inicia con actividades en algunos fragmentos de los trabajos originales de Darwin y Wallace. Con esta actividad se pretende que el estudiante normalista elabore sus propias interpretaciones, las exponga y contraste, tanto con las de sus compañeros como con las que se incluyen en algunos libros de texto. Se aprovecha la lectura de Ernest Mayr “La evolución para diferenciar la evolución cósmica y la evolución biológica”, así como también la importancia y los principios que toma en cuenta la evolución y como es que se publica “El origen de las especies”. Aquí los estudiantes normalistas deberán valorar los trabajos que se desarrollaron por Darwin, Wallace y Malthus y hacer comparaciones entre la capacidad que cada uno tiene para el desarrollo de la investigación científica. También se aprovecha el video “los monstruos emergen”, de la serie “Los dinosaurios” con la finalidad de convertirse en un testigo más de la primera reacción de un mundo escéptico ante el conocimiento de que el subsuelo de la tierra esta poblado de restos de gigantes desconocidos de un pasado remoto. BIBLIOGRAFÍA BÁSICA:

5
• Guillén Fedro (1998), El científico que creía en fantasmas, México, Pangea. • Mayr Ernest (1978), “La evolución”, en investigación y Ciencia, núm. 26, New Cork, Scientific
American, pp. 139-149. • Material de Biología, Barajas Esperanza (1984). En bios vida. México. Editorial Herrero S.A. y
Meter Alexander y otros (1992). En Biología New Jersey. Prentice Hall. • Video: Los dinosaurios, “Los monstruos emergen”. ACTIVIDADES SUGERIDAS: Tema 1. Darwin y Wallace. 1. Leer la lectura “el científico que creía en los fantasmas” Alfred Russel, Wallace de Fedro Carlos
Guillén y realizar las siguientes actividades: • ¿Qué actividades realizaba Wallace ? ¿Cuál se consideró más importante?. • ¿Cuáles fueron las ideas de Malthus que impactaron a Wallace? • ¿Cómo tituló el ensayo que escribió todavía enfermo? • Escriba brevemente la historia de la carta que envió Wallace a Darwin. • En que fecha envió la carta Wallace a Darwin? • ¿Cuándo se publicó “El origen de las especies por Darwin? • ¿Por qué se le consideró a Wallace como el espiritista científico, el biogeógrafo y el luchador
social? • Describa brevemente los dos textos de Wallace que se incluyen en estas lecturas.
2. De acuerdo a la lectura “ La evolución” de Ernest Mayr conteste las siguientes preguntas:
• ¿Cuál es la diferencia entre evolución cósmica y evolución biológica? • ¿Qué es la evolución biológica? • ¿Quién propuso la primer teoría sobre la evolución y en que año? • Señalar los cuatro principios que tomó en cuenta para explicar el curso particular de la
evolución? • En 1838 ¿Cuál era el mecanismo que podría explicar la evolución? • ¿Cuándo se publicó la teoría completa de Darwin “El origen de las especies? • Señale los cuatro postulados que destaca Darwin en su obra y de los cuáles dos están de
acuerdo con Lamarck? • Mencione las dos etapas en las que se basa la selección como un proceso según Darwin. • Según Malthus en su lectura hace reflexionar a Darwin sobre “que descendientes tienen una
probabilidad para sobrevivir?” señálelo. • Señale 3 avances que hubo para abandonar el saltacionismo. • Una vez alcanzada la nueva síntesis en la década de 1930-1940 algunos discrepantes de la
evolución se preguntaron si no había llegado ya la hora final de la investigación en este sentido. Según la biología evolutiva ¿Cuáles serían los nuevos problemas a tratar?
• Comparar y complementar las respuestas a nivel de grupo. 3. De acuerdo a materiales de Biología proporcionados conteste brevemente:
• ¿Cuáles son las dos ideas principales en la hipótesis de Lamarck sobre la evolución? • ¿Porqué los científicos rechazan la hipótesis de Lamarck sobre cómo ocurre la evolución? • Resuma los puntos principales de la teoría de Darwin de la evolución por selección natural. • ¿En qué se diferencian las ideas de Lamarck sobre la evolución de las ideas de Darwin? • ¿Cómo las observaciones de poblaciones de alevilla moteadas produjeron evidencia directa
sobre la evolución? 4. Analizar el video “Los monstruos emergen” de la serie los dinosaurios y conteste las siguientes
preguntas: • ¿Qué le llamó la atención? • ¿Cuánto tiempo los dinosaurios fueron los amos del planeta? • ¿Cómo los dinosaurios vuelven a la vida?¿En qué siglo comienza la historia de los dinosaurios
en la edad moderna? • ¿ Qué pieza fue de las primeras encontradas y por quién?. Después de 25 años se reclama la
pieza encontrada, ¿de qué parte? • ¿Cuándo se produce el primer hallazgo de fósil de dinosaurio y en donde?.

6
• ¿Qué se encuentra como evidencia incrustada en una roca? • ¿Por que se dice que en 1970 todo cambia? • ¿Crees que los dinosaurios han desaparecido y se fueron a volar? • ¿Qué semejanza habría con los pájaros? • Conclusión en el grupo.
Tema 2. La teoría sintética de la evolución. 1. ¿Qué otros nombres recibe la teoría sintética de la evolución? 2. ¿Qué es la teoría sintética de la evolución? 3. ¿Cuáles son los principales fundamentos de la teoría sintética? 4. ¿A quién se debe esta teoría? 5. ¿Qué son las mutaciones? 6. ¿Cuáles son los tipos de mutaciones y cómo se explican? 7. ¿Cuáles fueron las aportaciones de otras ciencias a la teoría de la evolución? 8. ¿Cómo es el panorama actual en biología? 9. ¿Cuáles son los resultados de las investigaciones que aportaron Eric, Watson, Wilson, Lorenz y
algunos biólogos mexicanos como Alfonso Herrera, Enrique Beltrán y Arturo Gómez Pompa?
BLOQUE II EL ESQUEMA EVOLUTIVO DE DARWIN
TEMAS: Tema 1. Selección natural: variabilidad, supervivencia y adaptación. Otras teorías sobre el cambio evolutivo. En el bloque número II se brinda información de consulta para que el estudiante normalista nuevamente retome y haga las investigaciones sobre la evolución. Se pretende que el estudiante normalista realice un breve informe sobre el viaje de Darwin. Se presenta una actividad para reconocer la selección natural por medio de un experimento. BIBLIOGRAFÍA BÁSICA: • Material de Biología, Barajas, Esperanza (1984). En bios vida. México. Editorial Herrero S. A. y
Meter Alexander y otros (1992). En Biología. New Jersey. Prentice Hall. • Darwin, Charles (1985), “La variación en la naturaleza”, la lucha por la existencia”, “La selección
natural, o la supervivencia de los más aptos”, en El origen de las especies, México Consejo Nacional de la Ciencia y la Tecnología, pp. 125-135, 136-155, 156-203.
• Reeves, Hubert y otros (1999), La más bella historia del mundo, México, Andrés Bello/SEP (Biblioteca para la actualización del maestro).
• “La adaptación” de Richard C. Lewontain.

7
ACTIVIDADES SUGERIDAS: Tema 1. Selección natural: variabilidad, supervivencia y adaptación, otras teorías sobre el cambio evolutivo. 1. De la información que se le proporciona investigar lo siguiente:
• ¿Qué es la evolución? • En el proceso de evolución ¿Cómo se explican los dos tipos de materia? Y ¿Cómo se explican
las dos etapas en la evolución de materia orgánica? • Señalar en que consisten las primeras teorías de la evolución. • ¿Cómo se originaron las primeras teorías de la evolución? Mencione los autores. • ¿Cómo explica Cubier la presencia de restos fósiles en diferentes partes de la tierra? • ¿Cómo explica Lamarck la evolución de los seres vivos? • ¿Qué señala la teoría de la evolución de Lamarck? • ¿Cuáles son las dos ideas principales en la hipótesis de Lamarck sobre la evolución? • ¿Por qué los científicos rechazan la hipótesis de Lamarck sobre la evolución? • Resuma los puntos principales de la teoría de Darwin de la evolución por selección natural
(5) y explica cada uno de ellos. • ¿Cómo las observaciones de poblaciones de alevillas moteadas proveyeron evidencia directa
sobre la evolución? • Realizar un debate en el grupo para hacer la conclusión final.
2. Prepara un informe acerca del viaje de Darwin que duró 5 años en el
• H. M. S. Beagle. Incluya un mapa del viaje. • ¿Cuáles fueron las consecuencias del viaje del Beagle? • ¿Cómo influyeron con Darwin: Malthus y Wallace?
3. ¿Cómo puede representar la selección natural? Realice la actividad que se propone a
continuación. ¿Cómo puede representarse la selección natural? Objetivos • Demostrarás cómo la coloración puede ser un factor en la selección natural. • Predecirás qué individuos sobrevivirán en una población de animales. • Predecirás los cambios en una población a través del tiempo. Materiales (para grupos de 4) • Dos hojas de papel periódico. • Una hoja de papel de construcción negro. • Una de papel de construcción blanco. • Tijeras. • Un envase grande. Procedimiento A. Corta una hoja del periódico que sea del mismo tamaño y forma del papel de construcción y
dóblala en 64 rectángulos y recórtalos. Cada uno medirá, aproximadamente, 2.7 cm. por 3.3 cm. De la misma forma, dobla y luego recorta el papel de construcción negro en 64 rectángulos. Haz lo mismo con el papel de construcción blanco.
B. Los rectángulos blancos, negros y de papel de periódico representan animales en un ambiente. La hoja de papel de periódico que no se recortó representa el ambiente.
C. Coloca todos los rectángulos en el envase y agítalo bien. Deja caer todos los rectángulos en la hoja de papel periódico, de manera que queden dispersos.
D. Tú y tus compañeros de laboratorio van a ser los “depredadores”. Ustedes atacarán a los animales que estén en el ambiente de la hoja de periódico. Cada persona en tu grupo debe “capturar” cinco animales de papel de construcción o de periódico de la siguiente forma: Mira hacia un lado un momento. Luego, vírate y coge un animal lo más rápidamente que puedas. Repite el procedimiento hasta que hayas capturado cinco animales.
E. En un papel copia la tabla de abajo y registra el número de cada tipo de animal de papel que capturaste.
F. Calcula el número de cada tipo de animal que sobrevivió (recuerda que empezaste con 64 de cada color).
G. Compara los datos que tu y tu grupo han recogido con los de los del resto de la clase. Calcula el número total de sobrevivientes de cada tipo de animal.

8
Preguntas y conclusiones 1. De los datos que has recogido de la clase completa, ¿cuáles de los animales de papel
sobrevivieron en mayor número? ¿Por qué crees que estos animales sobrevivieron? 2. Predice el tipo de cambio que es probable que puedas encontrar en esta población de
animales de papel a través de un periodo de tiempo. 3. Imagínate que el ambiente se hiciera, gradualmente, más oscuro. ¿Cómo podría este cambio
afectar el número y los tipos de animales en la población? 4. Describe una situación en el mundo real que ilustre lo que has representado.
ESTUDIANTE ANIMALES DE PAPEL
BLANCO ANIMALES DE PAPEL
NEGRO ANIMALES DE
PAPEL PERIODICO
4. De la información proporcionada:
• Prepare un breve resumen sobre Darwin y la selección natural. • ¿Qué es la variabilidad? y ¿Cuáles son los mecanismos que la producen? • ¿Qué es la adaptación? y explique los tres tipos de adaptación. • ¿Qué es la supervivencia? • Obtener las conclusiones del grupo.
5. Aplique sus conocimientos y conteste ¿Qué pasos del método científico empleó Darwin para la
elaboración de su teoría de la evolución? Seleccionado las palabras: análisis de datos, adaptación, recolección y observación; para complementar los siguientes párrafos: • Durante el viaje en Beagle, Darwin _________________________________ los
______________________________ muestras de rocas y restos fósiles. Al y _______________________ sus observaciones. Con base en lo anterior elaboró los postulados de su teoría.
• ¿Por qué a lo largo de la historia de la tierra han desaparecido y aparecido muchas especies de seres vivos______________________.
• Los cambios ambientales crean nuevas situaciones para los organismos, los cuáles deben _________________las nuevas condiciones.
• Los organismos mejor, _________________ al medio son los que lograrán tener descendencia que conservará la especie.
6. Lea nuevamente el texto referente al viaje de Darwin y por equipo contestar y discutir lo
siguiente. • ¿Qué elementos utilizó Darwin para postular la teoría de la evolución? • ¿Cuál es la importancia de la recolección de datos en la elaboración de la teoría? • ¿Por qué cree que los fósiles aportan datos para el estudio de la evolución de los
organismos? • Registrar en un cuaderno las conclusiones a las que llegaron.
7. Darwin en la isla Isabela. Lea con atención el siguiente texto escrito por Darwin durante en su
estancia en la Isla Isabela, del archipiélago de Galápagos.
“Es muy interesante contemplar una colina llena de maleza, cubierta de muchas plantas con pájaros que cantan entre los arbustos con variedades de insectos volando por el aire, gusanos arrastrándose por el suelo húmedo. Mientras reflexionamos que estas formas tan cuidadosamente construidas, que dependen las unas de las otras de manera compleja todas ellas han sido producidas por leyes son la del crecimiento con reproducción, de la herencia, considerada en la reproducción, la variabilidad, a partir de la acción indirecta y directa de las condiciones de vida externa y del uso y desuso. La de la progresión geométrica de crecimiento, capaz de provocar la lucha por la vida y en consecuencia la selección natural, que comparta la divergencia de caracteres y la extinción de las formas menos mejoradas. De este modo, de la guerra de la naturaleza, del hombre y de la

9
muerte se sigue directamente el objetivo supremo, es decir la producción de animales superiores. Es grande esta concepción de que la vida fue originalmente creada en unas pocas formas o en una sola; y que mientras este planeta ha ido girando, evolucionaron y evolucionarán a partir de un inicio simple, infinidad de formas a cual más bella y maravillosa”.
Charles Darwin. El origen de las especies. Septiembre 20 de 1835.
• Copie el texto en su cuaderno y marque con color las ideas que forman parte de la teoría de
la evolución. • Identifique en el texto la parte Darwin hace uso de la observación y en el análisis. • Conteste en equipo.
- ¿Qué es lo que observa Darwin y como lo interpreta? - ¿Qué leyes rigen la evolución de las especies según Darwin ¿ - ¿Qué diferencias existen entre el concepto evolutivo de Darwin y el de Lamarck?
8. Aplique sus conocimientos y conteste lo siguiente, puede ser en equipo:
• Use sus conocimientos de genética para explicar ¿Por qué las características adquiridas no son pasadas a la progenie?
• El uso de insecticidas a veces lleva el desarrollo de poblaciones resistentes a estos. Explique ¿Qué está ocurriendo en términos de la selección natural y de la evolución?
• ¿ Qué tipo de evidencia fósil puede apoyar la hipótesis del equilibrio puntual? • ¿Cree que hoy en día ocurra alguna evolución? Dé razones para apoyar su respuesta. • La mayoría de los países industriales esta tratando de reducir la contaminación ambiental.
Describa lo que podría ocurrirles a las poblaciones de alevillas moteadas si este esfuerzo tiene éxito en Inglaterra.
• Las variadas razas de perros pertenecen a la misma especie. Prepare un “árbol de especies” para demostrar cómo están relacionadas las diversas razas.
• Seleccione una especie o vegetal observe el organismo y busque información acerca de él. Prepare una lista de todas las adaptaciones del organismo. Identificando cada en cuanto a adaptación morfológica o de comportamiento. Use sus conocimientos de selección natural para explicar cómo pudo haber evolucionado cada adaptación.
• Pídale a un criador de animales o a un criador de plantas que describa cómo una mutación deseable se puede preservar a través de la selección artificial.
9. De acuerdo a la lectura “La adaptación” de Richard. Lewontin haga un breve resumen y
cuestione preguntas de una respuesta adecuada. 10. Investigue otras teorías sobre el cambio evolutivo. 11. Analiza lo siguiente:
¿A quién no le gustaría sentarse a charlar con un gran científico, preguntarle de los grandes enigmas de la existencia y escuchar sus respuestas en palabras usuales, amenas y con base a los más recientes conocimientos de la ciencia? Esta charla se realiza a través de la lectura “La explosión de las especies” del libro “La más bella historia del mundo” donde Dominique Simonnet plática con tres expertos de reconocimiento mundial un astrofísico “Reeves”, un divulgador de las ciencias (Roshay), y un paleontólogo (Coppens). “La más bella historia del mundo” ofrece también en esta manera de platicar de las ciencias, un atractivo modelo de cómo el maestro puede transmitir a sus alumnos conocimientos en términos que además de comprensibles estimula el interés de los estudiantes.
• Leer “la explosión de las especies de Reeves y otros y seleccione 6 relatos que le llamen la
atención y haga una comparación en equipo y aplíquelo en su vida diaria. • Hacer un ensayo sobre el material que ha seleccionado.
BLOQUE III MÉTODOS PARA LA ENSEÑANZA Y EL APRENDIZAJE DEL

10
TEMA DE EVOLUCIÓN EN LA ESCUELA SECUNDARIA TEMAS: Ideas de los alumnos en torno a la evolución biológica. Recursos materiales para la enseñanza y el aprendizaje de la evolución en el aula. En el bloque III presenta una serie de lecturas para realizar la unidad “La evolución” de tal manera que se relacionan una con otra para aplicar una serie de conocimientos, hábitos y actitudes los que se enriquecerán con las experiencias de los estudiantes. Se presenta también una estrategia didáctica titulada “El cambio conceptual y metodológico en la enseñanza y el aprendizaje de la evolución de los seres vivos” con la finalidad de que los estudiantes normalistas reconozcan lo fácil que es llevar un trabajo organizado como un ejemplo claro, dándoles libertad para realizar algo semejante lo cual se invita a la participación socializadora por lo que la actividad puede realizarse por equipo. BIBLIOGRAFÍA BÁSICA: • Durand Leticia Smith, “La evolución”, Guillén Fedro Carlos, “Problemas asociados a la enseñanza
de la evolución en la escuela secundaria: algunas sugerencias”, “Darwin y Wallace”; Sarukhán José, “Las musas de Darwin”; Leakey Richard E., “ Introducción a el origen de las especies” en la enseñanza de la Biología en la escuela secundaria, SEP, pp. 122-129, 159-170, 283-287, 74-81, 248-274.
• Gené A. (1991), “Cambio conceptual y metodológico en la enseñanza y el aprendizaje de la evolución de los seres vivos”, en Enseñanza de las ciencias, España, 9 (1), pp. 22-27.
ACTIVIDADES SUGERIDAS: Tema 1. Ideas de los alumnos en torno a la evolución biológica. Recursos materiales para la enseñanza y el aprendizaje de la evolución en el aula. 1. De acuerdo a la lectura “la evolución” de Leticia Durond Smith contestar de manera individual:
• ¿Por qué se señala que el hombre ha dependido siempre de su relación con otros animales? • En 1809 ¿qué propuso Jean B. Lamarck con respecto a los seres vivos? • Cuáles son los 4 elementos que se mezclaron para dar origen a la vida según Anaximandro y
Empedocles (600-500 AC.) • Cómo se presentaban los trabajos de los naturalistas. • Cuáles fueron las primeras observaciones de Darwin en su visita a las Islas Galápagos, • Mencionar las condiciones para que se desarrolle un proceso de evolución por selección
natural según Darwin. • Mencionar dos aspectos importantes en la teoría Darvinista. • Por qué Darwin no se explicaba cual era el mecanismo que permitía la transmisión de
caracteres de una generación a otra. • Quién da la solución al problema de los rasgos característicos de una generación a otra y
como lo explicaba. • Qué adelantos en Biología produjeron la necesidad de reconstruir la teoría Darvinista • Cuáles son los tres hechos que según el neodarwinismo y el darwinismo sostienen la
existencia de la selección natural como fuerza evolutiva. • A que se llama deriva génica. • Qué procesos se mencionan en la selección como organismos “exitosos” o “fracasados”. • ¿Cuáles son las principales críticas al neodarwinismo? • Obtener conclusiones a nivel grupal.

11
2. Leer la lectura “Problemas asociados a la enseñanza de la evolución en la enseñanza de la evolución en la escuela secundaria: algunas sugerencias” de Fedro Carlos Guillén. • Elabore en su cuaderno de conclusiones una tabla enlistando las ideas equivocadas de los
jóvenes sobre la evolución y explicando porque son erróneas.
Ideas de los jóvenes ¿Por qué está equivocada?
• A continuación haga un recuentro de sus experiencias en el salón de clases y enliste en su
cuaderno de conclusiones las ideas erróneas sobre la evolución que más frecuentemente ha escuchado entre sus alumnos.
• Para cada uno de los siguientes párrafos explique en su cuaderno de conclusiones:¿Qué autor propuso esa idea?, ¿Qué ideas semejantes a oído de sus alumnos u otras personas¿ y ¿Qué elementos históricos influyeron, de acuerdo con su criterio personal, en la postulación de esa teoría para facilitar la resolución de esta actividad? Analizar el ejemplo inicial. I. “El hombre es un ser superior, es capaz de pensar y resolver problemas, y ha
evolucionado mucho más que otros animales” a) La idea del hombre como un ser superior prevalece desde los griegos Anaximandro y
Aristóteles, quienes reconocían seres superiores e inferiores. b) He leído ideas semejantes, como por ejemplo de algunas personas que se oponían a
Darwin diciendo que era imposible que estuviéramos emparentados con los monos, ya que el hombre era un ser mucho más capaz y civilizado.
c) Creo que, dado que los griegos observaban al mundo con sus propios sentidos, es decir, que no contaban con instrumentos sofisticados que les permitieran observar detalles de la composición y funcionamiento de los seres vivos, lo que saltaba a la vista era la capacidad del hombre para pensar y comunicarse, lo que les hizo creer en la existencia de seres superiores e inferiores.
II. “Durante su vida los organismos adquieren características que les ayudan a sobrevivir”. III. Los organismos tienden hacia una mayor complejidad y protección”.
3. Darwin y la selección natural. Leer la lectura “Darwin y Wallace” de Fedro Carlos Guillén y
escriba en su cuaderno de conclusiones, el interés que este tipo de texto, donde se presentan aspectos de la vida cotidiana de los científicos, puede generar en sus alumnos, y la utilidad que puede tener en sus clases. También conteste las siguientes preguntas: • ¿Qué hubiera hecho si fuera Darwin? • ¿Cree que resolvió bien el problema? • ¿Cree que la condición de científico libera a las personas de los problemas y conflictos de
cualquier ser humano? • ¿Por qué existe el mito de los científicos como personas alejadas de la realidad, que viven
aisladas en sus laboratorios? • ¿Cómo podemos modificar esta concepción en nuestros alumnos?
4. El propósito es conocer la influencia que tuvieron Malthus y Wallace en el desarrollo del trabajo
de Darwin. • En el tema “La evolución es importante que los alumnos comprendan la gran trascendencia
que tuvieron las ideas de Malthus y Wallace en el pensamiento de Darwin y en el desarrollo de su teoría evolutiva. Para esto se sugiere pedirles que escriban un cuento con dos finales, en uno deben contar que paso cuando Darwin recibió la carta de Wallace y cuando leyó el libro de Malthus. “Ensayo sobre el principio de la población”, y en el otro cuento lo que hubiera pasado si Darwin no hubiera tenido contacto con estas personas. El maestro puede apoyarse en los siguientes datos biográficos para proporcionar información a los alumnos:
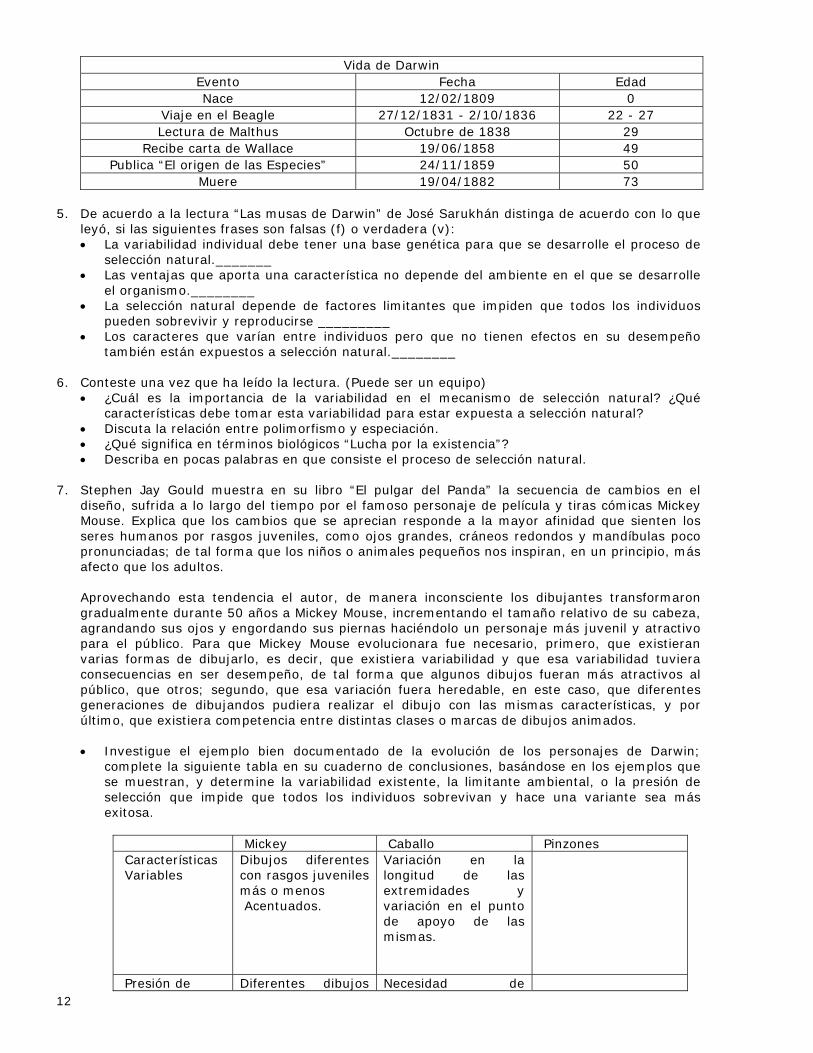
12
Vida de Darwin Evento Fecha Edad Nace 12/02/1809 0
Viaje en el Beagle 27/12/1831 - 2/10/1836 22 - 27 Lectura de Malthus Octubre de 1838 29
Recibe carta de Wallace 19/06/1858 49 Publica “El origen de las Especies” 24/11/1859 50
Muere 19/04/1882 73 5. De acuerdo a la lectura “Las musas de Darwin” de José Sarukhán distinga de acuerdo con lo que
leyó, si las siguientes frases son falsas (f) o verdadera (v): • La variabilidad individual debe tener una base genética para que se desarrolle el proceso de
selección natural._______ • Las ventajas que aporta una característica no depende del ambiente en el que se desarrolle
el organismo.________ • La selección natural depende de factores limitantes que impiden que todos los individuos
pueden sobrevivir y reproducirse _________ • Los caracteres que varían entre individuos pero que no tienen efectos en su desempeño
también están expuestos a selección natural.________ 6. Conteste una vez que ha leído la lectura. (Puede ser un equipo)
• ¿Cuál es la importancia de la variabilidad en el mecanismo de selección natural? ¿Qué características debe tomar esta variabilidad para estar expuesta a selección natural?
• Discuta la relación entre polimorfismo y especiación. • ¿Qué significa en términos biológicos “Lucha por la existencia”? • Describa en pocas palabras en que consiste el proceso de selección natural.
7. Stephen Jay Gould muestra en su libro “El pulgar del Panda” la secuencia de cambios en el
diseño, sufrida a lo largo del tiempo por el famoso personaje de película y tiras cómicas Mickey Mouse. Explica que los cambios que se aprecian responde a la mayor afinidad que sienten los seres humanos por rasgos juveniles, como ojos grandes, cráneos redondos y mandíbulas poco pronunciadas; de tal forma que los niños o animales pequeños nos inspiran, en un principio, más afecto que los adultos.
Aprovechando esta tendencia el autor, de manera inconsciente los dibujantes transformaron gradualmente durante 50 años a Mickey Mouse, incrementando el tamaño relativo de su cabeza, agrandando sus ojos y engordando sus piernas haciéndolo un personaje más juvenil y atractivo para el público. Para que Mickey Mouse evolucionara fue necesario, primero, que existieran varias formas de dibujarlo, es decir, que existiera variabilidad y que esa variabilidad tuviera consecuencias en ser desempeño, de tal forma que algunos dibujos fueran más atractivos al público, que otros; segundo, que esa variación fuera heredable, en este caso, que diferentes generaciones de dibujandos pudiera realizar el dibujo con las mismas características, y por último, que existiera competencia entre distintas clases o marcas de dibujos animados.
• Investigue el ejemplo bien documentado de la evolución de los personajes de Darwin;
complete la siguiente tabla en su cuaderno de conclusiones, basándose en los ejemplos que se muestran, y determine la variabilidad existente, la limitante ambiental, o la presión de selección que impide que todos los individuos sobrevivan y hace una variante sea más exitosa.
Mickey Caballo Pinzones Características Variables
Dibujos diferentes con rasgos juveniles más o menos Acentuados.
Variación en la longitud de las extremidades y variación en el punto de apoyo de las mismas.
Presión de Diferentes dibujos Necesidad de

13
selección animados Compiten por la audiencia, Aquellos que Agraden más al Público venden Más.
incrementar la velocidad de carrera. El hecho de que sus Extremidades se Apoyan en el 3er. Dedo y sean más largas, incrementa la velocidad de los movimientos
8. Lea el texto “Introducción a el origen de las especies”, escrito por Richard Leakey, que resume el
trabajo de Darwin y las dificultades que enfrentó para lograr la aceptación de su teoría. En su texto, R. Leakey menciona la aportación a la teoría evolutiva de muchos autores que no hemos mencionado en el transcurso de este proyecto pero que no por eso dejar de ser muy importantes. Para no olvidar la idea de estas personas, explique a continuación, en cuaderno de conclusiones, la contribución de cada uno de ellos: a) Charles Lyell. b) August Weisman. c) Alfred Rusell Wallace. d) Thomas Huxley. e) Thomas Malthus.
9. Imagine que existe una población de ratones en la parte medio de una montaña de gran altura,
donde las zonas bajas se caracterizan por el clima caluroso abundantes pastos, y las zonas altas por un clima muy frío, con nieve y poca vegetación, manteniéndose la zona central con un tipo de clima y vegetación intermedios. Si en nuestra población existe variación en torno al grosor de la piel, el color del pelo y los hábitos alimenticios, y de pronto el nivel de sobre población es el que parte de los ratones se mueve hacia las partes altas de la montaña otros a las partes bajas, y una tercera parte permanece en la región media, responda en su cuaderno de conclusiones: • ¿Qué sucedería si esas tres poblaciones se mantuvieran aislados entre sí durante un largo
período de tiempo? • ¿Qué características tendrían los ratones que habitan cada una de las regiones de la
montaña? • ¿Qué sucedería si entre la población original no existiera variación? • ¿Y qué sucedería si no se lograra un aislamiento efectivo entre las tres poblaciones? • Concluya entonces, ¿Por qué el proceso de selección natural depende de la variabilidad, la
herencia y la sobrepoblación? • ¿Por qué se pueden originar diferentes especies por selección natural?
10. Nuevas ideas sobre la selección natural: Desarrolle en su cuaderno de conclusiones una práctica
para sus alumnos utilizando como material una maqueta de plastilina, que represente un ambiente y una población de organismos. Explique las diferencias entre especiación por aislamiento y especiación de deriva génica, estableciendo en la maqueta las condiciones ambientales necesarias para que ocurran estos procesos, y detallando de que forma intervienen en estos fenómenos el azar y la selección natural.
11. Retome la lectura “Introducción a el origen de las especies” de Richard E. Leakey y responda en
su cuaderno de trabajo las siguientes preguntas: • ¿Qué es un polimorfismo y que relación guarda con la teoría neutralista? • ¿Además de la teoría neutralista y el equilibrio puntuado, que otros aspectos menciona
Leakey como puntos de discusión en Biología evolutiva? 12. A lo largo de este trabajo hemos revisado la forma en que han ido surgiendo y modificándose las
ideas sobre la evolución de los seres vivos. A manera de resumen, elabore en su cuaderno de conclusiones una cronología de estas ideas en forma de tabla, indicando la fecha en que se postuló la idea, su autor, una breve explicación de esa idea, y las pruebas que existen a su favor.
Conclusión. Como hemos visto, la teoría de la evolución se ha ido modificando a lo largo del tiempo. Sin embargo, podemos distinguir cinco etapas fundamentales:

14
a) La etapa predarwinista, en donde la mayor parte de las ideas en torno a la evolución estaba
ligada a la existencia de un creador o fuerza divina. b) La etapa darvinista, que inicia con la publicación del “El origen de las especies” por Charles
Darwin, y en la cual podemos incluir los años de polémica y cuestionamiento a los que estuvo expuesta la teoría de la selección natural, hasta su final aceptación.
c) La etapa sintética, en la década de los 30 y 40, cuñado se produce la teoría sintética de la evolución, en la que se unieron a los postulados básicos de Darwin, los conocimientos de la genética mendeliana, la paleontología y otras disciplinas.
d) La etapa del endurecimiento de la síntesis evolutiva en la que todo cambio evolutivo era explicado a partir del cambio gradual y la acción de la selección natural.
e) La etapa de crítica a la teoría sintética y propuesta alternativa, que se caracteriza por la producción de nuevos descubrimientos e idea que cuestionan el papel de la mutación, el azar, la selección natural y el cambio gradual en la evolución biológica, etapa en la cual nos encontraremos actualmente.
13. Elabore en su cuaderno de trabajo un cuadro sinóptico donde resuma los principales ideas
revisadas en este trabajo ayúdese siempre con dibujos. 14. De acuerdo al material que se proporciona sobre “El cambio conceptual y metodológico en la
enseñanza y el aprendizaje de la evolución de los seres vivos”. • Leer primeramente de manera individual haciendo un análisis lo más claro posible sobre todo
en el anexo en el cual se presentan algunas actividades del programa-guía diseñado para trabajar el tema de la evolución de los seres vivos por selección natural.
• Seleccione dos problemas que sean de su agrado diferentes a los 3 ya planteados y realice las actividades que sean necesarias tome en cuenta el seguimiento de la misma metodología para realizar su proyecto.
• Si es necesario emita una hipótesis que permita dar respuesta a la cuestión que se trate. • Se recomienda hacer el trabajo por equipo. • Haga conclusiones a nivel del grupo.
15
MATERIAL
DE
APOYO
-

16

17
LA HISTORIA DE UNA CARTA Tal como ocurre con los seres humanos, hay cartas destinadas a la gloria mientras que otras, la mayoría sufren nuestro olvido. “En primer lugar, a la pregunta de si te dejo patinar te contesto rotundamente que no. Soy demasiado celoso para permitir tal cosa. Yo no sé patinar y, aunque supiera, no tendría tiempo para acompañarte, y alguien habría de hacerlo, de modo que olvídalo”, le escribió el doctor Sigmund Freud a su novia Martha en 1885, sin sospechar que su fama como padre del psicoanálisis le jugaría la mala pasada de permitirnos al resto de los mortales la oportunidad de conocer sus limitadas normas de tolerancia.
Una carta trasciende a sus contemporáneos porque ha sido escrita por un gran hombre como freud (aunque no le gustara patinar), porque contiene ideas notables o, en ocasiones, por ambas cosas. La carta de que nos ocupamos es sin duda una de las más famosas que registra la historia de la ciencia y fue llevada al puesto de correos de Ternate –una isla de tamaño de un dedal, pérdida en el archipiélago malayo- por un hombre alto de barba crecida, el viernes 12 de marzo de 1858. El destinatario en Inglaterra era nada menos que Charles Darwin, conocido por todos nosotros como el indudable autor de teoría de la evolución.
El remitente de extraordinario documento era un joven naturalista ingles llamado Alfred Russel Wallace, que se encontraba en el archipiélago malayo desde 1854. Wallace se dedicaba a formar colecciones de aves, de reptiles y sobre todo de insectos, que luego mandaba a Europa con el propósito de comercializarlas entre los coleccionistas y los museos, y así ganarse la vida.
Alfred era un hombre trabajador, ya que en los ocho años que permaneció en Asia colecto nada menos que 125660 especímenes. Con la paciencia, el tiempo y la ociosidad suficientes podremos calcular que obtenía más de 40 ejemplares por día de trabajo, lo que no deja de ser prodigio de la recolección.
Sin embargo, Wallace tenia tiempo para otras actividades; una de ellas, quizá más importante que sus asombrosos 40 especímenes al día, era la de reflexionar sobre la manera en que las especies se originaran, sin duda el problema biológico más inquietante de su época.
Tradicionalmente se creía que las especies eran inmutables, es decir que habían sido creadas unos cuanto miles de años atrás por
un ser divino y no habían cambiado desde los tiempos de la creación. Esta era la explicación que la iglesia, y en consecuencia sus fieles, aceptaban. No era sencillo desafiar estas ideas; el mismo Darwin pasó tragos amargos cuando le explicó a su esposa Emma Wedgwood su teoría evolutiva, ya que ella era una fiel cristiana y no estaba muy segura de que las ideas de su cónyuge fueran aceptables.
No obstante, a lo largo de la historia de los seres humanos siempre han tenido el muy saludable impulso de poner en duda las verdades que se consideran establecidas. Esta época, por cierto, es una de las características más notables de la investigación científica. En el caso de la inmutabilidad de las especies no faltaron algunas voces que se alzaron cuestionando esa idea.
A finales del siglo XVIII Erasmus Darwin, abuelo de Charles, escribió un ensayo sobre la evolución. Don Erasmus no era un hombre común: tartamudeaba al hablar, fue miembro fundador de una sociedad llamada Los Lunáticos y acostumbraba escribir sus ideas científicas en formas de versos. No obstante, se anticipo a muchos en cuanto a la idea de que las especies podían evolucionar.
En 1809 el francés Jean Baptiste de Lamarck publico un libro en el que sugería que los seres vivos no eran inmutables. Planteaba que en las especies había un deseo de mejoría y que en las características que los organismos adquirían a lo largo de su existencia podían legarse a sus descendientes. Las ideas de Lamarck eran esencialmente equivocadas, pero su trabajo abrió una brecha que ya no se cerraría.
Existe la evidencia bien documentada de que antes de que Darwin publicara su libro El origen de las especies se habían producido más de veinte trabajos sobre temas evolutivos. Algunos, como vestigios de la historia natural de la creación, escrito por Robert Chambers (por cierto, de manera anónima), estaban plagados de ideas incorrectas. Chambers estaba convencido, por ejemplo, de que podía producir la vida en su laboratorio. Sus contemporáneos despedazaron sus ideas científicas sin ninguna misericordia. Sin embargo, el tema de la evolución de los organismos había salido por fin del armario y era parte de las discusiones públicas entre los científicos de la primera mitad del siglo XIX.
Podemos decir que en el momento en que Alfred Russel Wallace le puso los timbres a su carta algunos hombres de ciencia habían

18
sugerido que los organismos cambiaban en el tiempo, pero que nadie en este planeta sabía como se realizaban esos cambios.
Por lo menos eso pensaba Alfred.
Wallace sólo había visto una vez a Darwin en el museo Británico, pero sabia que investigaba las variedades de las especies y por ello le escribió algunas cartas en 1857 planteándole sus ideas. Darwin contesto que sobre la cuestión del cambio de las especies venia reflexionando desde hacia veinte años, pero que era imposible explicar su pensamiento en la extensión de una carta. Esto no era esencialmente cierto, ya que el 5 de septiembre de este mismo año le envío una carta a Asa Grey, un científico norteamericano, en la que exponía su teoría del cambio de las especies, pidiéndole que no divulgara. Parecería que Darwin no confiaba en Wallace. ¿Por qué?
Wallace siguió dándole vueltas al asunto y –según cuenta- en febrero de 1858 sufrió severos ataques de fiebre. De pronto le vino a la memoria un pasaje escrito por Thomas Robert Malthus, un economista ingles que proponía que en las poblaciones humanas los alimentos crecen con menos velocidad que la cantidad de los individuos de la población, por lo que dentro de los grupos humanos se debe generar una lucha por la existencia para hacerse de dichos recursos. Las ideas de Malthus, a pesar de que era clérigo, a veces no eran muy cristianas: proponía abolir las leyes que subsidiaban a los pobres y les recomendaba no casarse si no estaban en condiciones de mantener a una familia. Es por ello que su famoso libro Ensayo sobre el principio de la población, fue recibido con fuertes críticas en Inglaterra, sobre todo por la iglesia.
Al leer a Malthus, Wallace no penso en los pobres, sino en la capacidad de las poblaciones de crecer exponencialmente... siempre que no haya fuerzas que lo impidan.
Y entonces lo tuvo todo claro.
Wallace se dio cuenta (y cálculo) que una pareja de aves se podía multiplicar en quince años hasta un número de diez millones. ¿Por qué entonces el número de aves se mantenía constante en el tiempo y no crecían hasta límites infinitos? La respuesta que
Encontró era sencilla y genial: En las poblaciones había variaciones; algunos organismos eran más veloces, otros se ocultaban mejor en su medio. Estas variaciones benéficas permitirían el crecimiento y la reproducción de los individuos
que las poseyeran en la relación con aquellos otros individuos con variaciones nocivas. En la población se generaba una lucha por la existencia en la que solo los mejores sobrevivían y extendían en la población sus características ventajosas. La mortandad de los menos adaptados era el factor que mantenía constante el tamaño de la población.
Wallace se levanto de la cama y todavía enfermo escribio estas ideas utilizando quince cuartillas de papel. Llamo su ensayo “Sobre la tendencia de las variedades a diferenciarse indefinidamente del tipo original”, lo metió en un sobre y –junto con la petición a Darwin de que si lo conmiseraba adecuado le hiciera llegar el ensayo a Charles Lyell, el mejor geólogo de su tiempo- entró con paso firme, no solo a la oficina de correo de Ternate, sino a la historia de la biología.
Charles Darwin era un hombre de costumbres fijas. El viernes 18 de junio de 1858, lo mismo que todos los días, se sentó en su sillón a revisar la correspondencia. Encontró el grueso sobre de Wallace y se dispuso a leerlo. Al terminar, su rostro estaba demudado; el impacto de una bomba no podría haberlo afectado más: línea por línea descubrió que las idea enviadas por el joven Alfred Russel Wallace coincidían exactamente con las conclusiones a las que había llegado después de veinte años de trabajo. Su cautela (no olvidemos la discusión con su mujer) le había impedido publicar sus ideas. Ahora un hombre de apenas 35 años le explicaba su propia teoría en una carta y lo enfrentaba a la terrible posibilidad de perder la prioridad de la teoría evolutiva.
Esa misma tarde Darwin mando una carta a uno de sus grandes amigos, Charles Lyell, el eminente geólogo que había demostrado, entre otras cosas, que la tierra era mucho más vieja de lo que la gente creía. En el siglo XVII el arzobispo James Ussher había calculado, por ejemplo, con métodos que el solo entendía, que la tierra había sido creada en el año 4004 a. C., precisamente el 23 de octubre. Desde luego no aclaró el espinoso asunto acerca de las actividades de Dios en marzo o septiembre del 4004 a. C.
“Sus palabras han resultado ciertas con el agravante de que se me han adelantado”, escribió Darwin a Lyell. Con esta frase, en la que refleja un toque de angustia. Darwin inició una serie de cartas que parecían revelar un enorme conflicto esquizofrénico; por un lado, veía con desesperación como veinte años de trabajo y tres de preparar un manuscrito

19
donde exponía la misma idea se iban por la borda: “No hay nada en el esquema de Wallace que no este mucho más completo en el mío que pase en limpio en 1844”, escribió el 25 de junio. Por otro lado, Darwin deseaba comportarse como el caballero que era y no despojar a Wallace de la prioridad histórica: “No seria honrado publicar ahora”, escribió el 26 de junio, y tres días más tarde agrego: “Es una bajeza de mi parte preocuparme por la prioridad”.
Las cartas en las que Darwin expresaba sus temores fueron enviadas a Lyell y Joseph Hooker, un botánico amigo. Charles acudía a sus compañeros en busca de consejo y de una salida honorable. De hecho –y esta es mi interpretación- indujo a que los personajes tomaran la extraordinaria decisión de leer el ensayo de Wallace y algunas ideas de Darwin, escritas en 1844 y 1857 (la famosa carta a Asa Gray) ante la sociedad Linneana, una agrupación de científicos que se congregaba mensualmente para analizar las novedades que se producían en el mundo de la ciencia. Esta lectura se realizó el 1º de julio de 1858, apenas 13 días después de que Darwin recibiera la carta. La decisión era extraordinaria porque a Alfred Russel Wallace no se le consulto jamas.
La respuesta de la Sociedad Linneana fue gélida; nadie hizo cuestionamientos o entro en polémicas. Sin embargo, Darwin inició a finales de julio la redacción de un manuscrito que se publicaría (y se agotaría) el 24 de noviembre de 1859 bajo el nombre de El origen de las Especies, texto que sentaría las bases de la biología moderna... Pero esa es otra Historia.
Las interpretaciones históricas ante el comportamiento de Darwin son extremas. Hay quien piensa, por ejemplo, que oculto deliberadamente la fecha en la que llegó a sus manos la carta de Wallace a fin de ganar tiempo y poder preparar su propio ensayo. Con la minuciosidad de un detective, J. L. Brooks investigo que la carta de Wallace llegó a Londres el 14 de mayo de 1858 y concluyo que debería haber sido recibida en Down, el hogar de Darwin a unas millas de Londres, solo unos días después, y no el 18 de junio.
Otros sostienen, en cambio que Charles se comporto como un perfecto caballero victoriano y se deslindo del asunto dejándolo en manos de Lyell y Hooker, quienes tomaron la decisión más correcta.
Mi personal impresión es que ninguna de las dos alternativas mencionadas describe con verdadera claridad los hechos. Darwin siempre
dio muestras de rectitud y es muy difícil imaginarlo cometiendo un acto deliberadamente deshonesto. Pero es claro que no se comporto en forma adecuada, ya que en ningún momento penso consultar a Wallace (que era lo correcto) para tomar la decisión de incluir su propio trabajo en la lectura ante la Sociedad Linneana. Por otro lado, es cierto que, en estricto derecho, la prioridad le correspondía, y el mismo Wallace lo reconoció en una carta fechada en 1887: “En aquel tiempo yo no tenia ni la más remota idea de que él (Darwin) había llegado ya a una teoría definida, y aun menos de que esta era la que se me había ocurrido de repente Ternate en 1858... No es que hubiera pensado en morirme, pero si pensaba en desarrollar la teoría de todo lo posible cuando había adelantado tanto. Puedo decir ahora, como dije hace muchos años, que me alegro de que fuera así; porque yo no siento el amor por el trabajo, por la experimentación y el detalle que eran tan preeminentes en Darwin y sin los cuales nada de lo que yo hubiera escrito habría convencido al mundo”.
Resulta interesante comparar las interpretaciones históricas que analizan los caminos por los que Darwin y Wallace arribaron a una teoría común. Se sabe que Darwin realizo un viaje de juventud que habría de ser definitivo en la construcción de su teoría, que acumulo, a lo largo de muchos años y con paciencia infinita, evidencias que apoyaron sus ideas, y se asume que a partir de este gran cuerpo de datos sistematizo su teoría siguiendo una lógica inductiva. Se cree que Wallace, en cambio, necesito un par de semanas para llegar a las mismas conclusiones y todo fue producto de un chispazo inspirado en la lectura de Malthus (como chispazos fueron la manzana de Newton o la mano de Arquímedes subiendo en la tina mientras se daba un baño).
Lo anterior es falso y verdadero a la vez. Hoy sabemos que Darwin también recibió un impulso eléctrico al leer a Malthus y que Wallace pensaba en la idea del cambio de las especies desde 1845.
En esto nos sugiere que la ciencia no se construye a partir de ideas geniales que se le ocurren a algún hombre calvo con cara de lunático sin ningún antecedente que las justifique. La ciencia tampoco se basa en la acumulación lineal de evidencia. Es en la posibilidad de interpretar esta evidencia. Es en la posibilidad de interpretar esta evidencia dentro de un contexto de ideas como adquiere sentido el pensamiento científico.

20
UNA FAMILIA MUY POCO MALTHUSIANA
Frecuentemente tenemos a recordar a algún personaje por sus acciones más conspicuas. Evocamos, por ejemplo, al general Zaragoza, con sus anteojos redondos, masticando la pólvora que ahuyento a los franceses en la batalla de puebla el 5 de mayo de 1862. Sin embargo, prácticamente nadie sabe a que se dedico su vida don Ignacio antes de defendernos gloriosamente, lo cual fue su suerte después de esta significativa fecha (de hecho, murió de tifo ese mismo año).
Para Wallace las cosas no fueron muy diferentes; sin duda el hecho más conspicuo en su vida es el envío de la carta a Darwin en marzo de 1858. Es por ello que aparece fugazmente en los libros de texto escolares. A diferencia de lo que mucha gente piensa, ese episodio no concluyó su aportación su aportación a los estudios biológicos. Después de todo, un hombre que viajo más de 30 mil kilómetros a lo largo de su vida, que escribió un total de 10 mil paginas impresas, que invadió campos tan diversos como la geografía, la antropología, la critica social y el espiritismo, no era un ser ordinario. No puede ser ordinario alguien al que Gilbert K. Chesterton, el genial novelista ingles, describió como “uno de los grandes revolucionarios del pensamiento científico y social”.
Hagamos pues un repaso de la vida de este notable naturalista.
Alfred Russel Wallace nació –en el seno de una familia que contravenía todos los consejos del reverendo Robert Thomas Malthus- el 8 de enero de 1823, en la ciudad de Usk, en Gales, Gran Bretaña. Los Wallace no tenían mucho dinero y si muchas necesidades, ya que la familia estaba formada por nueve hermanos, de los cuales Alfred era el octavo.
En 1828el padre de Alfred decidió cambiar de aires y dirigirse con su prole a Hertford, y en 1837 la familia se mudó a Londres. En ese momento Alfred se enfrento a la necesidad de mantenerse a sí mismo y abandono la escuela (de la que por cierto no tenia muy buena opinión).
Ese mismo año regreso a Gales para colaborar con su hermano William y se dedico a aprender los secretos de la topografía. William Wallace, dadas sus opiniones filosóficas y profundas ideas liberales, ejerció una saludable influencia en el joven Alfred, quien cuenta que a partir de ese momento perdió toda capacidad de modificar su pensamiento
por perjuicios religiosos o por la influencia clerical. Este aprendizaje seria definitivo para aceptar más tarde que las especies no eran inmutables, como creía la iglesia.
No estaba mal: solo tenia 14 años.
Para cumplir su trabajo Alfred realizaba por el sur de Inglaterra recorridos que marcarían su destino como un hombre interesado en conocer los secretos de la naturaleza. Se aficiono a la botánica y empezó a realizar colectas de diversos ejemplares.
En el año de 1843 no fue el mejor en la vida de Alfred; su padre falleció y su hermano William se vio en la necesidad de pedirle que dejara su empleo, dada la falta de ofertas de trabajo. Para hacer frente a sus necesidades económicas tomo el camino de la docencia y se dirigió a Leicester, donde, 1844, se convirtió
En maestro de niños acomodados. Alfred les daba clases de ingles y de dibujo. Pero eso no era todo; él mismo relata que los llevo a una conferencia sobre mesmerismo, una doctrina espiritualista creada por Mesmer, un médico alemán, pionero del hipnotismo, a finales del siglo XVIII. Durante las pláticas Wallace “mesmerizo” a sus alumnos y los hizo entrar en trance hipnótico. Sin duda no era un maestro ordinario. (Alfred era aficionado a fenómenos sobre la existencia espiritual y por esta razón recibió grandes criticas. Pero esto lo veremos más adelante.)
Wallace era un lector apasionado y pasaba muchas horas en la biblioteca de Leicester. En esta época leyó el libro de Malthus que lo conmovería años más tarde. En la biblioteca de Leicester, Alfred no solo encontró información sino también un adolescente que se convertiría en su amigo y seria determinante en su futuro: Henry Walter Bates.
Bates, a pesar de su corta edad, era un espléndido entomólogo -especialista en insectos- y rápidamente compartió conocimientos con su nuevo amigo, introduciéndolo en el estudio de los escarabajos. Sin embargo otra muerte la de su hermano William, determinó que Wallace abandonara Leicester para trabajar con otro hermano, John, en Gales. En esta época escribió su primer ensayo científico, en el que relataba la captura de Trichius fasciatus, un insecto galés... Estaba definitivamente atrapado en su vocación de naturalista.
Para completar su preparación Alfred dedico a leer obras científicas: reviso los trabajos de Lyell sobre geología, el diario de viajes de

21
Charles Darwin y una obra que lo impresionaría profundamente: Los vestigios de la historia natural de la creación, de Robert Chambers, que, como hemos visto, era un libro lleno de impresiones, pero deposito en Wallace la semilla del pensamiento evolutivo.

22
EL NATURALISTA CURIOSO Después de dos años de trabajo Alfred y su amigo Henry decidieron emprender una expedición al Amazonas para recolectar especímenes y venderlos en Europa. En marzo de 1848 se embarcaron hacia Brasil y llegaron al puerto de Para (hoy Belem) en mayo de ese mismo año. El objetivo que se había propuesto Wallace –además de procurarse un modo honesto de vivir- era muy claro: reunir evidencias para resolver el problema de origen de las especies.
Alfred permaneció los siguientes cuatro años realizando colectas en la selva de Brasil. La diversidad biológica del amazonas, infinitamente mayor que la que él conocía en Europa, lo deslumbró. A lo largo de este tiempo emprendió largas expediciones y logro reunir una magnifica colección de insectos. En 1852 padeció varias enfermedades y decidió regresar a Inglaterra (Bates se quedaría en Sudamérica hasta 1859). Al llegar a Pará se encontró con que, por un mal entendido aduanal (la eficiencia de las aduanas tampoco ha cambiado con los años), la mayoría de sus colecciones se encontraban detenidas en el puerto. Supongo que paso un mal rato pero finalmente se embarco con sus posesiones en julio de 1852, sin imaginar que lo peor aún no llegaba. En efecto, si algo puede ser más molesto que un agente aduanal, es que el barco en que viajaba a Europa se incendie y se hunda a la mitad del océano. Exactamente eso sucedió el 6 de agosto de 1852 con el barco de Wallace, quien perdió sus colecciones y cerca estuvo de perder la vida, ya que paso diez días en un bote salvavidas sin agua y alimentos, Cuando subió al velero que lo rescató, Alfred llevaba consigo un sextante y algunas notas de viaje. El resto de sus tesoros se perdió en medio del Atlántico. Por cierto, el barco que lo rescato también estuvo a punto de hundirse, ya que fue sacudido en su viaje a Inglaterra por varias tormentas. Sin embargo, logro llegar a su destino y deposito a Wallace en suelo inglés sano y salvo... pero algo deprimido.
Los siguientes dos años los invirtió Wallace en descansar, visitar museos y escribir; también asistió a reuniones científicas y conoció a Darwin. Preparo artículos sobre su viaje y escribió sobre temas tan diversos como los monos, las mariposas y la costumbre de los indios de alimentarse con insectos. Por primera vez señalo aspectos que empezaban a llamarle la atención acerca de la distribución de las especies. Pero sus obras más importantes de la época son un par de libros: Las palmas del Amazonas y Narraciones de
viajes en el Amazonas y el río Negro. Con la terquedad propia de un hombre que ha nacido naturalista, Wallace decidió emprender otra expedición a los trópicos. Eligió el Archipiélago malayo, zona descrita tan fecundamente por el escritor Emilio Salgari en sus novelas de aventuras, en las que unos piratas –los tigres de Mompracem- se dedican a hacerle la vida imposible a los hijos del imperio británico.
Gracias a la ayuda económica de la Real Sociedad Geográfica inglesa, Wallace salió hacia Singapur. Corría el mes de marzo de 1854.
El archipiélago malayo esta constituido por un gran conjunto de islas. Las más grandes son Borneo y Sumatra, aunque existe una inmensa variedad de islas más pequeñas entre las que se cuentan las Celebres y las Molucas, así como la zona norte de Nueva Guinea. En las islas predomina el relieve montañoso y hay númeroso volcanes. En 1883, poco más de veinte años después de que Wallace dejara el archipiélago (para su fortuna), uno de estos volcanes, el Krakatoa, ubicado en la isla de Rakata, entre Java y Sumatra, entro en erupción. La explosión fue tan violenta que redujo casi a la mitad el área de la isla y las cenizas arrojadas dieron tres vueltas a la tierra, produciendo atardeceres notables a lo largo de ese año en todo el mundo.
La temperatura promedio anual en el archipiélago es de 26 grados centígrados y las lluvias son muy frecuentes. El archipiélago malayo es atravesado por el ecuador, es decir la latitud geográfica 0º, y esta es una buena noticia para un naturalista, ya que algo que Humboldt, Darwin y el propio Wallace sabían es que el número de especies por unidad de superficie se incrementa hacia las áreas de baja latitud y disminuye en las altas latitudes. Y Alfred llego a Malasia precisamente en busca de especies. Claro que si el señor Wallace pretendiera cumplir con su objetivo el día de hoy seguramente se encontraría con el razonable veto de muchas sociedades ambientalistas. Pero eran otros tiempos.
Se puede decir entonces que Alfred Russel Wallace, en el archipiélago malayo, en 1854, era el hombre correcto en el lugar preciso y el momento adecuado.
Al llegar a Singapur Wallace encontró una ciudad que representaba un mosaico de razas, religiones y costumbres. El gobierno y la guarnición de la ciudad de los ingleses pero la mayoría de los habitantes eran chinos; algunos de ellos comerciantes, otros artesanos y casi todos agricultores. También

23
encontró hindúes, árabes y muchos portugueses, ya que el archipiélago había sido colonizado por Portugal.
El puerto seguramente era un espectáculo impresionante; Alfred observo navíos de guerra, barcos mercantes, buques malayos y juncos chinos. La ciudad estaba construida con grandes edificios públicos, iglesias, mezquitas y templos hindúes. Había también casas de juegos y grandes bazares. El joven Wallace estaba encantado; descubrió que los bazares chinos, atendidos por los hombres de coleta, gordos y rozagantes, ofrecian mercancías a precios más baratos que en Inglaterra. Si Alfred hacia una compra, cada vez que volvía a pasar por la tienda era saludado por el comerciante que lo invitaba a tomar un té (actitud notablemente diferente a la de nuestros terribles vendedores modernos).El joven naturalista descubrió costumbres extrañas (porque extraño es que los sastres se sienten debajo de una mesa para trabajar), fue el zapatero y el barbero y encontró excelente el servicio. En fin, Wallace conoció una ciudad maravillosa que se presentaba como el tapete de bienvenida para una estancia de ocho años y más de 20 mil kilómetros de recorrido.
Como hemos visto al inicio de este libro, Alfred trabajo de firme durante su estancia en el archipiélago malayo, recorrió prácticamente todas las islas y describió nuevas especies. Fue uno de los primeros europeos en obtener un ave de paraíso y cazo orangutanes en Bordeo (por cierto, con técnicas ligeramente crueles). En sus expediciones observo la impresionante diversidad biológica; él mismo cuenta que en un día de trabajo obtuvo ¡76 especies diferentes!, Y que en el Borneo, en un área de kilometro y medio, colecto casi dos mil especies.
Cuando la lluvia o las fiebres no lo dejaban salir a trabajar, Wallace escribía. Se han registrado 53 documento enviados a Europa con información científica, correspondientes al periodo de su estancia en Malasia. Destacan las cartas en las que aportaba datos de sus colectas, diarios de viaje, la descripción de nuevas especies e informes sobre los hábitos de algunos animales, como los orangutanes.
Dos son los documentos esenciales que escribió Alfred en esa época. El primero, redactado en febrero de 1855, se llamo ”sobre la ley que regula la introducción de nuevas especies”. En una carta a su amigo Bates comenta: “mi articulo es, por supuesto, solo el anuncio de la teoría, no su desarrollo.” El problema que Wallace pretendía resolver era
el de la a parición de nuevas especies, y fueron sus observaciones sobre la distribución de los organismos en el amazonas y el archipiélago malayo las que le dieron la respuesta: “Cada especie se ha originado en tiempos y espacios coincidentes con otra preexistente cercanamente relacionada.” En pocas palabras, Wallace proponía el bosquejo de una teoría evolutiva.
Las ideas de Alfred se publicaron en una revista inglesa en septiembre de 1855, y no despertaron una gran reaccion. Lyell le señalo a Darwin el trabajo de Wallace y lo previno acerca de una posible perdida de prioridad en su teoría. Cuando Wallace le escribió a Darwin en 1857, este le contesto que su articulo le parecía, en términos generales, correcto y con conclusiones similares a las suyas propias. Sin embargo, ya vimos que no quiso ofrecer detalles acerca de su teoría.
El segundo gran trabajo que produjo Wallace en el archipiélago malayo fue su ensayo “sobre la tendencia de las variedades a diferenciarse indefinidamente del tipo original”. En la introducción de este libro analizamos ya que el efecto que causo con su articulo, pero podemos repetirlo: fue la bomba que desencadeno la publicación de la teoría evolutiva.
Después de ocho años de trabajo Wallace decidió que ya iba siendo tiempo de regresar. Al desembarcar en Inglaterra, en febrero de 1852, su figura era ya célebre en los círculos científicos. Se lo reconocía como el autor de una teoría evolutiva obtenida por medios independientes a la de Charles Darwin –que en esos momentos libraba su lucha de aceptación-, y además se le daba un enorme reconocimiento como coleccionista y escritor científico. Pero Wallace no era un hombre rico y debía trabajar para mantenerse. Para hacerlo de manera decorosa, se dedico a escribir y dar conferencias, además de estudiar sus colecciones.

24
DE VUELTA EN INGLATERRA En 1866 Alfred se caso con Annie, la hija de un amigo suyo, el botánico William Mitten, y decidió que la vida en Londres era poco atractiva, por lo que se mudo a una zona rural. Con Annie tuvo tres hijos, uno de los cuales murió al los 4 años de edad.
En 1869 publico un libro considerado como clásico en lo que a narraciones de viajes se refiere, El archipiélago malayo, en el que presenta todas sus impresiones respecto a su experiencia en Asia.
Un año más tarde reunió una serie de diez ensayos, casi todos ya publicados, en el libro Contribuciones a la teoría de la selección natural. En ese trabajo Wallace se encargo de aportar evidencia para sostener la teoría evolutiva de la que es considerado, digamos, el segundo padre. En la introducción de su texto Wallace explicaba que mientras algunos le daban más crédito del que merecía, otros (llamémoslos los villanos) planteaban que, a pesar de haber propuesto la teoría de la selección natural, no era capaz de darle un uso a esa teoría. Para demostrar que se equivocaban Wallace presentaba a la consideración del público lector sus ensayos.
El libro es un gran collage en el que se describen observaciones sobre la capacidad de los organismos para confundirse con el ambiente, se comparan los patrones de anidacion de las aves con la habilidad humana para la construcción, se analiza el instinto de los animales y los seres humano y, finalmente, las limitaciones que para Wallace tenia la teoría de la selección natural aplicada a los seres humanos. Este fue un tema en el que polemizo con el mismo Darwin ya que Alfred no creía que estructuras tan complejas como la mete humana, la voz y el uso de los miembros, pudieran haberse originado por medio del proceso evolutivo. Los ejemplos que Wallace utilizaba eran fascinantes; hablaba por ejemplo de la capacidad para elaborar concepciones abstractas como la de la eternidad o de la posibilidad de sentir placer ante una composición artística, y se preguntaba como puede favorecer la selección natural el desarrollo de habilidades mentales que no tenían ningún uso en las etapas primitivas del hombre.
Desde luego se equivocaba, pero debemos entender que en esos tiempos resultaba muy difícil sustraerse a la idea de que el ser humano (y sobre todo los ingleses) eran criaturas aparte dentro del mundo de los seres vivos.
EL ESPIRITISTA CIENTIFICO
En 1870 Alfred fue nombrado presidente de la sociedad Entomológica de Londres, cargo que habría de conservar hasta 1872. Dos años más tarde sorprendió a todos con una vuelta de tuerca en su vida y publicó Milagros y Espiritismo Moderno, libro en el que reunió una serie de ensayos en los que planteaba sus creencias acerca de la vida de los espíritus.
Una decisión tan extraordinaria requería una explicación, y Alfred se encargo de darla en la introducción de su libro. Aclaro en primer lugar que sus creencias no se basaban en perjuicios religiosos sino en hechos tangibles y reales. Preocupado por una critica que Anton Dohrn, un científico alemán, le planteo en términos de la incompatibilidad entre la selección natural y el espiritismo, Wallace puntualizo que sus divergencias con la opinión de Darwin no tenían ninguna relación con su aceptación de la clarividencia, los milagros y las apariciones de fantasmas.
Si algunos de ustedes, lectores, después de ver lo anterior, dejan este libro en el rezago y piensan con cierta condescendencia: “Pobre Wallace, se volvió completamente loco”, me darán razones suficientes para ofrecer una explicación.
Cuando en 1844 Wallace se enfrento al mesmerismo, quedo convencido de que existen fuerzas invisibles para nuestros ojos escépticos. más aun, en su declaración de principios planteo que, si un hombre honesto Relata su observación de una serie de hechos, por inverosímiles que parezcan, no se lo deberían descalificar como un impostor o peor todavía, un imbécil, con el solo uso de prejuicios, ya que los hechos científicos tienen su base en la evidencia.
Que seria del avance científico si se aceptaran estos rechazos sistemáticos, se preguntaba Alfred.
No es que Wallace creyera en lo sobrenatural; simplemente pensaba que ciertos acontecimientos extraordinarios, como la aparición de fantasmas, no eran más que hechos físicos que debían tratar de comprenderse por medio s científicos. Es decir, era un espiritista científico, y a la tumba se fue como tal. El hecho es que muchos de sus contemporáneos se decepcionaron del curso que tomaban las ideas de Wallace.
EL BIOGEÓGRAFO
Los animales y las plantas no se distribuyen en el planeta de manera azarosa. Un esquimal

25
jamas podrá capturar un camello en alguna de sus trampas y (para su fortuna) un beduino no perderá la vida en las fauces de un terrible oso polar. Es claro que los organismos difieren de acuerdo con el sitio que habitan. Las islas, por ejemplo, son lugares en los que habitualmente existen organismos característicos que es difícil encontrar en otros lugares. Cada continente posee especies endémicas, es decir que solo habitan en esa zona. Las reglas para comprender estos patrones e distribución son esencialmente simples: Mientras más cerca del ecuador se encuentre una zona, mayor diversidad de especies existirá. Sin embargo, esto no explica porque un país como México, ubicado en una latitud mayor que Brasil, cuenta con mayor riqueza de mamíferos y reptiles. La respuesta se halla en el hecho de que nuestro país es límite de dos zonas biogeográficas y esto determina diferentes condiciones que permiten una gran abundancia de organismos. Una región biogeográfica se define por sus diferencias en cuanto a los seres que habitan una zona del planeta, y la biogeografía es la disciplina que estudia estos patrones de distribución.
Alfred Russel Wallace fue pionero en estos estudios y se convirtió en uno de los primero investigadores que propuso la división del mundo en zonas biogeográficas. En 1876 publicó su libro La distribución geográfica de los animales, en 1878 escribió Naturaleza tropical y otros ensayos y en 1880 apareció la Vida Isleña. En estos trabajos Wallace realizo un pormenorizado recuento de muy diversos temas. Analizo aspectos climáticos y como éstos determinan a las plantas y a los animales. Discutió aspectos del color de plantas y animales y su relación con la selección natural. Asimismo analizo la evidencia existente acerca del origen del hombre. En sus libros Wallace nos ofrece explicaciones acerca de los mecanismos de dispersión de los seres vivos y estudia las condiciones particulares de una docena de islas, entre las que se cuentan las Galápagos, visitadas por Darwin en 1835, y Santa Helena, la isla que murió desterrado Napoleon Bonaparte en 1821.
En estos ensayos Wallace nos ofreció una muestra espléndida de su minuciosidad para analizar detalles acerca de aspectos climáticos y de las características de la flora y fauna de cada uno de los lugares descritos, resulta claro que si solo hubiera realizado estos estudios a lo largo de su vida, tendría un lugar importantisimo entre los que configuraron la
biología como la disciplina independiente de nuestros días.
Muchos de los estudios evolutivos de Wallace tuvieron, por supuesto, una base en la geografía, y es por ello que los historiadores científicos tradicionalmente lo retratan como un evolucionista con evidencia geográfica. No obstante, podríamos decir con mayor certeza que era un (gran) geógrafo metido a temas evolutivos.
Wallace realizo viajes extraordinarios y el informe de sus experiencias llena de cientos de páginas; se convirtió en un pionero en estudio de vida tropical; sus observaciones de la conducta humana durante sus viajes le permitieron realizar importantes observaciones etnográficas; realizo estudios sobre aspectos de geografía física y económica.
¿Se podría pedir más?
En 1870 fue premiado con la medalla de oro de la Sociedad Geográfica Francesa y en 18892 recibió una importante medalla por parte de la Real Sociedad Geográfica de Inglaterra. Sin embargo, en nuestros días poco parecen tener presente su enorme contribución al avance de la geografía, lo que no deja de ser una lastima. Por cierto, su ardor por el análisis de la vida natural nunca lo abandonó: en 1877 emprendió una excursión botánica a Bélgica y en 1895 viajo con el mismo objetivo a Suiza. Tenía 72 años.
EL LUCHADOR SOCIAL
Aunque nunca abandonó la biología, al final de su vida Alfred Wallace se interesó por las causas sociales y en 1881, cuando se formo la Sociedad para la Nacionalización de la Tierra, fue su primer presidente. Estaba convencido de que el cambio social era una manifestación de la evolución. Creía que los procesos sociales no eran más que uno de los eslabones refinados de un proceso que había iniciado biológicamente, y que se deberían orientar hacia la cooperación entre los seres humanos. Toda su obra sobre el tema se enfoco hacia el logro de esos propósitos.
Desde la perspectiva de Wallace el capitalismo era uno de los grandes culpables de los problemas sociales. Él proponía decididamente medidas como la nacionalización de la tierra, la desaparición de las grandes empresas, la búsqueda de mayores derechos para las mujeres, que en esos momentos (y para nuestra vergüenza aún hoy) no eran tratadas igual que los hombres. Alfred también

26
promovió el socialismo. No olvidemos que don Karl Marx había publicado ya sus teorías, y Wallace creí firmemente en una sociedad igualitaria en la que los demonios del capital no marcaran el paso.
Era un gran utopista, pero sus ideales respondían, sin duda, a los más elevados propósitos que pueden plantearse los seres humanos.
Mientras peleaba por la igualdad, Alfred Russel Wallace continuaba con sus esfuerzos científicos. Entre octubre de 1887 realizo un viaje a América para dar una serie de conferencias. Visito Canadá y Estados Unidos. Con el material que reunió publico, en 1889, su libro Darwinismo, en el que realiza la mejor síntesis de todas las ideas evolutivas de su tiempo.
El nuevo siglo encontró a un hombre de casi 80 años que escribía, daba conferencias y asistía a convenciones espiritistas. Parecía que la energía de Alfred no tendría fin.
En 1905 escribió su autobiografía y en 1908 fue condecorado con la orden del Mérito Británico.
Alrededor de 1900 el Darwinismo sufrió gran serie de ataques que eran inspirados por el resurgimiento de una filosofía más tradicional de la naturaleza. Algunos biólogos no habían sido capaces de sacudirse a Lamarck de la cabeza y proponían que la selección natural era solo un mecanismo secundario. A esta etapa de la historia de la ciencia se le conoce como “el eclipse de Darwinismo” y hubo quienes de plano extendieron un certificado de defunción a las ideas de Darwin y Wallace. El alemán Eberhardt Dennert escribió una obra llamada En el lecho de muerte del Darwinismo, cuyo titulo refleja las enormes discusiones de la época.
Todos sabemos que la teoría darwiniana se impuso a estas críticas y que, gracias a las aportaciones de la genética, la paleontología y la embriología, se ha convertido en la base conceptual de la biología. Pero Wallace ya no fue testigo de ese triunfo. En sus últimos años fue considerado como una especie de “reliquia viviente”.
Alfred Russel Wallace publico todavía en 1913 un par de libros y firmo un contrato para escribir dos ensayos monográficos, pero la muerte lo sorprendió el viernes 7 de noviembre de 1913 en Dorset. Tenía 90 años.
La historia no ha sido suficientemente justa con Wallace, un hombre que se impuso a sus propias condiciones de existencia y nos
demostró que el esfuerzo, asociado con un pensamiento sistemático y crítico, es capaz de alcanzar cualquier meta.
Quien sabe, a lo mejor Wallace esta esperando su oportunidad para manifestarse por medio de una tabla ouija y mesmerizarnos con su último adiós. Aunque francamente no lo creo. Después de todo yo también (y en gran medida gracias a él) soy un escéptico.
TEXTOS DE WALLACE
SOBRE LA TENDENCIA DE LAS VARIEDADES A ALEJARSE INDEFINIDAMENTE DEL TIPO ORIGINAL.
La inestabilidad de las variedades como supuesta prueba de la distinción permanente y original de las especies.
Uno de los más poderosos argumentos aducidos para probar la distinción permanente y original de las especies es que las variedades producidas en estado domestico son más o menos inestables, y a menudo presentan una tendencia, si las deja solas, a retornar a la forma normal de la especie progenitora; y esta estabilidad es considerada una peculiaridad característica de todas las variedades, incluso de aquellas que se producen entre los animales salvajes en su estado natural, y se cree que constituye una previsión para preservar inalteradas las especies originalmente creadas.
Ante la falta de escasez de hechos y observaciones en torno a las variedades que se producen entre los animales salvajes, este argumento ha sido de gran peso para los naturalistas y ha llevado a una creencia muy generalizada, y un tanto prejuiciada, en la estabilidad de las especies. Igualmente general, no obstante, es la creencia en las llamadas "variedades permanentes o verdaderas” razas de animales que propagan continuamente su propia naturaleza pero que difieren de manera tan ligera (aunque constante) de alguna otra raza, que alguna de ellas es considerada una variedad de la otra. Normalmente no existe otra forma de determinar cual es la variedad y cual es la especie original, excepto en aquellos contados casos en los que una de las razas ha producido una descendencia distinta de sí misma y similar a la otra. Esto, no obstante, parece incompatible con la “invariabilidad permanente de las especies”, pero esta dificultad queda obviada si se asume que estas variedades tienen limites estrictos y no pueden llegar a variar más con respecto al tipo original, aunque puedan regresar a él, lo

27
que, por analogía con los animales domésticos, es considerado altamente probable.
Se podrá observar que este argumento descansa por completo sobre el supuesto de que las variedades que aparecen en la naturaleza son, en todos los aspectos, análogas e incluso idénticas a las de los animales domésticos y están gobernadas por las mismas leyes en lo que se refiere a su permanencia o a su posterior variación. Pero el objeto del presente trabajo es mostrar que este supuesto es totalmente falso, que existe un principio general de la naturaleza que hace que muchas variedades sobrevivan a la especie progenitora y den lugar a sucesivas variaciones que se alejan cada vez más del tipo original, y que produce también en los animales domésticos la tendencia de las variedades a retornar a la forma progenitora.
LUCHA POR LA EXISTENCIA
La vida de los animales salvajes es una lucha por la supervivencia.
Deben utilizar al limite todas sus facultades y sus energías para preservar su propia existencia y proveer a la de sus jóvenes descendientes, la posibilidad de obtener alimentos durante estaciones menos favorables y la de escapar de los ataques de sus enemigos más peligrosos son las condiciones primarias que determinan la existencia de individuos y de especies enteras. Determinarán también la población de una especie, y gracias a una cuidadosa consideración de todas las circunstancias podemos llegar a comprender, y en alguna medida a explicar, lo que a primera vista parece inexplicable: la excesiva abundancia de algunas especies mientras que otras, muy próximas a ellas, son muy escasas.
LA LEY POBLACIONAL DE LAS ESPECIES.
La proporción general que debe darse entre ciertos grupos de animales es fácil de ver. Los grandes animales no pueden ser tan abundantes como los pequeños, los carnívoros han de ser menos númerosos que los herbívoros. Las águilas y los leones nunca podrán ser tan abundantes como las palomas y los antílopes. Los asnos salvajes de los desiertos tártaros no pueden igualar en número a los caballos de las feraces praderas y pampas de América. La mayor o menor fecundidad de un animal se considera a menudo como una de las causas
fundamentales de su abundancia o escasez, pero si tenemos en cuenta los hechos veremos que, en realidad, poco o nada tiene que ver con ello.
Hasta el menos prolifero de los animales crecería rápidamente en número si no se le impusieran limites, mientras que es evidente que la población animal del globo debe estar en estado estacionario, o tal vez, por influencia del hombre, decreciendo. Puede haber fluctuaciones, perro un crecimiento permanente, excepto en localidades restringidas, es casi imposible. Por ejemplo, nuestras propias observaciones deben convencernos de que las aves no siguen aumentando en número año tras año en razón geométrica, como ocurriría si no existiera algún poderoso factor limitativo para poner freno a su crecimiento natural. Muy pocas aves producen menos de dos crías al año, mientras que muchas tienen seis, ocho o diez. Cuatro es un número sin duda inferior a la media, y si consideramos que cada pareja produce crías tan solo cuatro veces a lo largo de su vida, esto estaría también por debajo de la media, suponiendo que no mueran ni por violencia ni por falta de alimentos. Con todo, y a este ritmo, el incremento seria enorme en unos pocos años a partir de solo una pareja. Un simple cálculo nos mostrará que en 15 años cada pareja de aves habría producido casi 10 millones de descendientes. Sin embargo, no tenemos motivos para suponer que él número de aves de cualquier país crezca en absoluto en 15 o en 150 años. Con tales capacidades de crecimiento, la población debe haber alcanzado su límite y debe encontrarse en estado estacionario desde pocos años después del origen de cada especie. Es evidente, por lo tanto, que cada año debe aparecer un número inmenso de aves -de hecho tantas como nacen- y ya que, como mínimo, la progenie es cada año el doble de númerosa que sus progenitores, cualquiera que sea él número medio de individuos de un país dado, deben perecer anualmente el doble; Un resultado llamativo, pero que da la impresión de ser, al menos, muy probable, y que seguramente no esta por encima sino por debajo de las cifras verdaderas. Por consiguiente, parecería que en lo que a la continuación de la especie y el mantenimiento del número medio de individuos se refiere, las grandes camadas resultan superfluas. En términos generales, todos los pichones, menos uno, se convierte en presa de los halcones y gavilanes, gatos salvajes y comadrejas, o perecen de frío y hambre al llegar el invierno. Esto queda espectacularmente demostrado en el caso de

28
las especies particulares, ya que descubrimos que la abundancia de sus números no tiene relación alguna con su fertilidad en el momento de producir descendencia. Tal vez el caso más notable de una población inmensa de aves el de la paloma migratoria de Estados Unidos, que pone tan solo uno o, como mucho, dos huevos y se dice que normalmente llega a criar un solo polluelo. ¿Por qué es esta ave tan extraordinariamente abundante, mientras que otras producen dos o tres veces este número de descendientes son mucho menos númerosas? La explicación no es difícil. La comida preferida de esta especie, y con la que más prospera, esta abundantemente distribuida a lo largo de una región muy extensa, en la que hay tales diferencias de suelo y de clima que en uno u otro lugar existe siempre alimento. Esta ave es capaz de un vuelo muy rápido a largas distancias, por lo que puede volar sin fatiga sobre la totalidad del territorio que habita y, en el momento que falta la comida en un lugar, descubre sin dificultad otro sito donde alimentarse. Este ejemplo nos muestra de modo llamativo que obtener un suministro constante de comida nutritiva es casi la única condición necesaria para garantizar un rápido crecimiento de una especie dada, ya que ni la limitación de la fecundidad ni los ataques que las aves de presa y del hombre resultan suficientes para ponerle freno. No existe otra ave en la que estas peculiaridades circunstancias estén tan llamativamente combinadas. O bien su alimento es más propenso a escasear, o no disponen de la suficiente capacidad de vuelo para buscarlo en territorio extenso, o durante alguna estación del año llega a escasear mucho, con lo que han de buscar sustitutos menos apropiados y por consiguiente, aun siendo más fértiles en su capacidad de crianza, jamas pueden aumentar en número más allá de la disponibilidad de alimentos en las estaciones menos favorables.
Hay muchas aves que cuando escasean sus alimentos, solo pueden sobrevivir por medio de la migración, volando hacia regiones de clima más suave o al menos diferente aunque, dado que estas aves migratorias rara vez son excesivamente abundantes, esta claro que los países que visitan son también deficientes en lo que a abundancia y constancia de alimento se refiere. Aquellas aves cuya organización no le permite emigrar al escasear periódicamente sus alimentos no pueden alcanzar nunca grandes poblaciones.
Probablemente esta es la razón por lo que los pájaros carpinteros son escasos en Inglaterra,
mientras que en los trópicos se encuentran entre las aves solitarias más abundantes. Así, el gorrión común es más abundante que el petirrojo debido a que su alimento es más constante y abundante: las semillas de las gramíneas se preservan durante el invierno, y nuestras granjas y campo de cultivo les suministran una fuente de alimentos casi inagotable. ¿Por qué, como regla general, las poblaciones de aves acuáticas y en especial de aves marinas son muy númerosas? No porque sean más prolíficas que otras, ya que en general es al contrario, sino porque su fuente de alimentos nunca falla: las playas y las riberas están siempre llenas de pequeños moluscos y crustáceos.
Estas mismas leyes aplicaran a los mamíferos. Los gatos salvajes son prolíficos y tienen relativamente pocos enemigos; ¿por qué entonces no alcanzan nunca la abundancia de los conejos? La única respuesta comprensible es que su fuente de alimentos es más precaria.
Resulta por tanto evidente que, mientras un territorio permanezca físicamente inalterado, el número de sus poblaciones animales materialmente no pueden aumentar. Si una especie crece, otras que requieren el mismo tipo de alimento deben disminuir de manera proporcional. El número de muertes anuales debe ser inmenso y, dado que la existencia individual de cada animal depende de sí mismo, los más débiles –los muy jóvenes, los enfermos, los ancianos-, mientras que aquellos que consiguen subsistir solo podrán ser los más perfectos en cuanto a salud y vigor, los más capaces de obtener alimentos regularmente y eludir a sus múltiples enemigos. Es como comentamos al principio, una “lucha por la existencia” en la que los seres más débiles y los que están menos perfectamente organizados deben siempre sucumbir.
LA ABUNDANCIA O LA RAREZA DE LAS ESPECIES DEPENDEN DE SU MEJOR O PEOR ADAPTACIÓN A LAS CONDICIONES DE SU EXISTENCIA.
Esta claro que lo que ocurre entre los individuos de una especie debe ocurrir también en las diversas especies aliadas de un grupo; es decir, que los que están mejor adaptados para obtener un suministro de alimento y defenderse de los ataques de sus enemigos las vicisitudes de las estaciones deben obtener y preservar necesariamente una población superior, mientras que aquellas especies que por algún defecto en sus

29
capacidades u organización sean los menos capaces de contrarrestar las alteraciones en el suministro de alimentos, etc., disminuirán en número y en casos extremos llegaran a extinguirse. Entre estos extremos la especie presenta diversos grados de capacidad para garantizar los medios que preserven su existencia, y así como puede explicarse la abundancia o la escasez de las especies. En general, nuestra ignorancia nos impedirá seguir con precisión la pista de los efectos hasta sus causas, pero si pudiéramos familiarizarnos perfectamente con la organización y hábitos de las diversas especies de animales, y si pudiésemos medir la capacidad de cada una de ellas para desempeñar las diferentes tareas necesarias para su seguridad y supervivencia en todas las circunstancias variables que la rodean, podríamos incluso al calcular la abundancia proporcional de individuos que es un resultado inevitable. Si ya hemos conseguido establecer estos dos puntos:
1] Que la población animal de un territorio es normalmente estacionaria, manteniendose así por la deficiencia periódica de alimentos y otros factores limitativos.
2] Que la abundancia o escasez comparativas de los individuos de las diversas especies obedece exclusivamente a su organización y a los hábitos que de ella resultan, la cual, al hacer más difícil que obtengan un suministro regular de alimentos y garantizarles su seguridad, en algunos casos más que otros, solo puede verse equilibrada por diferencias en las poblaciones que han de coexistir en un territorio dado, estaremos en condiciones de proceder al examen de las variedades, respecto a las cuales las anteriores observaciones tienen una aplicación directa y muy importante.
LAS VARIACIONES FAVORABLES TENDERAN A INCREMENTARSE; LAS VARIACIONES SIN SU USO O DAÑINAS A DISMINUIR.
La mayor parte o tal vez todas las variaciones de la forma típica de una especie deben tener algún efecto definido, por pequeño que sea, sobre los hábitos o la capacidad de los individuos. Hasta un cambio de color podría afectar su seguridad haciéndolos más o menos visibles; un mayor o menor desarrollo del pelo podría modificar sus hábitos. Los cambios más importantes, tales como un aumento en la fuerza o en la dimensión de las extremidades o de cualquier órgano exterior, afectarían en mayor o menor medida su forma de obtener alimentos o la extensión del territorio que
habitan. Es evidente que la mayor parte de los cambios afectarían asimismo su capacidad para prolongar su existencia favorable o desfavorablemente. Un antílope con patas más cortas o más débiles padecería necesariamente más bajas ante los ataques de los carnívoros felinos.
La paloma silvestre de alas menos fuertes se vería afectada tarde o temprano en su capacidad de obtener alimentos de manera regular y en ambos casos el resultado seria inevitablemente una disminución de la población de la especie modificada. Si, por otra parte, una especie cualquiera llegara a producir una variedad con una capacidad ligeramente mayor para la supervivencia, esa variedad adquiriría con el tiempo, inevitablemente, una superioridad numérica. Estos resultados son tan seguros como que la ancianidad, la violencia o la escasez de alimentos producen un aumento en la mortalidad. En ambos casos puede haber multitud de excepciones individuales, pero en general esta norma debe cumplirse. Por consiguiente, todas las variedades pertenecen a una de las dos clases: aquellas que en igualdad de condiciones jamas alcanzarían una población equivalente a la de la especie progenitora, y aquellas que con el tiempo pueden alcanzar y mantener una superioridad numérica.
Supongamos ahora que se produce determinada alteración en las condiciones físicas en el territorio –una sequía prolongada, la destrucción de la vegetación por alguna plaga de langostas, la irrupción de un nuevo animal en busca de “nuevos pastos”-, en resumen, cualquier cambio que dificulte aun más su existencia de la especie en cuestión y que ponga a prueba su capacidad de evitar un exterminio total. Es evidente que, todos los individuos que forman la especie, aquellos que compongan la variedad menos númerosa y más débilmente organizada serán los primeros en padecer los efectos de este cambio, y que se extinguirán en caso de una presión excesiva. De continuar en acción las mismas causas, la siguiente en padecer sus efectos seria la especie progenitora, que disminuiría gradualmente en número y podría llegar también a extinguirse con una recurrencia similar de condiciones desfavorables. Quedaría, pues, tan solo la variedad superior, y aumentaría rápidamente su número al reaparecer las condiciones favorables, ocupando el lugar de la especie y la variedad extintas.

30
LAS VARIEDADES SUPERIORES REEMPLAZARAN A LAS ESPECIES ORIGINALES.
La variedad habría pues sustituido a la especie, de la que seria una versión más perfectamente desarrollada y más altamente organizada. Estaría mejor adaptada en todos los aspectos para garantizar su seguridad y para prolongar la existencia de sus individuos y de la raza. Tal variedad no podría retornar a la forma original, ya que ésta es una forma inferior y jamas podría competir con ella por la supervivencia. Existe por lo tanto una “tendencia” a reproducir el tipo original de la especie, pero aun así la variedad debe ser siempre predominante en número y, bajo condiciones físicas adversas, seria, una vez mas, la única en sobrevivir. Pero esta nueva raza mejorada y populosa podría, a su vez, con el paso del tiempo, dar lugar a nuevas variedades con diversas modificaciones divergentes en su forma, cualquiera de las cuales, al tender a aumentar las facultades para la preservación de la vida, debería llegar a ser predominante por la ley general.
Nos encontramos, pues, ante una progresión y divergencia continuas deducidas de las leyes generales que regulan la existencia de los animales en estado natural, y ante el hecho indiscutible de que las variedades aparecen con frecuencia. No obstante, no pretendemos afirmar que este resultado seria invariable; un cambio en las condiciones físicas de la región podría, en ocasiones, modificarlo materialmente, haciendo que la raza que antes había sido capaz de sobrevivir con más facilidad dejara de hacerlo, e incluso haciendo que la nueva raza, temporalmente superior, llegara a extinguirse, mientras que la antigua especie o especie progenitora y sus primeras variedades continuarían floreciendo. Pueden producirse también variaciones también variaciones en partes sin importancia, sin efectos perceptibles en la capacidad de supervivencia, y este tipo de variedades podrían recorrer un camino paralelo al seguido antes por la especie progenitora, bien dando lugar a posteriores variaciones o retornando al tipo original.
Lo único que planteamos es que ciertas variedades tienden a mantener su existencia durante más tiempo que la especie original, y que esta tendencia tiene que reflejarse de algún modo, ya que aunque la doctrina de las probabilidades y de los valores medios nunca resulta fiable en una escala limitada, si se aplica a grandes númerosos resultados se
aproximan a los que exige la teoría y al acercarnos a un número infinito de ejemplos se convierten en resultados precisos. Ahora bien, la escala a la que actúa la naturaleza es tan vasta, el número de individuos y los periodos de tiempo sobre lo que actúa se aproximan tanto al infinito, que cualquier causa, por pequeña que sea, y por susceptible que sea de verse ocultada y contrarrestada por circunstancias accidentales, acabara produciendo sus legítimos resultados.
EXPLICACION DE LA REVERSIÓN PARCIAL DE LAS VARIEDADES DOMESTICADAS.
Pensemos ahora en los animales domésticos y preguntémonos como las variedades entre ellos producidas se ven afectadas por los principios enunciados aquí. La diferencia esencial de condición entre los animales salvajes y los domésticos es la siguiente: entre los primeros, su bienestar y su propia existencia dependen del ejercicio y la condición saludable de todos sus sentidos y capacidades físicas, mientras que, entre los últimos, éstos son solo utilizados de modo particular y en algunos casos, no se utilizan en absoluto. Un animal salvaje tiene que buscar, a menudo trabajosamente, cada bocado de su comida; debe utilizar la vista, el oído y el olfato en su busca, en la vigilancia ante los peligros, en conseguir abrigo para la inclemencia de las estaciones y para obtener alimentos y obtener alimento y garantizar la subsistencia de sus descendientes. Debe ejercitar a diario hasta él último músculo de su cuerpo; No hay sentido o facultad alguna que no se vean fortalecidos por un ejercitamiento continuo. Por el contrario, el animal domestico dispone de un suministro continuo de alimentos, vive protegido y a menudo confinado para evitar que lo afecten las vicisitudes de las estaciones, es cuidadosamente defendido de los ataques de sus enemigos y rara vez llega a criar a sus descendientes sin ayuda del hombre. La mitad de sus sentidos y sus facultades le resultan prácticamente inútiles, y el resto de ellos no entra en acción más que de vez en cuando, incluyendo sus músculos, que utiliza solo de manera ocasional.
Ahora bien, cuando aparece una variedad de un animal así, dotada de una mayor capacidad o fuerza en cualquier órgano o sentido, estas mejoras resultan totalmente inútiles; jamas llegan a entrar en acción y pueden incluso existir sin que el animal llegue a ser consciente de ellas. En el animal salvaje, por el contrario, al ser utilizadas al limite todas

31
sus facultades y capacidades para complementar las necesidades de la existencia, toda mejora tiene un efecto inmediato, se ve fortalecida por el ejercitamiento e incluso debe modificar ligeramente, los hábitos y la economía total de la raza. Crea, por así decirlo, un nuevo animal, un animal de poderes superiores, que necesariamente aumentara en número y sobrevivirá a aquellos que sean inferiores a él.
También en los animales domésticos todas las variaciones tienen la misma oportunidad de verse preservadas, y aquellas que harían sin duda de un animal fuera incapaz de competir por su supervivencia no suponen desventaja alguna en estado doméstico. Nuestros cerdos de rápida engorda, las ovejas de patas cortas, las palomas buchonas y los perros falderos jamás habrían aparecido en la naturaleza, ya que en el primer paso hacia estas formas inferiores habría llevado a una rápida extinción de la raza. Y aun menos podrían existir hoy en competencia con sus parientes silvestres. La gran velocidad por escasa resistencia del caballo de carreras, la torpe potencia de los caballos de arrastre, resultarían ambas inútiles en estado natural. Si fueran liberados en las pampas, es probable que tales animales se extinguieran rápidamente o en circunstancias favorables, tal vez perdieran esas cualidades extremas que jamas serian utilizadas, y en el transcurso de unas pocas generaciones revertirían a un tipo común que habría de ser aquel en el que las diversas capacidades y facultades existieran en una proporción capaz de adaptar óptimamente al animal para obtener alimento y seguridad: aquel en el que por medio del ejercitamiento total de cada parte de su organización el animal se asegura la supervivencia. Las variedades domesticas, al volverse silvestres, necesitan retornar a un tipo similar al tipo silvestre original o se extinguen totalmente.
Vemos pues que no pueden hacerse inferencias acerca del estado natural a partir de la observación de lo que ocurre a las variedades en el caso de los animales domésticos. Ambas situaciones son tan opuestas que lo que se aplica a una casi con seguridad no puede aplicarse a la otra. Los animales domésticos son anormales, irregulares, artificiales; presentan variedades que jamas aparecen ni pueden aparecer en estado natural. Su propia existencia depende de los cuidados humanos, hasta tal punto han llegado a alejarse algunos de ellos de esa equilibrada proporción de facultades, de ese autentico equilibrio en la organización por
medio del cual un animal puede preservar su existencia y da continuidad a su raza.
LA HIPÓTESIS DE LAMARCK ES MUY DIFERENTE DE LA QUE AQUÍ SE ADELANTA.
La hipótesis de Lamarck – que los cambios progresivos en las especies se han producido debido a los intentos de los animales por incrementar el desarrollo de sus propios órganos, modificando su estructura y sus hábitos- ha sido refutada repetida y fácilmente por todos los autores que abordan la cuestión de las variedades y las especies, y al parecer se ha dado por sentado que una vez hecho esto, la cuestión había quedado zanjeada. Pero la perspectiva aquí expuesta hace que esa hipótesis resulte innecesaria, al demostrar que pueden obtenerse resultados similares por la acción de principios que actúan de modo constante en la naturaleza. Las poderosas garras retráctiles de las tribus de los halcones y los gatos no han sido producidas ni mejoradas por el deseo de esos animales, sino que entre las diferentes variedades aparecidas en las formas anteriores y menos organizadas de estos grupos siempre sobrevivieron más tiempo aquellos que tenían mayor facilidad para aferrar a su presa. Tampoco la jirafa adquirió su cuello largo deseando alcanzar el follaje de los arbustos más altos y estirando el cuello para llegar a ellos, sino porque aquellas variedades que aparecieron con un cuello más largo que el normal obtuvieron inmediatamente una nueva fuente de alimentos en el mismo territorio que ocupaban sus compañeras de cuello corto, por lo que en la primera ocasión en que hubo escasez de comida consiguieron sobrevivir más aquellas. Incluso los peculiares colores de muchos animales, especialmente insectos, que tanto se asemejan al suelo o a las hojas o troncos sobre los que habitualmente residen, quedan explicados por el mismo principio, ya que aunque en el transcurso de las eras pueden haber aparecido variedades de muy diversos colores, las razas que por su color están mejor adaptadas para ocultarse de sus enemigos inevitablemente sobrevivirán más tiempo. Tenemos aquí también una causa activa para explicar el equilibrio tan a menudo observado en la naturaleza, al verse compensada toda deficiencia de una serie de órganos por un mayor desarrollo de algunos otros – patas débiles acompañadas de alas poderosas o la velocidad como compensación de la deficiencia de armas defensivas-; así ha quedado demostrado que las variedades en las que aparece una deficiencia no equilibrada

32
no logran sobrevivir mucho tiempo. La actuación de este principio es totalmente similar a la de regulador centrifugo de la maquina de vapor, que pone freno y corrige las irregularidades prácticamente antes de que sean perceptibles. De modo similar, ninguna deficiencia no compensada puede alcanzar jamas una magnitud conspicua en el ritmo animal, ya que se haría notar desde el primer momento, al hacer difícil la supervivencia y asegurar prácticamente la extinción.
Un origen como el aquí defendido se corresponde también con el carácter peculiar de las modificaciones en forma y estructura que aparece en los seres organizados; la multitud de líneas de divergencia a partir de un tipo central, la creciente eficacia y potencia de un órgano particular a través de una sucesión de especies aliadas y la notable persistencia de partes sin aparente importancia tales como el color, la textura del plumaje y el pelo, la forma de los cuernos o las crestas, en toda una serie de especies que difieren considerablemente en caracteres más esenciales. También nos suministra una razón de esa “estructura más especializada” que, según el profesor Owen, es una característica de las formas recientes en comparación con las formas extintas, y que, evidentemente, serian el resultado de la modificación progresiva de cualquier órgano aplicado a un fin especifico en la economía del animal.
CONCLUSION.
En nuestra opinión, hemos mostrado que existe una tendencia en la naturaleza a la progresión continuada de ciertas clases de variedades hacia formas cada vez más alejadas del tipo original –una progresión a la que no parece necesario asignar limites definidos-, y que el mismo principio que produce este resultado en estado natural explica también porque las variedades domesticas tienen tendencia a revertir al tipo original. Esta progresión a pasos diminutos, en diversas direcciones, pero siempre controlada y equilibrada por las condiciones necesarias, sin las cuales no puede preservarse la existencia, puede, en nuestra opinión, corresponderse con todos los fenómenos presentes en los seres organizados, con su extinción y su sucesión en eras remotas, y con todas las extraordinarias modificaciones de forma, instintos y hábitos que exhiben.

33
EL ARCHIPIÉLAGO MALAYO
BORNEO, EL ORANGUTÁN.
Llegue a Sawak el 1º de noviembre de 1854 y me fui el 25 de enero de 1856. En este lapso residí en muchos lugares diferentes y conocí bastante a las tribus nativas, los dyak, así como a los malayos de Borneo.
En marzo de 1855 decidí ir a las minas de carbón que comenzaban a explotarse cerca del río Simunjon, muy estrecho y sinuoso, sombreado por las altivas selvas que a veces se cierran casi sobre él. Toda la región que media entre el río y el mar es un pantano perfectamente plano y cubiertos de bosques, del cual surgen unas pocas colinas aisladas, al pie de una de las cuales se ubican las minas. Desde el desembarcadero hasta la colina se ha trazado un camino Dyak, que consiste exclusivamente en troncos puestos uno tras otro. Los nativos descalzos, que llevan pesadas cargas, caminan por ellos con la mayor facilidad, pero para un europeo calzado con botas es un trayecto de lo más resbaloso, y cuando su atención se ve atraída constantemente por los diferentes objetos de interés que lo rodean, unas cuantas caídas a la ciénega son casi inevitables. Durante mi primer paseo por este camino vi pocos insectos y aves, pero pude observar unas hermosas orquídeas en flor del genero coelogyne, grupo que, según descubrí después, es muy abundante y característico de esa zona.
Como el lugar me resultó muy adecuado, y me brindaba grandes facilidades para la recolección, me hice construir una casita de dos habitaciones con una galería. Me quede allí casi nueve meses, y reuní una inmensa una inmensa colección de insectos, a los que dedicaba mi atención primordial, ya que las circunstancias para ello eran especialmente favorables.
En los trópicos una gran proporción de los insectos de todos los órdenes, y en especial del gran grupo de los escarabajos, depende en mayor o menor medida de la vegetación y sobre todo de la madera, la corteza y las hojas en diversos estados de putrefacción. En los bosques vírgenes los insectos están dispersos por un territorio inmenso, en lugares en los que los árboles se han caído por viejos y enfermos o han sucumbido a la furia de las tempestades, y un pequeño claro pude contener más árboles caídos y en descomposición que 100 kilómetros cuadrados del terreno. La cantidad y la variedad de escarabajos y de muchos otros insectos que
pueden colectarse en un momento dado en una localidad tropical dependerá, primero de la cercanía de una gran extensión de selva virgen y, segundo, de la cantidad de árboles que durante los últimos meses se hayan cortado y hayan dejado secar en el piso. Durante doce años que pasé colectando en los trópicos, tanto orientales como occidentales, nunca goce de tantas ventajas en este sentido como en las minas de carbón de Simunjon. Durante varios meses entre veinte y cincuenta chinos y dyaks se dedicaron casi exclusivamente a limpiar un gran claro en la selva y a abrir un amplio espacio para el ferrocarril que lo uniría con el río Sadong, a tres kilómetros de distancia. Además, se establecieron aserraderos en diversos lugares de la jungla y se derribaron grandes árboles para hacer vigas y tablones. Un bosque magnifico se extendía cientos de kilómetros en todas direcciones, sobre llanuras y montañas, rocas y pantanos, y llegue allí justo cuando las lluvias comenzaban a menguar y el sol brillaba más tiempo cada día, temporada que siempre me ha parecido la más favorable para la colecta. Los númerosos claros, lugares soleados y senderos eran un atractivo para las avispas y las mariposas; pagaba un centavo por cada insecto que me llevaban, y obtuve tanto de los dyaks como de los chinos muchas langostas y fásmidos, al igual que atractivos escarabajos.
Cuando llegue a las minas, el 14 de marzo, había colectado, en los cuatro meses previos, 320 clases diferentes de escarabajos. En menos de una quincena duplique este número, con promedio de unas 24 especies nuevas por día. Hubo un día que recolecte 76 clases distintas, 34 de las cuales eran nuevas para mí. Para finales de abril tenia ya más de 1000 especies, que siguieron acumulándose, aunque a un ritmo más lento, de manera que en Borneo obtuve un total de 2000 tipos distintos, todos los cuales, con excepción de unos cien, fueron colectados en ese lugar, en una extensión de poco más de 4 kilómetros cuadrados. Los grupos de escarabajos más númerosos y atractivos fueron los de los longicornios y el de los Rhyncophora, ambos comedores de madera. Los primeros que se caracterizan por su forma graciosa y sus largas antenas, eran muy númerosos y sumaban casi trescientas especies, de las cuales noventa por ciento eran totalmente nuevas, y muchas resultaban notables por su gran tamaño, su extraña forma y su hermosos colorido.
Mi colección de mariposas no era muy grande, pero obtuve algunos insectos raros y muy

34
bellos; el más notable fue el Ornithoptera brookeana, que es una de las especies más elegantes que se conocen. Esta hermosa criatura tiene alas muy largas y puntiagudas, casi como las de una polilla esfinge. Es de un negro profundo, con una línea curva de manchas de un brillante verde metálico que se extiende de punta a punta de las alas; cada mancha tiene la forma exacta de una plumita triangular, y el efecto se asemeja mucho a una hilera de plumas de las alas de un quetzal mexicano, dispuestas sobre terciopelo negro. Las únicas otras marcas son un ancho collar de un vivido escarlata, y unos pocos toques de un blanco delicado en el borde exterior de las alas posteriores.
Esta especie que era desconocida hasta entonces, y a la que di nombre, era muy rara. Se la veía a veces, volando velozmente en los claros y posándose de tanto en tanto en charcos y lugares lodosos, de modo que solo logré capturar dos o tres especímenes, pero hasta ahora todos han sido machos, y no podemos conjeturar como será la hembra. Uno de los reptiles más curiosos e interesantes que encontré en Borneo fue una gran rana arborícola que me llevo uno de los trabajadores chinos. Me aseguro que la había visto descender en diagonal de un alto árbol, como si volase. Al examinarla vi que tenía dedos muy largos y unidos con una membrana hasta la punta, de modo que cuando se expandían representaban una superficie mucho más grande que la del cuerpo. Las patas delanteras también estaban bordeadas por una membrana y el cuerpo podía inflarse en forma considerable. El lomo y las patas eran de un verde resplandeciente y muy profundo, la superficie inferior y la cara interna de los dedos eran amarillas y las membranas interdigitales negras, con rayas amarillas. El cuerpo media unos diez centímetros de largo, mientras que las membranas de las patas raseras, al expandirse por entero, cubrían cada una superficie de diez centímetros de lado. Como los extremos de los dedos tienen discos de adhesión, lo que demuestra que se trata de una verdadera rana arborícola, es difícil imaginar que esa inmensa membrana de las patas solo tenga por propósito nadar, y se vuelve más creíble el relato del chino que dijo verla bajar volando del árbol. Creo que es el primer caso conocido de “rana voladora”, y es muy interesante para los darwinianos, ya que muestra que se ha aprovechado la variabilidad de los dedos, modificados ya para nadar y para adherirse al trepar, a fin de permitir que una especie emparentada se arroje por el aire como lagarto volador.
Uno de mis objetivos fundamentales al instalarme en Simunjon era ver al orangután (o gran mono hombre de Borneo) en su territorio nativo, estudiar sus hábitos y obtener buenos ejemplares de las diferentes variedades y especies de ambos sexos y tantos adultos como jóvenes. En todos estos objetivos mi éxito superó mis expectativas, y quiero narrar algunas de mis experiencias en la caza del orangután o “mias”, como lo llaman los nativos. Y como es un nombre corto y fácil de pronunciar, y lo usaré más que Simia satyrus u orangután. Justo una semana antes de llegar a las minas vi al primer mias. Había salido a colectar insectos, a no más de cuatrocientos metros de la casa, cuando oí un ruido de las hojas en un árbol próximo y al levantar las visa, observe a un gran animal de pelo rojizo que se movía lentamente, colgado por los brazos de las ramas. Fue pasando de árbol en árbol hasta que se perdió en la selva, tan pantanosa que no pude seguirlo. Esta forma de avanzar, no obstante, era muy poco usual, y es más característica de los hilobates que del orangután. Supongo que ese animal tenía alguna peculiaridad, o que la naturaleza de los árboles en ese preciso lugar determinaba que fuera la manera más fácil de moverse.
Unos quince días después oí decir que un mias estaba comiendo en un árbol en el pantano próximo a la casa; tomé mi escopeta y tuve la suerte de encontrarlo en el mismo lugar. Tan pronto como me acerque trató de ocultarse entre el follaje, pero logre dispararle y con el segundo tiro cayo casi muerto, ya que ambas balas lo habían alcanzado. Era un macho a medio crecer que escasamente medía un metro. El 26 de abril había salido de cacería con dos dyaks y encontramos otro más o menos del mismo tamaño. Cayó al primer disparo, pero no parecía seriamente herido y de inmediato trepo al árbol más próximo; volví a disparar y cayó de nuevo, con un brazo roto y una herida en el cuerpo. Los dos dyaks corrieron hacia él y lo tomaron por las manos mientras que me decían que cortara una rama para atarlo. Pero aunque tenía un brazo roto y estaba a medio crecer, era demasiado fuerte para esos jóvenes salvajes, a los cuales, pese a todos sus esfuerzos, jalaba hacia la boca, hasta que tuvieron que soltarlo para no sufrir serias mordeduras. El animal comenzó a trepar nuevamente el árbol y para evitar más problemas, le metí una bala en el corazón.

35
LAS ISLAS ARU. EL VIAJE Y MI RESIDENCIA EN EL INTERIOR.
Estuve en las islas Aru de marzo a mayo de 1857. Uno de mis primeros objetivos fue preguntar por la gente que acostumbre cazar las aves del paraíso. Vivían a cierta distancia, en la selva y mandaron a un hombre a buscarlos. Cuando llegaron hablamos por mediación del orang-kaya, el jefe de la tribu, quien actúo como interprete; Dijeron que creían poder conseguir algunas aves. Me explicaron que las cazaban con arcos y flechas, y que estas tenían en el extremo una punta cónica de madera, del tamaño de una taza, para matar a los pájaros por la violencia del impacto, sin lastimarlos ni derramar sangre. Los árboles a los que acuden las aves del paraíso son muy altos y frondosos, por lo que es necesario levantar un pequeño refugio de hojas entre las ramas, a la cual trepa el cazador antes de que salga el sol.
Se queda allí todo el día y cuando se posa un pájaro es casi seguro que lo atrapan. Esa misma noche los cazadores volvieron a sus casas y nunca volví a verlos; luego supe que en esa época del año no se podían obtener aves de buen plumaje.
Los dos o tres primeros días fueron muy húmedos y solo conseguí unos cuantos insectos y aves, pero por fin, cuando estaba empezando a desesperar un día regreso mi mozo Baderún con un ejemplar que me recompenso por los meses de demoras y expectativas. Era un ave pequeña, poco mayor que un mirlo. La mayor parte de su plumaje era de un intenso rojo cinabrio, con un resplandor como hilos de vidrio. Las plumas de la cabeza eran cortas y aterciopeladas, en tonos de naranja intenso. Del pecho para abajo era de un blanco puro, con la suavidad y el brillo de la seda; una banda de un oscuro verde metálico atravesaba del pecho y separaba el blanco del rojo de la garganta. Encima de cada ojo había una mancha redonda del mismo verde metálico; el pico era amarillo, y las patas y piernas eran de un bello azul cobalto, que contrastaba marcadamente con todas las demás partes del cuerpo. Por la disposición de los colores y la textura del plumaje ese pajarito era una joya de primera, y sin embargo esa no representaba más que la mitad de su extraña hermosura. A cada lado del pecho, ocultos normalmente debajo de las alas, brotaban unos mechonsitos de plumas grisáceas de unos cinco centímetros de largo; cada uno de ellos terminaba en una ancha franja de un verde esmeralda intenso. El ave
puede levantar a voluntad esas plumas, y extenderlas como un par de elegantes abanicos cuando eleva las alas. Pero éste no es su único ornamento. Las dos plumas medias de cola tienen la forma de delgados alambres de unos 13 centímetros de largo, que divergen en una hermosa curva doble. Alrededor de un centímetro, en el extremo de este alambre, muestra plumas solo por el lado externo; Es de un hermoso verde metálico y se curva en espiral hacia adentro, con lo que forma un par de elegantes botones resplandecientes que cuelgan debajo del cuerpo. Estos dos ornamentos, los abanicos de pecho y las plumas espiraladas de la cola, son algo totalmente únicos, y no se encuentra en ninguna otra de las ocho mil especies de aves que, según sabemos, existen en el mundo; combinados con el plumaje más exquisitamente bello, hacen de este pájaro una de las criaturas más encantadoras que nos ofrece la naturaleza. Mis raptos de admiración y deleite le resultaban divertidos a mi anfitrión de Aru, para el cual en burong reja como para nosotros un gorrión o un tordo. Se había cumplido así uno de los objetivos de mi viaje al Lejano Oriente. Había obtenido un ejemplar del ave real del paraíso (Paradisea regia), descrito por el Linné a partir de pieles mutiladas y conservadas por los nativos. Sabia que muy pocos europeos habían podido contemplar ese perfecto organismo que yo observaba, y que en Europa se le conocía muy mal. Las emociones que se despiertan en la mente de un naturalista que durante largo tiempo ha deseado ver el ser que solo conoce por descripciones, dibujos o pieles y plumas en mal estado de conservación, sobre todo cuando se trata de un ser de hermosura y rareza extraordinarias, solo pueden expresarse plenamente si se posee la facultad de la poesía. La isla remota en la que me encontraba, en un mar casi nunca visitado, lejos de las rutas marítimas; el lujurioso y salvaje bosque tropical, que se extendía por doquier en torno a mí; los rudos salvajes que me rodeaban... todo contribuía a las emociones que experimente al contemplar ese “ser de belleza”.
Después de obtener la primera ave real, me interne en la selva con mis hombres y no solo conseguimos otra, también con un plumaje perfecto, sino que pude ver algo de sus hábitos y de los de la especie más grande. Frecuenta los árboles bajos los bosques menos densos y es muy activa; vuela con fuerza y brinca o vuela continuamente de rama en rama. Se alimenta de unas frutas del tamaño de una cereza y de hueso duro, y agita las alas elevando los hermosos abanicos

36
que adornan su pecho. Los nativos lo llaman “gobi-gobi”. Un día me encontré debajo de un árbol donde se reunía cierto número de aves gigantes del paraíso, pero estaban muy arriba, en lo más espeso del follaje y volaban y saltaban sin cesar, de manera que no pude verlas bien. Por fin cacé una, pero era un ejemplar joven, todo de un intenso color chocolate, y no tenía la garganta verde metálica ni las plumas amarillas de las aves adultas. Todos los que había visto eran parecidos y los nativos me dijeron que tenían que pasar otros dos meses para que pudiese encontrar especímenes con todo su plumaje. Me quedaba la esperanza de conseguir alguno. Tienen una voz extraordinaria. Muy temprano, antes de que salga el sol, oímos un sonoro griterío que resuena en toda la selva y cambia constantemente de dirección. Es el ave gigante del paraíso que va a buscar su desayuno.
Me llevaron una cacatúa, la primera de este tipo que haya visto, un verdadero trofeo. Tiene un cuerpo bastante pequeño y débil, largas patas endebles, grandes alas y una cabeza enormemente desarrollada, adornada con una cresta magnifica y armada con un pico encorvado y agudo de fuerza y tamaño enormes. El plumaje es todo negro, pero esta cubierto por esa extraña secreción polvorosa y blanca característica de las cacatúas. Tiene las mejillas desnudas y de un intenso color rojo sangre. En lugar del grito áspero de las cacatúas blancas produce una especie de un silbido lastimero. La lengua es un órgano extraño, un delgado cilindro carnoso de color rojo oscuro, que termina en una placa córnea negra, ranurada y algo prensil. Habita en la parte baja de la selva; se la ve sola o sí acaso, en grupos de dos o tres aves. Vuela lenta y silenciosamente, una herida bastante leve puede matarla. Se alimenta de diversa frutas y semillas, pero parece muy aficionada a la nuez de Canarium commune, abundante en las islas en las que se encuentra esta ave. La forma en que se extrae las semillas muestra una correlación de estructura y hábitos que indicaría que esa nuez constituye su alimento fundamental. La cascara de la nuez es tan dura que solo un pesado martillo puede romperla; es más o menos triangular y su exterior es muy liso. La forma en la que cacatúa la abre resulta muy curiosa. La sostiene por un extremo y manteniéndola firme con la presión de la lengua, con movimiento lateral de la filosa mandíbula inferior va aserrando una ranura.
Después sostiene la nuez con la pata, arranca con el pico un trozo de hoja, que mantiene en
la profunda muesca de la mandíbula superior, vuelve a tomar la nuez, que no puede resbalar contra el tejido elástico de la hoja; coloca el borde de la mandíbula inferior en la ranura y con un poderoso esfuerzo, arranca un pedazo de cáscara. Se pasa otra vez la nuez a la pata, inserta en ella la punta larga y aguzada del pico y va extrayendo la carne, trocito a trocito, con la lengua. De esta manera, cada detalle de la forma y la estructura del extraordinario pico de esta ave parece tener su propio uso, y es fácil suponer que las cacatúas negras han logrado competir con sus similares blancas, más activas y númerosas, gracias a su capacidad de subsistir con un tipo de alimentos que ningún otra ave lora extraer de la pétrea cáscara.

37
LA EVOLUCIÓN INTRODUCCION GENERAL A ESTE NÚMERO EXTRAORDINARIO, EDICADO A LA HISTORIA DE LA VIDA SOBRE LA TIERRA A LA LUZ DE LA TEORIA “SINTETICA” DE LA EVOLUCION A TRAVÉS DE LA SELECCIÓN NATURAL.
(ERNEST MAYR)
El cambio que más influyo sobre la visión que el hombre tenia del mundo, de la naturaleza viviente y de sí mismo, se produjo con la introducción, que duro unos 100 años y se inicio en el siglo XVIII, de la propia idea de cambio, de cambios a lo largo de extensos periodos de tiempo, en una palabra, de la evolución. La visión que el hombre tiene del mundo hoy día esta dominada por la certeza de que el universo, las estrellas, la tierra y todos los seres vivos han evolucionados a través de una larga historia que no estuvo ordenada de antemano ni programada, una historia de cambio gradual y continuo, moldeada por procesos naturales más o menos direccionales, que concuerdan con las leyes de la física. La evolución cósmica y la evolución biológica tienen todo esto en común. Sin embargo, la evolución biológica presenta diferencias fundamentales con respecto a la evolución cósmica en númerosos aspectos. Así, la evolución biológica es más compleja que la cósmica, y los sistemas vivientes, que son los productos de la primera, mucho más complejos que con cualquier sistema no vivo; otras diferencias irán quedando de manifiesto a lo largo de este artículo. Este número de investigación y ciencia trata del origen, la historia y las interrelaciones de los sistemas vivientes. Tal como presentan a la luz de la teoría general de la vida, comúnmente aceptada: la teoría de la evolución por medio de la selección natural; propuesta hace más de 100 años por Charles Darwin y posteriormente modicada y explicada por la ciencia de la genética, y que sigue siendo en la actualidad el principio organizador de la biología.
Los mitos de los pueblos primitivos y de la mayoría de las religiones acerca de la creación tenían en común un concepto esencialmente estático de un mundo que, una vez creado, no había cambiado, y que además no llevaba mucho tiempo de existencia. El cálculo del obispo Ussher en el siglo XVII, según el cual el mundo había sido creado en el año 4004 a. C., es digno de mención aunque solo sea por su equivocada precisión, en una época en que la duración de la historia estaba todavía limitada por el alcance reducido de los testimonios escritos y de la propia tradición.
Fueron los naturalistas y filósofos de la lustración en el siglo XVIII, y los geólogos y biólogos del siglo XIX, quienes tuvieron que empezar a extender la dimensión temporal del mundo. En 174 el conde Buffon, naturalista francés, fue el primero en emprender el calculo de la edad de la tierra. Estimó que tenia 70.000 años por lo menos (y sugirió, en sus notas no publicadas, una edad de hasta unos 500.000 años). Immanuel Kant fue mucho más lejos en su cosmogonía (1775), en la que hablaba de millones e incluso de centenares de millones de años. Esta claro que, tanto Buffon como Kant, concebían un universo físico que había evolucionado.
“Evolución” implica cambio con continuidad, normalmente con un componente direccional. La evolución biológica se define mejor como cambio en la diversidad y adaptación de las poblaciones de organismos. La primera teoría coherente de la evolución la propuso, en 1809, el naturalista y filosofo francés Jean Baptiste de Lamarck, quien centró su atención en el proceso de cambio a lo largo del tiempo, es decir, en lo que parecía una progresión de la naturaleza desde los organismos visibles más pequeños hasta los animales y plantas más complejos y casi perfectos, y por tanto, hasta del hombre. Para explicar el curso particular de la evolución, Lamarck invoco cuatro principio: la existencia de los organismos de un impulso interno hacia la perfección; la capacidad de los organismos para adaptarse a las “circunstancias”, es decir, al ambiente; el hecho frecuente de la generación espontanea; y la herencia de los caracteres o rasgos adquiridos. La creencia de que los caracteres adquiridos son hereditarios, error por el que principalmente se recuerda a Lamarck, no la introdujo él. Se trataba de una creencia universal en su época, firmemente arraigada en la cultura popular (expresión de la cual es, por ejemplo, la historia bíblica de Jacob y la división del ganado rayado y moteado). La creencia continuó persistiendo. Darwin, por ejemplo, asumió que el uso o desuso de una estructura por parte de una generación podía reflejarse en la generación siguiente, lo mismo pensaron númerosos evolucionistas hasta finales de siglo, en el que biólogo alemán August Weismann demostró la imposibilidad, o por lo menos la improbabilidad, de la herencia de los caracteres adquiridos. Las suposiciones de Lamarck acerca de la tendencia a la perfección y sobre la frecuente generación espontanea, tampoco se vieron confirmadas, pero tenía razón al reconocer que buena parte de la evolución es, tal como se llama ahora, adaptativa. Además, Lamarck entendía que

38
solo era posible explicar la gran diversidad de organismos vivientes si se aceptaba el postulado de que la tierra tenía una gran antigüedad, y que la evolución era un proceso gradual.
El principal interés de Lamarck lo constituía la evolución en su dimensión temporal: la evolución vertical, por así decirlo. Darwin, por el contrario, estuvo inicialmente intrigado por el problema del origen de la diversidad, y más específicamente por el origen de la diversidad, y más específicamente por el origen de las especies a través de la diversificación en una dimensión geográfica, esto es, por la evolución horizontal. Como es bien sabido, este interés por la diversificación y la especiación despertó durante su viaje alrededor del mundo, de cinco años de duración, como naturalista a bordo del H.M.S. Beagle. Así, en las islas galápagos vio que cada isla tenía su propia forma de tortuga, de cercion y de pinzón. Las diversas formas, aunque estaban muy estrechamente relacionadas, eran completamente distintas. Al sopesar sus observaciones, de retorno ya en Inglaterra, Darwin llegó a la conclusión de que la población de cada isla constituía una especie incipiente y, con ello, a la idea de “transmutación” o evolución de las especies. En 1838, concibió el mecanismo que podría explicar la evolución: la selección natural. Después de muchos años de observación y experimentación, con amplio conocimientos de geología, zoología y otras materias, adquiridos tras muchas horas de lectura, Darwin anuncio, en 1858, un esbozo de su teoría sobre la evoluciona través de la selección natural, en una comunicación presentada a la Linnean Socity de Londres. Alfred Russel Wallace, un joven naturalista ingles que se hallaba realizando trabajos de campo en las Indias Orientales, había llegado a su vez, a la idea de selección natural, y había plasmado sus ideas en un manuscrito que envío por correo a Darwin: su comunicación se leyó en la misma sesión que el informe de Darwin. La teoría completa de Darwin, reforzada con innumerables observaciones personales y cuidadosamente argumentada, se publico el 24 de noviembre de 1859 en On the Origin of Species (“el origen de las especies”). El amplio esquema aclaratorio constaba de varias subteorias o postulados, de las que señalare las que, a mi juicio, constituyen las cuatro principales. Dos de ellas concuerdan con el modo de pensar de Lamarck: la primera es el postulado de que el mundo no es estático, sino que evoluciona; las especies cambian continuamente, se originaba unas y se extinguen otras. Las
biotas, tal como refleja el registro fósil, cambian con el tiempo; cuanto más antiguas son, más diferenciadas se nos presentan con respeto a los organismos vivos. Al contemplar la naturaleza viviente se observan fenómenos que carecen de sentido a no ser que se interpreten en el marco de la evolución. El segundo concepto lamarckiano que asumió Darwin era el postulado de que el proceso de la evolución es gradual y continuo; y que no consiste en saltos discontinuos o cambios súbitos.
Los otros dos postulados principales de Darwin encerraban, en sus grandes líneas, ideas nuevas. Uno era postulado de la comunidad de descendencia. Para Lamarck, cada organismo o grupo de organismos representaba una línea evolutiva independiente que había tenido principio en la generación espontanea y se había esforzado en constante tendencia a la perfección. Darwin, por el contrario, postulaba que los organismos semejantes estaban emparentados, y descendían de un antepasado común. Afirmaba que todos los mamíferos habían derivado e una especie ancestral, que todos los insectos tenían un antepasado común, y que, en definitiva, tal era lo que ocurría entre todos los demás organismos de cualquier grupo. Lo que de hecho implicaba que los organismos vivientes podían remontarse hasta un origen único de la vida.
Hubo muchas personas que consideraron un insulto imperdonable a la raza humana la inclusión, por parte de Darwin, del hombre de la comunidad de descendencia de los mamíferos; Ello provocó un aluvión de propuestas. Pero la idea de la comunidad de descendencia tenia un poder explicativo tan enorme que, casi inmediatamente, fue adoptada por la mayoría de los biólogos. Daba cuenta tanto de la jerarquización linneana de las características taxonómicas como del hecho, revelado por la anatomía comparada, de que podían asignarse todos los organismos a un número limitado de tipos morfológicos.
La cuarta subteória de Darwin, la de la selección natural, constituía la clave para interpretar su amplio esquema. El cambio evolutivo, decía Darwin, no es el resultado de algun misterioso impulso lamarckiano, ni una simple cuestión de azar, sino el resultado de la selección. La selección es un proceso que consta de dos fases, la primera etapa de las cuales es la producción de variabilidad. En cada generación, según Darwin, se genera una enorme cantidad de variabilidad: Darwin no conocía el origen de esta variación, que no

39
se esclarecería hasta después de la aparición de la genética como ciencia. Todo lo que tenia era un conocimiento empírico de la reserva, aparentemente inagotable, de grandes y pequeñas diferencias intraespecíficas.
La segunda etapa es la selección a través de la supervivencia en la lucha por la existencia. En la mayoría de especies animales y vegetales, una sola pareja de progenitores llega a procrear millares, sino millones, de descendientes. La lectura de Thomas Malthus reveló a Darwin que solamente muy pocos de ellos podían sobrevivir. ¿Qué descendientes tienen una probabilidad mayor de sobrevivir? Serian aquellos individuas que presentaran la combinación de caracteres más idónea para hacer frente al ambiente, entendiendo por tal el clima, los competidores y los enemigos; ellos tendrían una probabilidad mayor de sobrevivir, reproducirse y dejar descendientes, y sus caracteres pasarían, por tanto, al siguiente ciclo de selección. La idea de un mundo en evolución, que sustituía a la de un mundo en evolución, que sustituía a la de un mundo estático, la aceptaron casi sin excepciones todos los científicos serios, incluso antes de la muerte de Darwin, ocurrida en 1882; quienes aceptaban la evolución, aceptaron también la idea de comunidad de descendencia (aunque algunos de ellos insistieron en excluir al hombre de este linaje común). Sin embargo, los otros dos postulados de Darwin corrieron una suerte muy distinta, pues fueron dura y agriamente combatidos por un elevado número de estudiosos y eruditos durante los 50-80 años siguientes.
Uno de estos postulados era la idea de gradualismo. Ni siquiera T.H. Huxley, a quien se le llamaría el “bulldog de Darwin” por su vigorosa actitud como abanderado de la mayoría de puntos de la nueva teoría, acepta el origen gradual de los tipos superiores ni de las nuevas especies: Huxley, por su parte propugnaba un origen saltacional. El saltacionismo se hizo popular también entre los biólogos de la categoría de Hugo de Vries, uno de los redescubridores de las leyes de la herencia de Gregor Mendel. En 1901, De Vries propuso una teoría según la cual nuevas especies se originaban por mutación. Todavía en 19040 el genético Rchard B.G. Goldschimidt defendía las “mutaciones sistemáticas” como mecanismo de explicación del origen de los nuevos tipos superiores. Con el tiempo se desarrollaron tres avances que dieron al traste con las teorías saltacionistas. Fue uno la adopción gradual de una nueva actitud hacia el mundo físico y su variabilidad.
Desde la época de Platon el punto de vista dominante había sido lo que el filósofo Karl Popper denomino “esencialismo”: según dicha teoría el mundo estaría integrado por un número limitado de esencias invaginares (las ideas de Platón). De las que las manifestaciones variables del mundo visible serian meros reflejos incompletos e imprecisos. Desde este punto de vista solo podría producirse un autentico cambio a través del origen de una nueva esencia, ya fuera por creación o mediante un salto espontaneo (mutación).
Las clases de objetos físicos estaban compuestas por entidades idénticas, y las constantes físicas eran invaginares bajo condiciones idénticas, por lo que (en el siglo XIX) no existía conflicto entre las matemáticas o las ciencias físicas y la filosofía esencialista.
La biología requería una filosofía distinta. Los organismos vivientes se caracterizan por la singularidad; cualquier población de organismos consta de individuos únicos y distintos. Desde un “punto de vista poblacional”, los valores medios son abstracciones; solo tienen la realidad las variantes individuales. La importancia de la población reside en constituir un acervo de variaciones (un acervo génico, en el lenguaje de la genética). La perspectiva poblacional hace posible la evolución gradual; enfoque que es el que hoy domina todos los aspectos de la teoría de la evolución.
El segundo avance que obligaría a abandonar el saltacionismo fue el descubrimiento de la inmensa variabilidad de las poblaciones y el reconocimiento de que una gran variedad de factores genéticos discontinuos pueden manifestarse en una variación continua del organismo, siempre que haya suficiente número de los mismos y que las discontinuidades entre ellos sean lo bastante pequeñas. El tercer avance fue la demostración que hicieron los naturalistas de que los procesos de la evolución gradual son totalmente capaces de explicar el origen de discontinuidades, tales como nuevas especies y nuevos tipos y el origen de innovaciones evolutivas tales como las alas de las aves o los pulmones de los vertebrados.
La otra idea de Darwin a la que se resistieron durante mucho tiempo la mayoría de los biólogos y filósofos fue la selección natural. Al principio, muchos la habían rechazado porque no era determinista y, por tanto predictiva, en la línea de la ciencia del siglo XIX. ¿Cómo podía una “ley natural”, cual la selección natural, ser enteramente una cuestión de

40
azar? Otros atacaron su “materialismo craso”. En el siglo XIX atribuir la armonía del mundo viviente resultado arbitrario de la selección natural era socavar el “argumento del proyecto” de los teólogos naturales, según el cual la existencia de un Creador podría inferirse de la belleza del proyecto de sus obras. Quienes rechazaron la selección natural desde un punto de vista religioso o filosófico, o simplemente debido a que les parecía un proceso excesivamente azaroso como para explicar la continuidad de la evolución, postularon durante muchos años otras alternativas con nombre tales como ortogénesis, nomogénisis, aristogénesis o el “punto omega” de Teilhard, todas ellas basadas en una tendencia interna o impulso hacia la perfección o el progreso. Todas estas teorías eran finalistas: postulaban algún tipo de teología cósmica, de propósito o programa.
Los defensores de las teorías teleologícas, pese a todos sus esfuerzos, han sido incapaces de encontrar ningún mecanismo (excepto los sobrenaturales) que pueda explicar el finalismo que postulan. Hoy en día, la posibilidad de que pueda existir algún mecanismo de este tipo esta totalmente descartada, merced a los hallazgos de la biología molecular. Tal como señalo Jacques Monod con particular énfasis, el material genético es constante; solo puede cambiar por medio de mutación. Las pruebas aportadas por la paleontología han contribuido también a rebatir las teorías finalistas, como ha demostrado claramente George Gaylord Simpson. Cuando se estudia detenidamente la tenencia evolutiva de un carácter, por ejemplo la tendencia hacia un mayor tamaño del cuerpo o de los dientes, dicha tendencia resulta incoherente, ya que cambia de dirección repetidamente, y, a veces, incluso se invierte. La frecuencia de extinción en cada periodo geológico es otro argumento poderoso contra cualquier tendencia finalista hacia la perfección.
No es difícil rebatir la objeción al supuesto componente aleatorio de la selección natural. El proceso no es, en absoluto, cuestión de puro azar. Aunque surgen variaciones por procesos aleatorios, se clasifican dentro de la segunda etapa del proceso: la selección por la supervivencia que es, en gran medida, un factor antiazar. Y si también es cierto que parte de la evolución es resultado del azar, en la actualidad sabemos que los procesos físicos presentan, en general, un componente probabilistico mucho mayor de lo que se reconocía hace 100 años. A pesar de ello. ¿Puede la selección natural explicar la larga
progresión evolutiva hasta las plantas y animales “superiores”, incluido el hombre, desde el origen de la vida hace entre tres y cuatro mil millones de años? ¿De que manera puede la selección natural explicarnos solamente la supervivencia diferencial y los cambios adaptativos intraespecificos, sino también la aparición de nuevas especies, diversamente adaptadas? De nuevo fue Darwin quien sugirió la respuesta adecuada. Un organismo compite no solo con otros organismos de la misma especie, sino también con individuos de otras especies. Una nueva aspiración o mejora fisiológica general convertirá al individuo y a sus descendientes en competidores interespecificos más fuertes, y, por tanto, contribuirá a la diversificación y a la especialización. Dicha especialización conduce a menudo a un callejón sin salida, como es, por ejemplo, el caso de la adaptación a la vida cavernícola o en aguas termales. Sin embargo, muchas especializaciones, sobre todo aquellas que se adquirieron al principio de la historia evolutiva, abrieron niveles completamente nuevos de radiación adaptativa. Estos iban desde la invención de las membranas y de un núcleo celular organizado [véase, más adelante, “La evolución de las células primitivas” por J. William Shopf] y la agregación de células para formar organismos pluricelulares, hasta la aparición de sistemas nerviosos centrales altamente desarrollados y la invención del cuidado prolongado de la prole.
La evolución, como Simpson puso de manifiesto, es oportunista en extremo: favorece cualquier variación que confiera al individuo una ventaja en la competencia con los demás miembros de su población o sobre individuos de otras especies. A lo largo de miles de millones de años este proceso ha venido funcionando automáticamente, impulsando lo que nosotros llamamos proceso evolutivo. Ningún programa controlaba o dirigía esta progresión; ha sido el resultado de la decisión momentánea de la selección natural. El desconocimiento de Darwin en lo concerniente al origen de la variabilidad genética. Que constituye la materia prima para la selección natural, dejo un gran vacío en su argumentación; dicha laguna quedaría subsanada por la ciencia de la genética. En 1865, Mendel descubrió que los factores que transmiten la información hereditaria son unidades discretas transmitidas a la descendencia por cada progenitor, unidades que se preservan puras y se mezclan de nuevo en cada generación. Darwin no llegó a conocer nunca los hallazgos de Mendel,

41
ignorados durante mucho tiempo hasta el redescubrimiento de los mismos en 1900.
Hoy sabemos que el Adn del núcleo celular esta organizado en números genes que se autorreplican (unidades hereditarias mendelinas). Los cuales pueden mutar para formar alelos distintos, o formas alternativas. Existen genes estructurales, que codifican la información para la síntesis de proteínas específicas, y genes reguladores, que activan y desactivan a los genes estructurales. Un gen estructural mutado puede codificar para una proteína variante, comportando un carácter variante. Los genes se disponen en los cromosomas ordenadamente y pueden recombinarse entre sí durante el proceso celular de la meiosis, que precede a la formación de células y germinales en las especies dotadas de reproducción sexual. La diversidad de genotipos (dotaciones completas de genes) que pueden producirse durante la meiosis es tan elevada que rebasa casi lo inimaginable; gran parte de esa diversidad se conserva en las poblaciones a pesar de la selección natural.
Por extraño que nos parezca, los primeros seguidores de Mendel no aceptaron la teoría de la selección natural. Eran esencialistas y saltacionistas, que veían en la mutación la fuerza impulsora verosímil de la evolución. El panorama empezó a cambiar con el desarrollo de la genética de poblaciones en la década de 1920. Y, a lo largo de las dos décadas subsiguientes, se llegaría a una síntesis, que se expreso y comento ampliamente a través de los libros escritos por Theodosius Dobzhansky, Julian Huxley, Bernhard Rensch, Simpson G.Ledyard Stebbins y el autor de esta introducción. La nueva “teoría sintética” de la evolución ampliada por Darwin a la luz de la teoría cromosomica de la herencia, de la genética de poblaciones, la idea biológica de especie y otras nociones de biología y paleontología. La nueva síntesis se caracterizo por el rechazo total de la herencia de los caracteres adquiridos, un énfasis en la condición gradual de la evolución y el reconocimiento de la importancia decisiva de la elección natural.
La comprensión de los procesos evolutivos, lograda gracias a la teoría sintética, tuvo una profunda resonancia en toda la biología. Hizo comprender que cualquier problema biológico plantea una cuestión evolutiva; que, con respecto a cualquier estructura, función o proceso biológico, es lícito preguntarse ¿Por qué está ahí? ¿Qué ventaja selectiva tenia cuando se adquirió? Preguntas todas de enorme incidencia de cualquier sector
biológico y, de manera partícula, en biología molecular en etología y ecología.
A los filósofos y los físicos, y al mismo hombre de la calle, les sigue constando entender la moderna teoría de la evolución orgánica a través de la selección natural. A riesgo de repetir algunos puntos que ya he tratado en su contexto histórico, permítaseme subrayar los rasgos especiales de la teoría en boga, prestando particular atención a aquello que se distingue a la evolución cósmica y de otros procesos tratados por físicos. La evolución a través de la selección natural es un proceso (vuelvo a repetirme) lección. Esa constelación de presiones cambia con las estaciones, a lo largo de los años y de un sitio geográfico a otro.
La segunda fase de la selección natural, la selección propiamente dicha, constituye un principio intrínseco de regulación. En una población compuesta de miles o millones de individualidades, ciertos individuos tendrán dotaciones génicas mejor dispuestas para hacer frente al conjunto de presiones ecológicas dominantes. Esos individuos poseerán, estadísticamente hablando, una probabilidad de supervivencia y de procreación mayor que el resto de los miembros que componen la población.
Esta segunda fase de la selección natural determina la dirección del proceso evolutivo, al aumentar la frecuencia de los genes y constelaciones de genes adaptados a un tiempo y lugar determinados, al aumentar la eficacia biológica, al promover la especialización y al dar origen a radiaciones adaptativas y a todo lo que, de u modo impreciso, se denomina progreso evolutivo.
Con otras palabras, la evolución por selección no es ni un fenómeno fortuito ni tampoco un fenómeno determinístico, sino un proceso bifásico que combina las ventajas de uno y otro. Como escribiera el pionero de la genética de poblaciones Sewall Wright: “El proceso darwiniano de continua interrelación entre un proceso aleatorio y otro selectivo no es un camino medio entre el puro azar y el determinismo absoluto, sino una va de consecuencias entera y cualitativamente diferentes de la de ambos”.
No me consta que ningún darwinista cuestione el hecho de que los procesos de la evolución orgánica sean coherentes con las leyes de la ciencias físicas; pero carece de sentido afirmar que la evolución biológica “se reduce” a las leyes de la física. La evolución biológica resulta de procesos específicos que actúan sobre sistemas específicos, y cuya explicación

42
solo tiene razón de ser a nivel de complejidad de tales procesos y tales sistemas. Y la teoría clásica de la evolución tampoco ha reducido a una “teoría molecular de la evolución”, opinión que se funda en ciertas definiciones reduccionistas de la evolución del tipo de la siguiente “cambio en las frecuencias génicas de poblaciones naturales”. Esa definición reduccionista omite los aspectos cruciales de la evolución: cambios en la diversidad y en la adaptación. (En cierta ocasión le di un terrón de azúcar a un mapache en el zoo. Se lo llevo corriendo a la pila de agua y lo lavo vigorosamente, hasta que se deshizo por completo. Ningún sistema complejo debería eliminarse hasta el punto de no dejar nada valido del mismo.
Una vez alcanzada la nueva síntesis en las décadas de 1930 y 1940, algunos discrepantes de la evolución se preguntaron si no había llegado ya la hora final de la investigación en ese sentido, y si no se había respondido ya a todas las cuestiones. La contestación a ambas preguntas es rotundamente no; según queda puesto de manifiesto en el aumento exponencial del número de publicaciones sobre biología evolutiva. Mencionare algunos problemas que suelen concentrar el interés de los investigadores en ese campo.
Uno de los temas de mayor dedicación investigadora es la función del azar. Ya en 1871 se había sugerido que quizá solo del cambio evolutivo se debiera a la selección, en tanto que la porción destacada restante, sin casi todo el cambio, obedeciera a la variación accidental, a lo que ahora conocemos por mutaciones “neutras”. Esta insinuación se ha venido repitiendo muchas veces desde entonces. El problema adquirió una nueva dimensión cuando la técnica de la electroforesis permitió detectar pequeñas diferencias en la composición de un encima particular en una amplia muestra de individuos; ello revelaba el enorme grado de variabilidad alélica, ¿Qué fracción de esa variabilidad es “ruido” evolutivo y que fracción depende de la selección? ¿Por qué procedimiento podemos partir de la variabilidad en alelos neutros y alelos dotados de cierta significación?
El descubrimiento por parte de la biología molecular de la existencia de genes reguladores y genes estructurales plantea nuevos interrogantes en el campo de la evolución; ¿Es idéntico el ritmo de evolución en ambos tipos de genes? ¿Son ambos igualmente susceptibles de selección natural? Por lo que respecta a la especiación o al
origen de grupos taxonómicos superiores, ¿es un tipo de genes más importante que el otro? (Por ejemplo, los genes estructurales del chimpancé y del hombre son notablemente parecidos; serán entonces los genes reguladores los responsables de la mayoría de las diferencias entre el chimpancé y el hombre?) ¿Hay tal vez, otros tipos de genes?
El problema que más caro le era a Darwin, el de la multiplicación de las especies vuelve a ser objeto de investigación. En ciertos grupos de organismos. Como el de las aves por ejemplo, las nuevas especies parecen originarse exclusivamente por especiación geográfica: a través de la reestructuración genética de poblaciones que se hallaban aisladas del resto de la especie, en una isla por ejemplo. En las planta y en otros grupos animales, muy pocos por cierto, ocurre un tipo distinto de especiación por poliploidia (duplicación de la dotación cromosómica), ya que los individuos poliploides quedan inmediatamente aislados, desde el punto de vista reproductor, de sus progenitores. Un tercer tipo de especiación es el llamado especiación “simpátrica”, que se observa en parásitos o en insectos adaptados a vivir sobre un planta huésped determinada. Puede suceder que una nueva especie huésped sea colonizada accidentalmente, y que los descendientes de los inmigrantes, ayudados quizá por cierta combinación favorable de sus genes, lleguen a constituir una colonia prospera. En este caso existirá una fuerte selección de los genes que favorezcan la reproducción con otros individuos que vivan sobre la nueva especie huésped, de suerte que las condiciones favorezcan el desarrollo de una nueva raza adaptada al nuevo huésped y, con el tiempo, a una nueva especie que sea especifica del hospedante en cuestión. La frecuencia de la especiación simpátrica sigue siendo motivo de polémica, al igual que continua debatiéndose también el papel respectivo de genes y cromosomas en el proceso de espición.
En pocos campos de la biología ha sido tan fructífera la introducción de la perspectiva evolutiva como en el sector de la biología del comportamiento. Los etólogos clásicos habían puesto relieve que ciertas pautas de comportamiento, como las exhibiciones de cortejo, podían reflejar unas relaciones taxonómicas tan ajustadas como los propios caracteres estructurales.
Se han realizado clasificaciones basadas en el comportamiento que concuerdan notablemente con las configuradas a partir de la estructura; y los datos etólogos han

43
mostrado ser frecuentemente decisivos en los casos que en las pruebas morfológicas no resultaban clarificadoras. Y lo que es más importante, se ha demostrado que a menudo –o quizá siempre- la conductora es el marcapasos de la evolución. Un cambio en el comportamiento, la selección de un nuevo hábitat o fuente alimentaria por ejemplo, desencadena nuevas presiones selectivas y puede conducir a importantes cambios adapatativos. Hay casi plena certeza de que algunos de los principales acontecimientos de la historia de la vida, como la conquista del medio terrestre o el medio aéreo, comenzó a raíz de modificaciones en el comportamiento. Las presiones de selección que robustecen esos avances evolutivos están siendo ahora objeto de particular atención.
La percepción de que el mundo no es estático, sino que se halla en continuo cambio y de que nuestra propia especie es el producto de la evolución ha incidido inevitable y profundamente en el entendimiento humano. Sabemos ya que la línea evolutiva a la que pertenecemos surgió a partir de antepasados de naturaleza antropoide en el curso de millones de años, y que las etapas fundamentales de ese proceso acaecieron aproximadamente en el último millón de la historia del planeta. Conocemos asimismo que la selección natural tuvo que ser la responsable de ese avance. ¿Qué es lo que esos acontecimientos pretéritos nos permiten predecir respecto al futuro de la humanidad? Puesto que no existe ningún elemento teleologico en la evolución orgánica, ni se heredan los caracteres adquiridos, la selección constituye obviamente el único mecanismo potencialmente capaz de influir en la evolución biológica del hombre.
Esa conclusión plantea un dilema. La eugenesia, o selección deliberada, podría entrar en conflicto con determinados valores humanos respetables. Aun cuando no hubiera objeciones de índole moral, no siempre se dispone de la información necesaria sobre la que debe fundarse tal selección. No se sabe casi nada sobre el componente genético de rasgos humanos no físicos. Hay innumerables, y multiformes tipos de seres humanos “buenos”, “útiles” o adaptados. Aun cuando en un momento consiguiéramos seleccionar un conjunto de caracteres ideales, los cambios que generan en la sociedad los adelantos de la técnica se dan con tal rapidez que no podríamos predecir cual seria el conjunto de talentos que habría de conducir, en el futuro, a la más armoniosa de las sociedades humanas. Dobzhansky decía que la
humanidad sigue en estado evolutivo, pero no sabemos, desde una óptica biológica, a donde se dirige.
Hay, empero, otro tipo de evolución: la evolución cultural. Se trata de un proceso peculiarmente humano por el que el hombre, hasta cierto punto, se amolda y se adapta a su en torno. (En tanto que las aves, los murciélagos y los insectos llegaron a volar en virtud de una evolución genética que duro millones de años, Dobzhansky señalaba que “el hombre se ha convertido en el volador más poderoso por propia construcción de maquinas planeadoras, no por reconstrucción de su genotipo”.) La evolución cultural es un proceso mucho más veloz que la evolución biológica.
Uno de sus rasgos es la capacidad fundamental (y de viejo sabor lamarckista) de los seres humanos para evolucionar culturalmente a través de la transmisión, de generación, de la información aprendida, en la que se encuadran los valores morales –y los inmortales-. Seguramente podrán registrase todavía grandes progresos en este terrenos, si consideramos el bajo nivel de los valores, si consideramos el bajo nivel de los valores morales que manifiesta hoy la humanidad. Aun cuando no hubiera manera de condicionar nuestra propia evolución biológica, existiría la posibilidad cierta de influir en nuestra evolución cultural y moral. Hacerlo en direcciones que resulten adaptativas para toda la humanidad constituiría un objetivo evolutivo realista, por más que sea cierta la existencia de límites a la evolución cultural y moral en una especie humana que no haya sido manipulada desde el punto e vista genético.
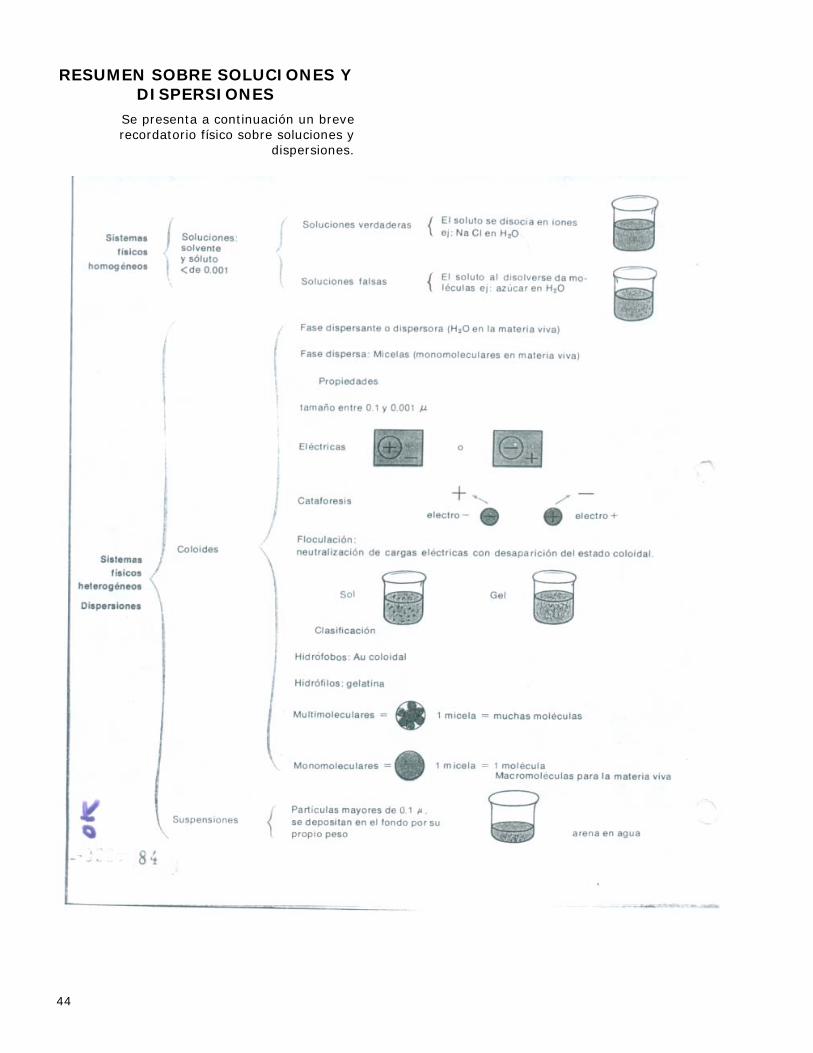
44
RESUMEN SOBRE SOLUCIONES Y DISPERSIONES
Se presenta a continuación un breve recordatorio físico sobre soluciones y
dispersiones.
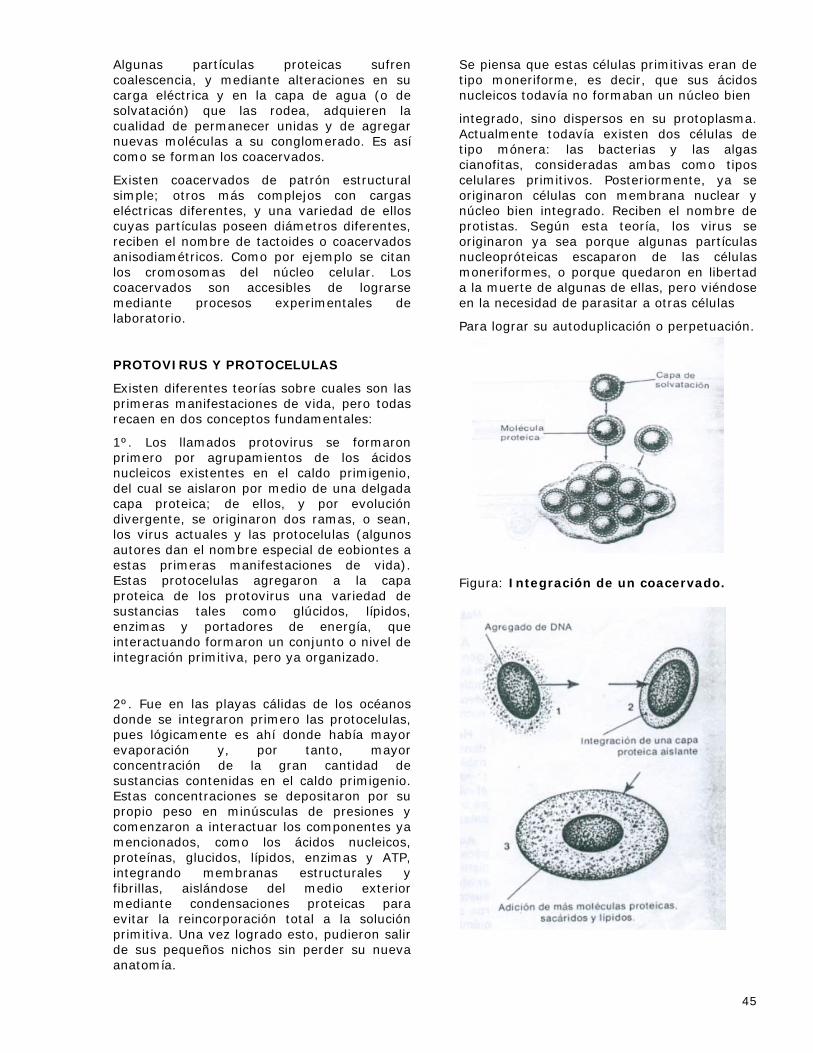
45
Algunas partículas proteicas sufren coalescencia, y mediante alteraciones en su carga eléctrica y en la capa de agua (o de solvatación) que las rodea, adquieren la cualidad de permanecer unidas y de agregar nuevas moléculas a su conglomerado. Es así como se forman los coacervados.
Existen coacervados de patrón estructural simple; otros más complejos con cargas eléctricas diferentes, y una variedad de ellos cuyas partículas poseen diámetros diferentes, reciben el nombre de tactoides o coacervados anisodiamétricos. Como por ejemplo se citan los cromosomas del núcleo celular. Los coacervados son accesibles de lograrse mediante procesos experimentales de laboratorio.
PROTOVIRUS Y PROTOCELULAS
Existen diferentes teorías sobre cuales son las primeras manifestaciones de vida, pero todas recaen en dos conceptos fundamentales:
1º. Los llamados protovirus se formaron primero por agrupamientos de los ácidos nucleicos existentes en el caldo primigenio, del cual se aislaron por medio de una delgada capa proteica; de ellos, y por evolución divergente, se originaron dos ramas, o sean, los virus actuales y las protocelulas (algunos autores dan el nombre especial de eobiontes a estas primeras manifestaciones de vida). Estas protocelulas agregaron a la capa proteica de los protovirus una variedad de sustancias tales como glúcidos, lípidos, enzimas y portadores de energía, que interactuando formaron un conjunto o nivel de integración primitiva, pero ya organizado.
2º. Fue en las playas cálidas de los océanos donde se integraron primero las protocelulas, pues lógicamente es ahí donde había mayor evaporación y, por tanto, mayor concentración de la gran cantidad de sustancias contenidas en el caldo primigenio. Estas concentraciones se depositaron por su propio peso en minúsculas de presiones y comenzaron a interactuar los componentes ya mencionados, como los ácidos nucleicos, proteínas, glucidos, lípidos, enzimas y ATP, integrando membranas estructurales y fibrillas, aislándose del medio exterior mediante condensaciones proteicas para evitar la reincorporación total a la solución primitiva. Una vez logrado esto, pudieron salir de sus pequeños nichos sin perder su nueva anatomía.
Se piensa que estas células primitivas eran de tipo moneriforme, es decir, que sus ácidos nucleicos todavía no formaban un núcleo bien
integrado, sino dispersos en su protoplasma. Actualmente todavía existen dos células de tipo mónera: las bacterias y las algas cianofitas, consideradas ambas como tipos celulares primitivos. Posteriormente, ya se originaron células con membrana nuclear y núcleo bien integrado. Reciben el nombre de protistas. Según esta teoría, los virus se originaron ya sea porque algunas partículas nucleopróteicas escaparon de las células moneriformes, o porque quedaron en libertad a la muerte de algunas de ellas, pero viéndose en la necesidad de parasitar a otras células
Para lograr su autoduplicación o perpetuación.
Figura: Integración de un coacervado.

46
Figura: Hipótesis sobre la formación de eobiontes: de protovirus a protocélula.
HETEROTROFISMO Y AUTOTROFISMO
A través de las anteriores consideraciones sobre el origen paulatino de la vida mediante organizaciones cada vez más complejas de la materia no viviente durante millones y millones de años, se supone que los primeros organismos vivos así formados fueron muy simples. Sobre su naturaleza nutricional existen dos teorías: la heterótrofa y la autótrofa.
HETERÓTROFA: Esta hipótesis supone que habiendo tal abundancia de materiales en el caldo primigenio, es más razonable que un organismo sencillo pueda con más facilidad tomar sus partículas nutricias ya integradas que sintetizarlas él mismo. La diferente manera de tomar estos alimentos originó tres subtipos de nutrición heterótrofa, de las que se trato anteriormente: el saprofitismo, parasitismo y holotrofismo.
AUTÓTROFA: Supone que los primeros organismos eran capaces de sintetizar sus propios alimentos utilizando para ello distintas fuentes de energía, tales como la luz solar, la cual existe en abundancia; o bien, energía química a partir de sustancias inorgánicas, como el S o el Fe. Así se originaron dos subtipos de nutrición autótrofa: la fotosíntesis y la quimiosíntesis. La oposición a esta teoría señala el hecho de que las reacciones que se lleven a cabo en el autotrofismo, son demasiado complejas para ser llevadas a cabo por organismos tan simples como se supone que fueron los primeros vivientes.
Figura: Hipótesis sobre la integración de las primeras manifestaciones de vida: protocélula y evolución radiada.
CENOZOICA: Calculada en 75 millones de años y dividida en periodos Terciarios y Cuaternario con sus correspondientes subdivisiones. Predominio de los angiospermas, aunque se desconoce su filogenia exacta. A la extinción, por superespecialización, de los grandes reptiles, se sucede el predominio de sus descendientes mamíferos con la consecuente evolución radiada de la cual se conservan tres ramas principales, las dos primeras con escasos representantes: prototeria, metateria y euteria o verdaderos placentados, con doce grandes grupos, uno de los cuales (insectívoros) origino un tronco que se adaptó a la vida arbórea (primates) y del cual evolucionaron los prosimios, los monos del Nuevo y Viejo Continentes, los antropoides o grandes monos y finalmente los hominidos con su serie de los llamados erróneamente eslabones perdidos que en realidad son muestras de poblaciones de una especie politípica (con subespecies), tal y como es la humana.

47
Figura: Presencia de fósiles para identificación de capas terrestres.
FÓSILES
Los datos proporcionados en los temas anteriores se basan principalmente en las mediciones hechas a través de la desintegración natural de los materiales radiactivos que se encuentran en los diferentes estratos terrestres, y en la observación de los fósiles en ellos embebidos que forman una documentación monumental sobre la evolución de la vida en las diferentes eras geológicas.
Un fósil es la traza visible de un organismo que vivió en el pasado geológico. Un hecho incontrovertible que podemos palpar con nuestras manos y observar con nuestros propios ojos. Rinde una información valiosa sobre el lugar geográfico donde se encontró, el tipo de rocas que lo albergaron, así como su edad geológica y el conjunto de otras poblaciones de fósiles con las cuales convivió en su época.
Su formación sigue varias normas entre las cuales cuentan una serie de accidentes oportunos, como el de que el organismo quedo enterrado antes de que se descompusiera, aunque usualmente se conservan mejor las partes duras formadas por materiales inorgánicos, residentes, tales como los exo y endo-esqueletos y restos celulosicos. Hay veces que los huecos dejados por las partes blandas son rellenados por materiales cristalizables provenientes del suelo (sílice, carbonatos), los cuales forman una especie de molde del organismo original. En otros casos, no se conserva el organismo en si, pero si su huella o impresión dejada en alguna parte blanda del terreno que se endurece y conserva después. Las filtraciones de agua y cristalizaciones desempeñan un papel importante en la fosilización.
Con el transcurso del tiempo, nuevas capas se depositan sobre los fósiles que así desaparecen hasta que las fuerzas erosivas (paso de ríos a través de terrenos de sedimentación blanda, como el Gran Cañón; fuertes temblores de tierra como en los Andes Chilenos, o bien, el viento y aun accidentes provocados por la mano del hombre, como la construcción de carreteras o puentes) los ponen de manifiesto. Su recolección debe ser hecha por expertos para conservar lo mejor posible su testimonio y evitar confusiones constructoras inorgánicas que por su aspecto han recibido el nombre de “pseudofosiles”. Estas rocas también deben diferenciarse e los “gastrolitos” (inclusiones pétreas del tubo
digestivo de los dinosaurios) y de los “coprolitos”, excrementos fósiles de animales extintos.
Hay que mencionar casos muy raros de preservación total, como el de los fósiles de los mamuts congelados hallados en Siberia y Alaska con antigüedad de 25,000 años, y un rinoceronte completo incluido en asfalto (Polonia). En las regiones secas sudamericanas se han encontrado restos momificados (desecación) en algunas cuevas. Algunas resinas vegetales también se han fosilizado y forman el llamado “ámbar”.
Los hombres primitivos dejaron también su testimonio bajo el aspecto de sus restos fósiles o de sus utensilios, herramientas y pinturas.
Figuras: Resina fósil (ámbar) con un insecto atrapado en ella.

48
Figura: Evolución de los mamíferos a partir e un ancestro reptiliano Tecodonto (1) Paleoceno: Phenacodus (2) Eoceno: Coryphodon (3) Uintaherium (4) Pleistoceno: Megatherium (5) Nothrotherium (6) Megaceros (7).

49
Figura: Restos de un Mamut parcialmente conservado en los hielos siberianos.
Figura: Identificación y reconstrucción de
fósiles de animales y vegetales.

50
EVOLUCIÓN La idea filosófica de la evolución no es una novedad. Ya en la época griega se hablaba de ella, aunque oscurecida por las metamorfosis maravillosas tan en boga en las creencias de ese tiempo. Sin embargo, se expreso que los animales existentes provenían de otras formas anteriores a ellos y que había variedad de individuos dentro de una especie, principalmente en los sometidos a la domesticidad.
Estas ideas sufrieron un franco estancamiento debido a la opinión expresada por Platón referente a lo que el llamaba el Eidos: forma tangible y concreta de la idea que no admite variación ni alteración alguna, lo que consideraban como una imperfección que debía desecharse de inmediato.
Aristóteles siguió este criterio al hacer la primera clasificación tipológica de los animales por él conocidos y sus ideas influenciaron durante mucho tiempo a los filósofos de los siglos XVII, XVIII y XIX.
A medida que avanzaron los conocimientos biológicos, se hizo aparente el cambio de los seres vivientes a través del tiempo, especialmente en las series paleontológicas que hicieron emitir a Cuvier una débil teoría sobre cataclismos sucesivos a los que seguirían otras tantas creaciones que explicarían la variación observada.
La primera hipótesis evolutiva corresponde a Jean Baptiste Lamarck que apoyo a su teoría en varios hechos, unos inadecuados y confusos y otros con algún acierto:
1-. Principio del automejoramiento de las especies.
2-. Uso y desuso de los órganos que provocaban la hipertrofia o atrofia de los mismos.
3-. Herencia de los caracteres adquiridos.
4-. Negación total de la teoría de las creaciones sucesivas.
Consideró a la especie no como una serie, sino como una ramificación.
La biología actual reconoce el mérito del trabajo de Lamarck al enfrentarse con un medio totalmente hostil que desprecio su trabajo, pero en el que abrió una brecha a las ideas evolucionistas, aunque los mecanismos por él invocado no fueran totalmente acertados.
Muchas de las teorías evolucionistas de los primeros tiempos y las posteriores adolecen del defecto e tratar de basar todo el proceso evolutivo en un solo hecho, por lo que se les dio el titulo de teorías monísticas.
El primero en reconocer que la evolución era debida un equilibrio de diferentes fuerzas en conflicto, fue Charles Darwin (1809-1882).
Sus ideas abrieron campo a las actuales teorías llamadas sintéticas que consideraban la evolución como un hecho demasiado complejo para residir en un solo factor, e invocan varios de ellos para explicarlo.
La teoría evolutiva de Darwin es la que más se acerca al concepto actual de evolución y es interesante conocer la cadenas de circunstancias que contribuyeron a orientar el pensamiento de Darwin por este sendero. Conestudios previos sobre ciencias naturales y geología, Darwin visitó asiduamente los museos. La diversidad de los seres vivos era para él conocida bajo el aspecto de especímenes de laboratorio o museo. A los 22 años se embarco como naturalista en el velero Beagle en un viaje de 5 años que le llevo alrededor del mundo. El primer impacto que recibió Darwin en su escala en Brasil, fue que los seres vivos en su medio natural diferían de los disecados, en que parecían formar parte del todo biológico donde se encontraban, por lo que debían considerarse no como individuos aislados sino como partes integrantes del conjunto de seres vivos.
La segunda observación fue que la vida se desarrolla en una competencia no solo interespecífica, sino también intraespecífica. A esto se le denomino lucha por la existencia, en la cual hay siempre una parte ganadora y otra perdedora de la población; la diferencia entre los ganadores y perdedores recibió de Darwin el nombre de selección natural, por medio de la cual perduran los más fuertes, aptos y mejor adaptados al medio.
En su escala en las Islas Galápagos, pudo percibir la variación geográfica al observar especie aisladas de las del continente, que
Diferían en varias características morfológicos, por lo que dedico especial atención a tortugas y pájaros pinzones, cuyo pico mostraba una serie de adaptaciones a los distintos regímenes alimentación.
La observación del fósil Glyptodon y los actuales armadillos favoreció más las ideas que, deducidas de sus cuidadosas observaciones y datos acumulados, iban juntamente en su mente. Casi en contra de su voluntad, se vio Darwin obligado a aceptar el

51
hecho del cambio evolutivo de las especies y se dedico con verdaderas honradez y autoexistencia científica a acumular más y más datos a favor de esta teoría.
Dos trabajos fueron de ayuda para él: uno de Lyell sobre los cambios geológicos de la corteza terrestre en las diversas épocas, lo que le hizo pensar que estos cambios debieron haber influido en el medio imperante de los seres vivos y que estos, a su vez, debieron haber cambiado también. El otro trabajo fue el que decía que la especie humana se reproducía en progresión geométrica y los alimentos solo en progresión aritmética; orientando sus observaciones hacia este punto, Darwin puso en claro que, si bien las especies tenían una mayor reproductividad, había factores que frenaban este crecimiento y mantenían más o menos igual el número de pobladores de la especie humana (selección natural, lucha por la existencia).
Estaba Darwin ando los últimos toques a su trabajo para presentarlo, cuando recibió del naturalista Alfred Russel Wallace, quien hacia investigaciones en Malaya, un trabajo concebido en términos casi iguales a los suyos. Correspondiente, por lo tanto, el mérito de ambos.
La obra total de Darwin, con sus múltiples trabajos, nos hace apreciar su gran capacidad, su excelente preparación y perspicacia científica, así como la curiosidad e inquietud como naturalista.
Posteriormente a Darwin, hubo dos conocimientos de gran importancia que han contribuido mucho en el esclarecimiento de muchos fenómenos evolutivos que él no pudo explicar: la genética y la estructura de las poblaciones.
CONCEPTO DE ESPECIE
El concepto tipológico de la especie prevaleció muchos años a partir de las declaraciones fijistas de Carlos Linneo, a quien corresponde el innegable mérito de haber ordenado y clasificado en forma competente la mayoría de las especies vivientes conocidas. Según este criterio, la especie proviene tal cual es, de una creación; es inmutable y todos los individuos que la integran son semejantes entre si.
Lamarck opino que la especie es “ una colección de individuos semejantes que la generación perpetua en el mismo estado, hasta que diversas circunstancias del medio ambiente la hagan cambiar”. Además,
pensaba que en vez de ser una serie, es una ramificación.
Posteriormente se agregó la condición de que los individuos que integran una especie, deben tener “descendencia fecunda”.
Mas tarde se extendió el concepto de considerar la especie como una entidad ficticia convencional y no como el grupo natural que es, hasta que se vio que los mismos individuos limitan su especie verificando el cruzamiento entre ellos y no con ninguna otra especie, a pesar de que lo podrían hacer y que sus descendientes serian fecundos.
Así que la propiedad característica de la especie que se considera actualmente, es que sea un grupo de individuos que se crucen entre si.
Las propiedades biológicas básicas de las especies son tres:
1) Adaptación al medio físico.
2) Capacidad de coexistencia.
3) Conservación de su aislamiento reproductor.
ADAPTACIÓN AL MEDIO FISICO.
Cada especie tiene necesidades ecológicas específicas como características propias y un límite de tolerancia a la variación de ellas (temperatura, humedad, altura, etc.).
Entonces, cuando los factores ecológicos han sobrepasado al límite de tolerancia de una especie, obran como barreras que inhiben su distribución.
Así pues, existirá un medio optimo en donde se encontrará el mayor número de poblaciones de una especie que se ira haciendo menor a medida que se acerque a las barreras ecológicas limitantes.
Las poblaciones de las especies que se hallan distribuidas en estos límites soportan un grado máximo de condiciones poco favorables, muchas de ellas desaparecerán, pero hay algunas que soportan la variación del medio y son el material vivo más adecuado para iniciar una variación de donde puede seguir un cambio evolutivo. (Lám. XXXIII).
COEXISTENCIA.
En un mismo medio no pueden existir dos especies diferentes que tengan idénticas condiciones ecológicas, a no ser que este

52
medio sea muy amplio y tenga lugares o nichos vacíos.
Los competidores de una especie son aquellos que tienen las mismas necesidades ecológicas. Las especies que habitan en el mismo lugar, al mismo tiempo, se llaman especies simpátricas, y se extinguirán, respecto a sus competidores, aquellas que tengan menor potencial biótico (capacidad reproductora). Otro factor desfavorable es que una especie intervenga en la competencia en condiciones desventajosas del número de su población.
CONSERVACIÓN DEL AISLAMIENTO REPRODUCTOR.
Esta es una de las características más notables que conserva integras a las especies e impide la hidratación. El término que se utiliza para nombrar en general a las propiedades biológicas que evitan el cruzamiento interespecífico es el mecanismo de aislamiento:
1) Mecanismo (como ejemplo) anterior al cruce, tenemos:
Aislamiento por diferentes estaciones de celo o diferentes hábitos de vida que impiden que los posibles cónyuge se encuentren.

53
EL ORIGEN DE LAS ESPECIES Carlos Darwin
INTRODUCCION
DE RICHARD E. LEAKEY
SEXTA EDICION.
CAPÍTULO II
LA VARIACION EN LA NATURALEZA
Antes de aplicar a los seres orgánicos en estado natural los principios a que hemos llegado en el capitulo anterior, podemos discutir brevemente si estos seres están sujetos a alguna variación. No discutiré aquí las varias definiciones que se han dado de la palabra especie. Generalmente, esta palabra encierra el elemento desconocido de un acto distinto de la creación. La palabra variedad es casi tan difícil de definir, pero en ella se sobreentiende casi universalmente comunidad de origen, aunque esta rara vez pueda ser probada. Tenemos además lo que se llama monstruosidades, las cuales pasan gradualmente a las variedades. Por monstruosidad supongo que se entiende alguna considerable anomalía de conformación, generalmente perjudicial o inútil para la especie. Algunos autores usan técnicamente la palabra variación, en el sentido de modificación debida directamente a las condiciones físicas de vida, y se supone que las variaciones de esta clase son hereditarias; pero ¿quién puede afirmar que el enanismo de los moluscos en las aguas nauseabundas del Báltico o las plantas enanas de las cimas de los Alpes, o la piel más espesa de un animal de muy al norte no serian heredados en algunos casos durante varias generaciones por lo menos? Y siendo así, presumo que la forma seria llamada variedad. Puede dudarse de que una desviación repentina y considerable de estructura, como las que vemos ocasionalmente en estado de naturaleza. Casi todas las partes de cualquier ser orgánico están adaptadas en tal forma a las complejas condiciones de vida que parece improbable que ninguna parte haya sido producida súbitamente en su perfección, como lo es que una maquina complicada hubiera sido creada perfecta desde el principio por el hombre. En el estado domestico se producen a veces monstruosidades parecidas a estructuras que eran normales en animales muy diferentes. Así, a veces han nacido cerdos con una especie de trompa, y si algún animal salvaje del mismo género hubiera poseído trompa por naturaleza, podría haberse afirmado que nació como
monstruosidad. Pero hasta ahora, a pesar de búsquedas diligentes, no he podido encontrar casos de monstruosidades que se parezcan a estructuras que son normales en formas estrechamente vinculadas, que son las únicas que importan en esta cuestión. Si formas monstruosas de esa clase aparecen alguna vez en estado de naturaleza y son capaces de reproducirse - lo que no siempre ocurre -, su preservación, puesto que se han producido raramente y en forma aislada, dependería de circunstancias excepcionalmente favorables. Y durante la primera y las sucesivas generaciones se cruzarían también con la forma ordinaria y así se perdería inevitablemente su carácter anormal. Pero tendré que volver en un capitulo futuro sobre la preservación y perpetuación de variaciones aisladas u ocasionales.
DIFERENCIAS INDIVIDUALES.
Las muchas diferencias leves que len los descendientes de los mismos padres, o que puede suponerse que se han surgido así por ser observadas en los ejemplares de la misma especie que habitan la misma región cerrada, pueden ser llamadas diferencias individuales. Nadie supone que todos los individuos de la misma especie han sido fundidos exactamente en el mismo molde. Estas diferencias individuales tienen la mayor importancia para nosotros, porque a menudo son hereditarias, como todos estarán acostumbrados a observar, y por eso proporcionan materiales para que opere y se acumule la selección natural, de la propia manera en que el hombre acumula en una dirección dada diferencias individuales en sus productos domésticos. Esas diferencias individuales afectan generalmente a lo que los naturalistas consideran partes sin importancia; pero y podría demostrar con un largo catálogo de hechos que ciertas partes que es forzoso llamar importantes, ya sea desde el punto de vista fisiológico o desde el punto de vista de la clasificación, varían a veces en ejemplares de la misma especie. Estoy convencido de que los naturalistas más experimentados quedarían sorprendidos ante el número de casos de variabilidad, inclusive en partes importantes de la estructura, que podrían recoger de datos autorizados, como los he recogido yo durante años. Debe recordarse que los sistematizadores están muy lejos de sentirse complacidos cuando encuentran variabilidad en caracteres importantes, y que son muchos los hombres que quieren consagrarse a examinar laboriosamente orgánicos internos e importantes y compararlos en muchos

54
especimenes de una especie determinada. Nunca se habría esperado que la ramificación de los nervios principales próximos al ganglio central de un insecto habría sido variable en la misma especie; podría haberse pensado que cambios de esta naturaleza solo habían de manifestarse por lentas graduaciones; sin embargo, Sir J. Lubbock ha demostrado que el coctus (Insecto del orden de los hemípteros), presenta un grado de variabilidad en dichos nervios que casi pueden compararse con la ramificación irregular del tronco de un árbol. Puedo añadir que este naturalista filósofo, ha demostrado también que los músculos de las larvas de ciertos insectos estan lejos de ser uniformes. Los autores suelen argumentar en un circulo vicioso cuando afirman que los órganos importantes nunca varían; porque esos mismos autores (como han confesado francamente algunos naturalistas) califican de importantes virtualmente a todas las partes que no varían, y de ese modo nunca se encontrara una parte importante que varíe. Bajo el otro punto de vista se pueden encontrar de seguro muchos ejemplos.
En lo que se refiere a las diferencias individuales hay un hecho que produce extremada perplejidad: acudo a esos géneros que han sido llamados proteicos o polimorfos, en que las especies presentan un grado inusitado de variación. Con respecto a muchas de esas formas, apenas habrá dos naturalistas que estén de acuerdo sobre si han de clasificarlas como especies o como variedades. Podemos citar el rubus, la rosa y el hieracium entre las plantas, varios géneros de insectos y de moluscos braquiópodos. En la mayoría de los géneros polimorfos, algunas especies tienen caracteres fijos y definidos. Géneros que son polimórficos en un país, parece serlo también, con pocas excepciones, en los otros y haberlo sido igualmente, a juzgar por los braquiópodos, en épocas anteriores. Estos hechos desconciertan, porque parecen indicar que esta especie de variabilidad es independiente de las condiciones de vida. Me inclino a sospechar que vemos, por lo menos en algunos de esos géneros polimorfos, variaciones que no son útiles ni perjudiciales para la especie y que, por l tanto, no han sido fijadas y convertidas en definitivas por selección natural, como se explicara más adelante. Ejemplares de la misma especie presentan a menudo, como todos saben, grandes diferencias de estructura, independientemente de la variación, como ocurre en los dos sexos de diversos animales, en las dos o tres castas de hembras estériles u obreras entre los insectos, y en los estados larvales o incompletos de
muchos animales inferiores. También hay casos de dimorfismos y trimorfismos, tanto en los animales como en las plantas. Así, Mr. Wallace, que recientemente llamo la atención sobre el tema, ha demostrado que las hembras de ciertas especies de mariposas en el archipiélago Malayo a parecen regularmente bajo dos y aun tres formas conspicuamente diferentes, no relacionadas por variedades intermedias. Fritz Müller ha descrito casos análogos pero más extraordinarios en los machos de ciertos crustáceos brasileños: por ejemplo, el macho de los tanais se presenta regularmente bajo dos formas distintas; una de ellas tiene pinzas fueres y de forma diferente y la otra tiene antenas con muchos más pelos de olfato. Aunque en la mayoría de esos casos, las dos o tres formas, tanto animales como vegetales, no están ahora vinculadas por graduaciones intermedias, es probable que lo estuviesen en otro tiempo. Mr. Wallace describe, por ejemplo, cierta mariposa que presenta en la misma isla gran número de variedades relacionadas por eslabones extremos de la cadena se parecen estrechamente a las dos formas de una especie dimórfica relacionada que habita en otra parte del archipiélago Malayo. Así pasa también que, en las hormigas, las varias castas de obreras son generalmente bien distintas; pero en algunos casos, como veremos después, las castas están relacionadas por variedades sutilmente graduadas. Lo mismo se comprueba, como lo he observado personalmente, en algunas plantas dimorficas. Ciertamente al principio parece un hecho muy notable que la misma mariposa hembra tuviera el poder de producir simultáneamente tres formas de hembras distintas y una macho; y que una planta hermafrodita produjese de la misma cápsula de semilla tres formas hermafroditas distintas, con tres clases distintas de hembras y tres o aun seis clases diferentes de machos. Sin embargo, estos casos no son más que exageraciones del hecho común de que la hembra produce hijos de dos sexos que ha veces difieren extraordinariamente unos de otros.
ESPECIES DUDOSAS
Las formas que poseen en grado considerables el carácter de especies, pero que se parecen mucho a otras formas o están tan estrechamente vinculadas con ellas por graduaciones intermedias que los naturalistas no gustan de clasificarlas como especies distintas, son varios sentidos las más importantes para nosotros. Tenemos todas las

55
razones para creer que muchas de las formas dudosas y estrechamente relacionadas han conservado siempre sus caracteres durante largo tiempo; tanto tiempo, por lo que sabemos, como las buenas y verdaderas especies. Prácticamente, cuando un naturalista puede unir, por medio de eslabones intermedios, dos formas intermedias, trata a la una como variedad de la otra, clasificando a la más común, pero a veces a la primera descrita, como especie y a la otra como variedad de ella. Pero a veces surgen casos más difíciles, que no he de enumerar aquí, para decidir si conviene o no clasificar una forma como variedad de otra, aun cuando estén estrechamente relacionadas por eslabones intermedios; y el carácter híbrido, según se supone generalmente, de la forma intermedia, no ha de resolver siempre la dificultad. En muchos casos, empero, una forma es clasificada como variedad de otra, no porque hayan sido realmente encontrados los eslabones intermedios, sino porque la analogía lleva al observador a suponer que existen ahora o que pueden haber existido antes, y con eso queda abierto una ancha puerta a la duda y a las conjeturas.
Por consiguiente, para determinar si una forma debe ser clasificada como especie o como variedad, la única guía parece ser la opinión de naturalistas de sano juicio y amplia experiencia. Sin embargo, en muchos casos debemos decidir por mayoría de opiniones, porque son pocas las variedades bien señaladas y bien conocidas que no hayan sido clasificadas como especies al menos por algunos jueces competentes. No se puede negar que variedades de esta naturaleza dudosa están lejos de ser excepcionales. Compárense las diversas flores de Gran Bretaña, de Francia o de los Estados Unidos, trazadas por diferentes botánicos, y se vera que sorprendente número de formas han sido clasificadas por algún botánico como especie definida y por otro como mera variedad. Mr. H. C.Watson, a quien estoy profundamente agradecido por la ayuda de toda clase que me ha prestado. Me ha marcado 182 plantas británicas que generalmente son consideradas variedades, pero todas las cuales han sido juzgadas especies por algunos botánicos; y al hacer esta lista ha omitido muchas variedades de poca significación que, sin embargo, han sido clasificadas por algunos botánicos como especies, y ha omitido enteramente varios géneros muy polimórficos. Bajo la denominación de géneros, incluyendo formas muy polimórficas, Mr. Babington ha presentado 251 especies mientras Mr. Bentham da solamente 112: ¡una diferencia
de 139 formas dudosas! Entre los animales, que se unen para cada nacimiento y que tienen tanta movilidad, las formas, las formas dudosas, clasificadas como especie por un zoólogo y como variedad por otro, rara vez pueden encontrarse en el mismo país, pero son comunes en regiones separadas. ¡Cuantos de los pájaros e insectos de la América del Norte y de Europa, que difieren muy levemente unos de otros, han sido clasificados por un naturalista eminente como especie indudable y por otro como variedades o, según se les llama a menudo, razas geográficas! Mr. Wallace, en varios trabajos valiosos sobre los diversos animales, especialmente los lepidópteros, que habitan las islas del gran archipiélago Malayo, demuestra que pueden ser clasificados en cuatro categoría, a saber: como formas variables, como formas variables, como formas locales, como razas o subespecies geográficas y como verdaderas especies representativas. Las primeras – formas variables- varían mucho menos dentro de limites de la misma isla. Las formas locales son relativamente constantes y distintas en cada isla; pero cuando se comparan todas las de las de las diversas islas, las diferencias resultan tan leves y graduadas, que es imposible definirlas o distribuirlas, aunque al mismo tiempo las formas extremas son suficientemente distintas. Las razas o subespecies geográficas son formas locales completamente fijas y aisladas, pero como no difieren unas de otras por caracteres importantes y fuertemente marcados, “no hay piedra de toque posible y solo la opinión individual ha de determinar cual de ellas debe ser considerada como especie y cual como variedad”. Finalmente, las especies representativas ocupan en la economía natural de la isla el mismo lugar que las formas locales y las subespecies, pero se distinguen unas de otras por mayores diferencias que las existen entre las formas locales y las subespecies por eso son casi universalmente clasificadas por los naturalistas como especies de verdad. No obstante, es imposible dar un criterio seguro para distinguir las formas variables, las formas locales, las subespecies y las especies representativas. Hace muchos años, cuando yo comparaba y veía comparar a otros las aves de las islas vecinas del archipiélago de las Galápagos, unas con otras, y las y las mismas con las de la tierra firme americana, me sorprendió mucho notar cuan vaga y arbitraria era la distinción entre las especies y variedades. En las islitas del pequeño grupo de las de Madera hay muchos insectos que

56
son caracterizados como variedades en la admirable obra de Mr. Wollaston, pero que ciertamente serian clasificados como especies distintas por muchos entomólogos. Hasta Irlanda tiene algunos animales, ahora considerados como variedades, pero que han sido clasificados como especies por algunos zoólogos. Varios ornitólogos experimentados consideran nuestra gallineta roja británica, como nada más que una raza fuertemente marcada de una especie noruega, mientras la mayoría la clasifica como especie indudable, peculiar a gran Bretaña. Una gran distancia entre las regiones que habitan dos formas distintas induce a muchos naturalistas a considerarlas como especies definidas; pero se han preguntado, con razón, que distancia bastara: si la que media entre Europa y América es amplia, ¿será suficiente la que hay entre Europa y las Azores, o las Islas de Madera. O las canarias, o entre las varias islas de estos pequeños archipiélagos?
Mr. B. D. Walsh, distinguido entomólogo de los Estados Unidos, ha descrito lo que el llama variedades fitófagas y especies fitófagas. La mayoría de los insectos que se alimenta de vegetales viven de una u otra clase de planta o grupo de plantas; algunos se alimentan indistintamente de muchas clases, pro no por ello varían. En diversos casos, sin embargo, Mr. Walsh ha observado insectos encontrados en diferentes plantas que presentaban en su estado larval o incompleto, o en ambos estados, diferencias ligeras pero constantes en el color, el tamaño en la naturaleza de sus secreciones. En algunos casos los machos solos, en otros los machos y las hembras, diferían así levemente, según observo. Cuando las diferencias son algo más marcadas y cuando ambos sexos y todas las edades son afectados por ellas, todos los entomólogos clasifican las formas respectiva como especies. Pero ningún observador puede determinar para beneficio de otro, aunque pueda hacerlo par sí mismo, cual de esas formas fitófagas debería ser llamada especie y cual variedad. Mr. Walsh clasifica las formas que puede suponerse que se cruzarían libremente, como variedades, y las que parecen haber perdido ese poder, como especies. Pero las diferencias dependen de que los insectos se han alimentado duramente largo tiempo de plantas distintas, y por eso no pueden esperarse que los eslabones intermedios que relacionan las diversas formas hayan de encontrarse ahora. Así el naturalista pierde su mejor guía para determinar si debe clasificar las formas dudosas como variedades o como especies. Necesariamente ocurre lo mismo con
organismos estrechamente aliados que habitan continentes o islas distintos. Por otra parte, cuando un animal o planta existe en todo el mismo continente o en muchas islas del mismo archipiélago y presentan formas diferentes en las diversas regiones, siempre hay buenas probabilidades de que se descubrirán formas intermedias que vincularan los estados extremos y entonces éstas son rebajadas al grado de variedades.
Unos pocos naturalistas sostienen que los animales nunca presentan variedades; pero esos mismos naturalistas clasifican después la más leve diferencia como de valor específico y cuando se encuentra la misma forma idéntica en dos países distintos o en dos formaciones geológicas diferentes, creen que hay dos especies distintas ocultas bajo el mismo ropaje. Es decir que el mismo término de especie viene a ser empleado como mera abstracción inútil, implicando y dando por supuesto un acto separado de la creación. Es seguro que muchas formas, consideradas por jueces muy competentes como variedades, se parecen tan completamente a las especies en sus caracteres, que han sido clasificadas como tales por otros jueces muy competentes. Pero discutir si deben o no ser llamadas especies o variedades antes de que se haya aceptado generalmente una definición de esos términos es espectacular en el aire.
Muchos de los casos de variedades fuertemente marcadas o especies dudosas merecen consideración, pues en la tentativa para determinar su categoría se han expuesto varias líneas interesantes de argumentación relativas a la distribución geográficas, a la distribución geográfica, a la variación analógica, al hibridismo, etc. Pero el espacio de que dispongo no me permite estudiarlas. En muchos casos no cabe duda de que una investigación atenta hará que los naturalistas se pongan de acuerdo sobre como clasificar formas dudosas. Sin embargo, hay que confesar que es en los países más conocidos donde encontramos mayor número de ellas. Me ha llamado la atención el hecho de que si un animal o planta en estado de naturaleza es muy útil al hombre o llama su atención por alguna causa, inmediatamente se registraran variedades del mismo. Esas variedades, además, serán clasificadas a menudo por algunos autores como especies. Obsérvese el roble común: ¡ cuantamente ha sido estudiado! Sin embargo, un autor alemán descubre más de una docena de especies entre las formas casi universalmente los otros botánicos consideraban variedades; y en este país se puede citar a las más altas

57
autoridades botánicas y hombres prácticos para demostrar que los robles sesil y pedunculado son verdaderas y definidas especies o meras variedades.
Puedo aludir aquí a una notable memoria publicada últimamente por A. De Candolle sobre los robles de todo el mundo. Nadie ha tenido materiales más amplios para discernir las especies ni los ha trabajado con más celo y capacidad.
Primero expone en detalle todos los númerosos puntos de la estructura que varían en las diferentes especies y calcula numéricamente la frecuencia relativa de la variación. Especifica más de una docena de caracteres que pueden encontrarse variando hasta en la misma rama, a veces según la edad o el desarrollo, a veces sin alguna razón asignable. Tales caracteres, por supuesto, no tienen valor de especie, pero son como ha observado Asa Gray comentando este estudio, de los que entran generalmente en las definiciones de especie. De Candolle dice después que da la categoría de especie a las formas que difieren en caracteres que nunca varían en el mismo árbol y que nunca se han encontrado relacionados por estados intermedios. Después de este pasaje, resultado de tanto trabajo, recalca “ se engañan los que repiten que la mayoría de nuestras especies están claramente delimitada y que las especies dudosas son la exigua minoría. Esto parecía cierto mientras se conocía imperfectamente el género, y sus especies se fundaban en pocos ejemplares, es decir, que eran provisionales. En cuanto empezamos a conocerlas mejor aparecen formas intermedias y aumentan las dudas sobre los limites en las especies”. Añade que las especies más conocidas son las que presentan mayor número de variedades y subvariedades espontaneas. Así la quercus robur tiene 28 variedades, todas las cuales, exceptuando seis, están agrupadas en torno a tres subespecies, a saber: la quercus pedunculata, la sesiliflora y la bubescens. Las formas que vinculan entre si a esas tres subespecies son relativamente raras; y, como observa también Asa Gray, si esas vinculadoras raras ahora se extinguieran totalmente, las tres subespecies conservarían entre si exactamente la misma relación que las cuatro o cinco especies provisionalmente admitidas, que rodean tan estrechamente a la típica quercus robur. Finalmente, de Candolle admite que las 300 especies que serán enumeradas en su Prodromo como pertenecientes a la familia del roble, por lo menos dos tercios son especies provisionales,
es decir, que no se saben que cumplan estrictamente la definición, dada antes, de una especie verdadera. Debe añadirse que de Candolle ya no cree que las especies sean creaciones inmutables, son concluye que la teoría derivada es la más natural y “ la más acorde con los hechos conocidos de la paleontología, la botánica y la zoología geográficas, y de la estructura y clasificación anatómicas”.
Cuando un joven naturalista empieza a estudiar un grupo de organismos completamente desconocidos para él, se ve muy perplejo al principio para determinar que diferencias ha de considerar en la categoría de especies y cuales en la de variedad, porque no sabia nada del grado de variación a que esta sometido ese grupo, al menos, con cuanta frecuencia hay variaciones. Pero si limita su atención a una sola clase en un solo país, pronto se decidirá en cuanto a como clasificar la mayoría de las formas dudosas. Su tendencia general será enumerar muchas especies, porque quedara impresionado, como los dos criadores de palomas o gallinitas a que aludimos, ante el alcance de las diferencias en las formas que continuamente esta estudiando, y tiene poco conocimiento general de la variaron analógica en otros grupos y otros países con lo cual corregiría sus primeras impresiones. A medida que amplia el campo de sus observaciones se encontrará cada vez con más casos de dificultad, porque encontrará mayor número de formas estrechamente relacionadas entre si. Pero si sus observaciones son ampliamente extendidas, el final podrá probablemente decidirse, aunque lo conseguirá con tal de admitir mucha variación, y lo apropiado de esta admisión será discutido por otros naturalistas. Cuando llegue a estudiar formas vinculadas traídas de países que no son continuos, caso en el cual no puede abrigar la esperanza de encontrar eslabones intermedios, se vera obligado a confiar casi exclusivamente en la analogía, y entonces sus dificultades habrán llegado al máximo.
Ciertamente, no se ha trazado aun una línea clara de demarcación entre las especies y las subespecies, es decir, las formas que en opinión e algunos naturalistas se acercan mucho pero no llegan a la categoría de especies; ni tampoco entre las subespecies y las variedades menores y diferencias individuales.
Esas diferencias se mezclan unas con otras mediante una serie insensible, y una serie da al espíritu la impresión de un pasaje verdadero.

58
Por eso, aunque las diferencias individuales son de escaso interés para el sistematizador, yo las miro como de la mayor importancia para nosotros, pues constituyen el primer paso hacia esas variedades leves que apenas son dignas de recordarse en las obras de la historia natural. Y miro las variedades que en algún grado son más distintas y permanentes, como pasos hacia variedades más permanentes y marcadas; y estas ultimas, como a las que conducen a las subespecies, y de ahí a las especies. El paso de una etapa de diferencia a otra puede ser en muchos casos simple resultado de la naturaleza del organismo y de las diferentes condiciones físicas a que ha estado expuesto por mucho tiempo; pero con respecto a los caracteres más importantes y adaptativos, este paso de una etapa de diferencia a otra puede ser atribuido sin temor a la acción acumulada de la selección natural, que explicaremos más adelante, y a los efectos del aumento del uso y desuso de las partes. Una variedad bien marcada, por consecuencia, puede ser llamada especie incipiente; pero para juzgar si esta opinión es justificada o no, habrá que considerar el peso de los diversos hechos y reflexiones que expondremos en el curso de esta obra.
No hay que suponer que todas las variedades o especies incipientes alcanzaran la categoría de especies. Pueden extinguirse o pueden resistir como variedades por periodos muy largos, según ha demostrado Mr. Wollaston con las variedades de ciertos moluscos fósiles terrestres de Madera y Gaston de Saporta con ciertas plantas. Si una variedad prospera hasta superar en número a la especie de la madre, tendrá entonces la categoría de especie y aquella pasara a la variedad; o podría llegar a suplantar y exterminar a la especie madre; o ambas podrían coexistir y ser consideradas como especies independientes. Pero sobre esto hemos de volver más adelante.
Por todas estas observaciones, considero el termino especie como arbitrariamente dado, por comodidad, a un conjunto de ejemplares que se parecen estrechamente; y creo que no defiere en lo esencial del termino variedad, que se de a formas menos distintas y más fluctuantes. Además, la expresión variedad, en comparación con las diferencias meramente individuales, es aplicada también arbitrariamente, por razones de comodidad.
LAS ESPECIES DE GRAN DISPERSIÓN, LAS MUY DIFUNDIDAS Y LAS más COMUNES, SON LAS QUE VARIAN MAS.
Guiado por consideraciones teóricas pensé que se podrían obtener algunos resultados interesantes con respecto a la naturaleza de las relaciones entre las especies que más varían tabulando todas las variedades de diversas floras bien elaboradas. Al principio esto parecía tarea fácil; pero Mr. H. C. Watson, a quien debo mucho por sus valioso consejos y ayuda en este asunto, no tardo en convencerme de que había muchas dificultades, como lo hizo después constar el Dr. Hooker, en términos aun más enérgicos. Reservare para una obra futura la discusión de estas dificultades y tablas de los números proporcionales de las especies que varían. El Dr. Hooker me permito añadir que después de haber leído cuidadosamente mi manuscrito, y examinado las tablas, piensa que las afirmaciones siguientes están bien establecidas. Todo el tema, empero, tratando como lo es necesariamente aquí en forma muy breve, resulta más bien desconcertante y no pueden evitarse alusiones a la lucha por la existencia, divergencias de caracteres y otras cuestiones que habrán de ser estudiadas más adelante.
Alfonso de Candolle y otros han demostrado que las plantas que abarcan amplias regiones, generalmente presentan variedades; y esto era de esperar, puesto que están sometidas a diversas condiciones físicas y entran en competencia (lo cual, como veremos más adelante, es circunstancia igual o más importante) con diferentes conjuntos de seres orgánicos. Pero mis tablas demuestran además que en un país limitado las especies que son más comunes, es decir, que más abundan en ejemplares, y las especies que están difundidas más ampliamente en su propio país (hecho diferente del que abarquen muchas regiones, y hasta cierto punto también del que sean o no comunes) muy a menudo dan origen a variedades suficientemente marcadas como para haber sido registradas las obras botánicas. De ahí que las especies más prosperadas o, como podrían llamarse. Dominantes, -las que abarcan muchas regiones, están muy difundidas en su propio país y son las más númerosas en ejemplares- son las que producen más a menudo variedades bien marcadas o, como las considero yo, especies incipientes. Y esto, quizá podría preverse: porque como las variedades, para convertirse de algún modo en permanentes, tienen que luchar con los otros habitantes del país, las

59
especies que ya son dominantes serán las que más probablemente den descendientes que, si bien modificados en grado todavía leve, heredan esas ventajas que permitieron a sus antecesores predominar sobre sus compatriotas.
En estas observaciones sobre predomino debe entenderse que se alude solamente a las formas que entran en competencia unas con otras, y más especialmente a los miembros del mismo genero o clase, que tienen hábitos parecidos de vida. Con respecto al número de individuos o a lo común de la especie, la comparación, por supuesto, se refiere solamente a los miembros del mismo grupo. Una de las plantas más elevadas puede ser considerada predominante si tiene ejemplares más númerosos y más difundidos que las otras plantas del mismo país que viven bajo las mismas condiciones. Una planta de esta clase no es menos predominante por el hecho de que algunas hierbas filamentosas del agua o algunos hongos parásitos sean infinitamente más númerosos como ejemplares y están difundidos con más amplitud. Pero si esas hierbas filamentosas u hongos parásitos superan a sus semejantes en los sentidos mencionados, entonces serán los que predominan dentro de su clase.
ESPECIES DE LOS GENEROS MAYORES EN CADA PAIS VARIAN CON más FRECUENCIA QUE LAS ESPECES DE LOS GENEROS MENORES.
Si las plantas que habitan un país, tal como las describe cualquier flora, son divididas en dos masas iguales, poniendo e un lado a todas las de los géneros mayores, (es decir los géneros que comprenden muchas especies y del otro las de los géneros menores, se vera que los primeros incluyen un número algo mayor de las especies muy comunes o muy difundidas o dominantes.
Esto podría preverse, porque el mero hecho de que muchas especies del mismo genero habiten un país demuestra que en algunas condiciones orgánicas o inorgánicas de ese país hay algo favorable para el genero; en consecuencia, podría esperarse encontrar en los géneros mayores, o sea lo que incluyen muchas especies, un número proporcionalmente mayor de especies dominantes.
Pero son tantas las causas que tienden a obscurecer este resultado, que todavía me sorprende que mis tablas den una pequeña mayoría del lado de los géneros mayores.
Aludiré aquí solamente a dos causas de obscuridad. Las plantas que buscan aguas dulces y sales generalmente crecen en regiones muy extensas y están muy difundidas, pero esto parece relacionarse con la naturaleza de las zonas habitadas por ellas y tienen poca ninguna relación con el volumen de los géneros a que pertenecen las especies. Además, plantas que ocupan lugares inferiores en la escala de la organización están mucho más difundidas que las que ocupan puestos superiores; y aquí tampoco existe relación estrecha con el volumen de los géneros. La causa de que las plantas inferiores aparezcan en regiones tan extensas será tratada en nuestro capitulo sobre la distribución geográfica.
Por mirar las especies solamente como variedades muy marcadas y bien definidas, pude prever que las especies de los géneros mayores en cada país presentarían a menudo más variedades que las de los géneros menores, pues dondequiera que se han formado muchas especies estrechamente relacionadas (es decir especies del mismo genero) muchas variedades o especies incipientes deberán estar formándose ahora, por regla general. Donde crecen muchos árboles grandes, esperamos encontrar retoños. Donde se han formado muchas especies de un genero por variación, las circunstancias han sido favorables para la variación, y por so esperamos que sigan siendo favorables para ello. En cambio, si miramos cada especie como un acto especial de creación, n hay razones visibles para que se produzcan más variedades en un grupo que contiene muchas especies que en uno que contiene pocas. Para probar la verdad de este pronostico he distribuido las plantas de doce países y los insectos coleópteros de dos distritos en dos masas casi iguales, poniendo en una las especies de los géneros mayores y en la otra las de los géneros menores; e invariablemente la proporción de los géneros mayores que presentaba variedades era mayor que la de las especies de los géneros menores. Además, las especies de los géneros mayores que presentan algunas variedades siempre ofrecen mayor número medio de variedades que las especies de los géneros reducidos. Ambos resultados se producen igualmente cuando todos los géneros menores, que tienen solamente de una a cuatro especies, excluidos totalmente de las tablas. Estos hechos cobran un significado claro si se supone que las especies no son más que variedades fuertemente marcadas y permanentes, porque dondequiera que se han formado muchas especies de los mismos

60
géneros, o dondequiera – si podemos decirlo así- que la manufactura de las especies ha estado activa, deberíamos encontrar generalmente la manufactura todavía en acción, sobre todo porque tenemos todas las razones para creer que el proceso de manufacturar nuevas especies es lento. Y esto ciertamente se comprueba también si se considera a las variedades como especies incipientes; pues mis tablas demuestran claramente como regla general que dondequiera que se han formado muchas especies de un genero, las especies de ese genero presentan cierto número de variedades, es decir de especies incipientes, superior al número medio. No es que todos los géneros amplios estén variando mucho ahora y por consiguiente aumentando en el número de sus especies, ni que dejen de variar y aumentar los géneros pequeños; Pues si así ocurriera, esto seria fatal para mi teoría, ya que la geología nos demuestra claramente que géneros pequeños, en el curso del tiempo, aumentan mucho de volumen, a menudo; y que géneros grandes con frecuencia llevan a su culminación, decadencia y desaparición. Todo lo que deseamos poner de manifiesto es que, cuando se han formado muchas especies de un género, por termino medio se están formando muchas aun, y esto efectivamente se comprueba.
Muchas de las especies incluidas en los géneros mayores se parecen a las variedades en que están relacionadas muy estrechamente, pero en forma desigual, unas con otras, y también en que abarcan regiones limitadas.
Hay otras relaciones entre las especies de los géneros grandes y sus variedades registradas que merecen ser señaladas. Hemos visto como no existe criterio infalible para distinguir especies de variedades bien marcadas, y cuando no se han encontrado los eslabones intermedios entre las formas dudosas, los naturalistas se ven obligados a decidirse por el grado d diferencia entre ellas, juzgando por analogía si ese grado de diferencia basta o no para elevar a una de ella o las dos categoría de especies. Por consiguiente, el grado de diferencia es un criterio muy importante para establecer si dos formas deben ser clasificadas como especies o como variedades. Ahora bien; Fries, en relación con las plantas, y Westwood, en relación con los insectos, han observado que en los géneros grandes el grado de diferencia entre las especie es a menudo excesivamente exiguo. He procurado comprobar esto numéricamente por términos medio, y hasta donde llegan, mis resultados
imperfectos confirman la opinión. También he consultado a varios sagaces y experimentados investigadores, y después de pensarlo han coincidido en este punto de vista. En tal sentido, por consiguiente, las especies de los géneros mayores se parecen a las variedades más que las especies de los géneros menores. O puede formularse el principio en otra forma, diciendo que en los géneros mayores, en los cuales se están manufacturando ahora un número de variedades o especies incipientes mayor del termino medio, muchas de las especies ya manufacturadas también hasta cierto punto se parecen a variedades porque difieren entre sí por menos cosas de las que constituyen el grado habitual de diferencia.
Además, las especies de los géneros mayores se relacionan entre sí del mismo modo en que las variedades de cualquier especie se relacionan unas con otras. Ningún naturalista pretende que todas las especies de un genero sean igualmente distintas entre sí; generalmente pueden ser divididas en subgéneros o secciones o grupos menores.
Como bien ha observado Fries, pequeños grupos de especies están generalmente reunidos como satélite en torno a otras especies. Y ¿qué son las variedades sino grupos de formas desigualmente relacionadas entre sí y reunidas en torno a sus especies madres? Indudablemente, existe un punto muy importante de diferencia entre variedades y especies, a saber: que el grado de diferencia entre variedades, cuando se las compara entre sí o con sus especies madres, es mucho menor que el que existe entre las especies del mismo genero. Pero cuando lleguemos a discutir este principio, como yo lo llamo, de la divergencia de los caracteres, veremos como se puede explicar esto y como las diferencias menores entre las variedades tienden a transformarse en las diferencias mayores entre las especies.
Hay otro punto digno de atención. Generalmente las variedades abarcan zonas restringidas: esta afirmación apenas parece algo más que una perogrullada, porque si se advirtiera que una variedad abarca zonas mayores que la de su presunta madre, habría que invertir los términos. Pero hay razones para creer que las especies estrechamente vinculadas a otras especies que por ello parecen variedades, cubren a menudo comarcas muy restringidas. Por ejemplo, Mr. H. C. Watson me ha marcado en el bien ordenado Catalogo de Plantas de Londres (4ª edición), 63 plantas que ahí están clasificadas como especies, pero que él considera tan estrechamente vinculadas con otras especies

61
que resultan de valor dudoso: esas 63 llamadas especies abarcan por lo general un termino medio de 6,9 de las regiones en que Mr. Watson ha dividido a Gran Bretaña. Ahora bien, en ese mismo Catalogo, 53 variedades reconocidas están registradas, y estas abarcan 7,7 de las regiones, mientras que las especies a que pertenecen esas variedades abarcan más de las 14,3 regiones. De modo que las variedades reconocidas tienen casi el mismo campo restringido que las formas estrechamente vinculadas que Mr. Warson me ha marcado como especie dudosas, pero que los botánicos clasifican casi universalmente como especies verdaderas.
RESUMEN
En suma: las variedades no pueden ser distinguidas de las especies si no es, primero por el descubrimiento de formas vinculadoras intermedias, y, segundo, por cierto grado no definido se diferencia entre ellas; pues dos formas que difieran muy poco son generalmente clasificadas como variedades, a pesar de que no pueden ser estrechamente vinculadas una con otra; pero no se puede definir el grado de diferencia que se considera necesario para dar a dos formas cualesquiera la categoría de especie. En los géneros que comprenden un número de especies mayor que el término medio en cualquier país, las especies de esos géneros tienen un número de variedades mayor que el término medio. En los grandes géneros, la tendencia es que las especies estén estrechamente, aunque en forma desigual, vinculadas entre sí formando como pequeños racimos en torno a otras especies. Especies muy vinculadas con otras especies tienen el parecer zonas de prosperidad muy restringidas. En todos esos sentidos, las especies de los grandes géneros presentan fuerte analogía con las variedades. Y podemos comprender claramente esas analogías si las especies existieron alguna vez como variedades y se formaron así; Mientras que esas analogías son completamente inexplicables si las especies son creaciones independientes. También hemos visto que son las especies más prosperas o dominantes de los géneros mayores dentro de cada clase las que, por termino medio, producen el mayor número de variedades; y las variedades, como veremos después, tienden a convertiste en nueva y distintas especies. Así los géneros mayores tienden a aumentar, y en toda la naturaleza las formas de vida que son ahora predominantes tienden a volverse más predominantes aun, dejando muchos descendientes modificados y dominantes. Pero
por pasos que se explicaran más adelante, los géneros mayores tienden a dividirse en géneros menores, y así las formas de vida en todo el universo resultan divididas en grupos subordinados a otros grupos.
CAPITULO III
LA LUCHA POR LA EXISTENCIA
SU RELACION CN LA SELECCIÓN NATURAL.
Antes de entrar en el tema de este capitulo debo hacer algunas observaciones preliminares para demostrar como influye la lucha por la existencia sobre la selección natural. Se ha visto en el último capitulo lo que entre los seres orgánicos en estado de naturaleza hay alguna variabilidad individual: a decir verdad, no tengo conciencia de que ello haya sido impugnado nunca.
Para nosotros no tiene importancia que una multitud de formas dudosas sean llamadas especies o subespecies o variedades, por ejemplo, que categoría han de ocupar las doscientas o trescientas formas dudosas de plantas británicas, siempre que se reconozca la existencia de algunas variedades bien marcadas. Pero la mera existencia de la variabilidad individual y de unas cuantas variedades bien marcadas, aunque es necesario como fundamento de la obra, nos ayuda poco a comprender como surgen las especies de la naturaleza. ¿Cómo se han formado todas esas exquisitas adaptaciones de una parte de la organización a otra parte, y a las condiciones de vida, y de un ser orgánico a otro? Vemos esas notables coadaptaciones clarísimas en picamaderos y en el muérdago, y solamente con un poco menos de claridad en el más humilde parásito que se aferra a los pelos de un cuadrúpedo o las plumas de una ave; las vemos en la estructura del escarabajo que se zambulle en el agua, en la semilla emplumada que se traslada a impulsos de la brisa más ligera; en suma, vemos notables adaptaciones en todas partes y en todas las cosas del mundo orgánico.
También puede preguntarse como es que las variedades, que se ha llamado especies incipientes, se convierten al fin en especies verdaderas y diferenciadas, las cuales en la mayoría de los casos difieren evidentemente unas de las otras mucho más que las variedades de la misma especie. ¿Cómo surgen esos mismos grupos de especies que constituyen lo que se llama géneros distintos y que difieren entre sí más que las especies del mismo genero? Todos esos resultados, como veremos más detalladamente en el

62
capitulo próximo, se siguen de la lucha por la vida. Debido a esa lucha las variaciones, por leves que sean y provengan de donde provengan, han de tender, si son beneficiosas de algún modo para los individuos de la especies en sus relaciones infinitamente complejas con otros seres orgánicos y con sus condiciones físicas de vida, a la preservación de tales individuos, y generalmente serán heredadas por la descendencia. Los descendientes tendrán así mejores probabilidades de sobrevivir porque de los muchos ejemplares de cualquier especie que nacen periódicamente, solamente puede sobrevivir un número reducido. He dado a este principio, por el cual se preserva cualquier variación leve si es útil, el nombre de la selección natural, a fin de señalar su relación con el poder de la selección que tiene el hombre mismo. Pero la expresión utilizada a menudo por Mr. Herbert Spencer, supervivencia de los más aptos, es más precisa, y a veces igualmente adecuada. Hemos visto que el hombre, mediante la selección, puede producir ciertamente grandes resultados y puede adaptar seres orgánicos a sus propios fines gracias a la acumulación de variedades ligeras pero útiles que le ha presentado la mano de la naturaleza. Pero la selección natural, como veremos después, es un poder que esta siempre pronto para obrar y tan inconmensurablemente superior a los débiles esfuerzos del hombre como lo son las obras de la naturaleza con respecto a las del arte.
Trataremos ahora con un poco más de detalle la lucha por la existencia. En mi obra futura este tema será estudiado, como lo merece, con más extensión. El mayor de los Candolle y Lyell han demostrado amplia y filosóficamente que todos los seres orgánicos están expuestos a intensa competencia. Con respecto a las plantas, nadie ha tratado este tema con más ingenio y capacidad que W. Herbert, deán de Manchester, gracias, evidentemente, a sus grandes conocimientos de horticultor. Nada más fácil que admitir de palabra de la verdad de la lucha universal por la vida; Nada más difícil –al menos así lo he encontrado yo- que tener continuamente en cuenta esta conclusión. Pero a menos que ella este bien asimilada por nuestra mente, toda la economía de la naturaleza, con cada uno de sus hechos sobre distribución, rareza, abundancia, extinción, y variación, será vista confusamente o del todo tergiversada. Vemos la faz de la naturaleza radiante de alegría; vemos a menudo superabundancia de alimento, pero no vemos, o no recordamos que las aves que cantan descuidadamente en
torno a nosotros viven casi todas de insectos o semillas y destruyen así constantemente la vida; no recordamos en que cantidad son destruidos estos cantores, o sus huevos, o sus crías, por aves y bestias de rapiña; Y no siempre tenemos en cuenta que aunque el alimento puede sobrar ahora, no ocurre así en todas las estaciones de cada año.
ÉL TERMINO “LUCHA POR LA VIDA” SE EMPLEA EN SENTIDO AMPLIO.
Uso esta expresión en un sentido amplio y metafórico, que incluye la dependencia de un ser respecto de otro y –lo que es más importante- incluye no solo la vida del individuo, sino también el éxito al dejar descendencia. De dos cánidos, en tiempo de hambre, luchan entre sí por el cual conseguirá comer o vivir, pero de una planta en él limite de un desierto se dice que lucha por la vida contra la sequedad, aunque más propio seria decir que depende de la humedad. De una planta que produce anual mente un millar de las que, por termino medio, solo una llega a completo desarrollo, puede decirse, con más exactitud, que lucha por las plantas de la misma clase o de otras que ya cubrían el suelo. El muérdago depende del manzano y de algunos otros árboles; más solo en un sentido muy amplio puede afirmarse que lucha con estos árboles, pues si sobre uno mismo de estos crecen demasiados parásitos, el árbol se extenúa y muere; pero de varias plantitas de muérdago que crecen muy juntas sobre la misma rama puede decirse con más exactitud que luchan mutuamente. Como el muérdago es diseminado por los pájaros, su existencia depende de ellos, y cabe decir metafóricamente que lucha con otras plantas frutales, tentando a los pájaros a tragar y diseminar de este modo sus semillas. En estos varios sentidos, que pasan insensiblemente de uno a otro, empleo por razón de conveniencia la expresión general lucha por la existencia.
PROGRESIÓN GEOMETRICA DEL AUMENTO
Una lucha por la existencia se desprende inevitablemente de la alta proporción en que tienden a aumentar todos los seres orgánicos. Cada ser que durante su tiempo natural de vida produce varios huevos o semillas debe estar sometido a destrucción en algún periodo de su vida y en alguna estación o año ocasional porque de otro modo, por l principio de su aumento en progresión geométrica, su número se volvería enseguida tan desmesuradamente grande que ningún país

63
podría sostenerlo. De ahí que como son producidos más ejemplares de los que tienen posibilidades de sobrevivir, tiene que haber en cada caso una lucha por la existencia, ya sea de un individuo con otro de la misma especie, o con individuos de especies distintas, ya sea con las condiciones de vida. Es la doctrina de Malthus aplicada con fuerza multiplicada a todo el reino animal y a todo el reino vegetal, porque en este caso no puede haber aumento artificial de nutrición ni restricciones al casamiento dictadas por la prudencia. Aunque algunas especies pueden estar aumentando en número ahora, más o menos rápidamente, no puede ocurrir eso con todas porque el mundo no podría sostenerlas.
No hay excepción para la regla de que todo ser orgánico aumenta naturalmente en tal proporción que si no fuera destruido la tierra no tardaría en quedar cubierta por la descendencia de una sola pareja. Hasta el hombre, que es lento para reproducirse, ha duplicado su número en 25 años, y a ese paso en menos de mil años no habría, literalmente hablando, espacio para su progenie. Linneo ha calculado que si una planta anual produjese solo dos semillas – y no hay vegetal que produzca tan poco- y los retoños consiguientes produjeran otras dos cada uno al otro año, y así sucesivamente, al cabo de veinte años habría un millón de esas plantas. Se calcula que el elefante es el animal más lento para reproducirse entre todos los conocidos, y yo me he tomado algún trabajo para calcular su probable proporción mínima de aumento natural; se puede suponer con toda tranquilidad que empieza a procrear a los treinta años y sigue haciéndolo hasta los noventa, produciendo seis hijos en el intervalo y sobreviviendo aun hasta los cien años: en tal caso, después de un periodo d 740 ó 750 años habría cerca de 19 millones de elefantes vivos, descendientes de la primera pareja.
Pero tenemos mejores pruebas sobre este tema que los simples cálculos teóricos, a saber, los númerosos casos registrados de incremento asombrosamente rápido de varios animales en estado de naturaleza, cuando las circunstancias le han sido favorables durante dos o tres estaciones seguidas más notable es la prueba tomada de nuestros animales domésticos de muchas clases que se han vuelto salvajes en varias partes del mundo: si las afirmaciones sobre la progresión del aumento de los vacunos y caballos – animales de procreación lenta- en la América del Sur, y más recientemente en Australia, no hubieran sido bien verificadas, parecerían increíbles. Lo mismo ocurre con los vegetales; se podrían
presentar casos de plantas importadas que se han vuelto comunes en islas enteras al cabo de diez años apenas. Varias de las plantas, como el cardo silvestre y un cardo alto que ahora son de las más comunes en todas las llanuras del plata cubriendo leguas enteras de superficie con exclusión casi total de todo otro vegetal, han sido importadas de Europa; y hay plantas que ahora se extienden por la India, según me informa el Dr. Falconer, desde el cabo Comorin hasta el Himalaya y que han sido importadas de América desde que esta fue descubierta. En tales casos e infinitos más que podrían darse, nadie supone que la fecundidad de esos animales y plantas han sido súbitamente y en forma temporal aumentada en grado apreciable. La explicación obvia es que las condiciones de vida han sido muy favorables y por consecuencia ha habitado menos destrucción de ejemplares y descendientes han podido reproducirse. Simple progresión geométrica de aumento, cuyo resultado nunca deja de ser asombroso, explica su incremento extraordinariamente rápido y la más amplia difusión obtenida en su nueva tierra.
En estado de naturaleza casi toda planta adulta produce anualmente semilla y entre los animales hay muy pocos que no se apareen todos los años. De ahí podemos afirmar confiadamente que todas las plantas y animales tienden a aumentar en progresión geométrica, que todos se acumularían rápidamente en cualquier región en que pueden existir de algún modo, y que esta tendencia geométrica a aumenta debe ser contenida mediante la destrucción en algún periodo de la vida. Estamos tan familiarizados con los animales domésticos más grandes que nos exponemos a equivocarnos: no vemos que sean destruidos en gran número, pero no recordamos que millares de ellos son sacrificados anualmente para nuestra comida y que en estado de naturaleza un número igual tendría que ser liquidado de algún modo.
La única diferencia entre organismos que anualmente producen huevos y semillas por millares y los que producen extremadamente pocos es que los de reproducción lenta requerirían algunos años más para poblar, bajo condiciones favorables, una región entera, por grande que sea. El cóndor pone un par de huevos y el avestruz una veintena, y sin embargo, en el mismo país el cóndor puede ser el más númeroso de los dos; el petrel de Fulmar pone un solo huevo, pero se cree que es el ave más númerosa del mundo. Una mosca deposita cientos de huevos, y otra, la hippoboska, pone solamente uno; pero esta

64
diferencia no determina cuantos ejemplares de las dos especies pueden ser sostenidos por una región. La gran cantidad de huevos tiene alguna importancia para esas especies que dependen de una provisión variable de comida, porque les permite aumentar rápidamente. Pero la verdadera importancia de la gran cantidad de huevos o semillas es compensar la destrucción en algún periodo de la vida; y este periodo, en la mayoría de los casos, sobreviene tempranamente. Si un animal puede proteger de algún modo sus huevos o sus hijos podrá producirlos en número reducido y sin embargo se mantendrá el término medio de sus ejemplares; Pero si son destruidos muchos huevos o hijos, habrá que producir muchos porque si no la especie se extinguirá. Para mantener el número completo de ejemplares de cierto árbol que viviera por termino medio mil años, bastaría que produjese una sola semilla en mil años, siempre que esa semilla no fuese destruida y se le pudiese asegurar la germinación en lugar adecuado. De modo que en todos los casos el número medio de cualquier animal o planta depende solo indirectamente del número de huevos y semillas.
Al mirar la naturaleza es indispensable tener en cuenta las conclusiones mencionadas y no olvidar nunca que de cada ser orgánico se puede decir que esta esforzándose hasta el máximo por aumentar; que cada uno vive mediante una lucha en algún periodo de su vida; que una abundante destrucción recae inevitablemente sobre los nuevos y los viejos, durante cada generación o con intervalos que se repiten. Alíviense cualquier obstáculo, mitíguese la destrucción por poco que sea, y casi instantáneamente aumentara el número de la especie hasta un nivel incalculable.
NATURALEZA DE LOS OBSTACULOS AL AUMENTO.
Las causas que traban la tendencia natural de cada especie a aumentar son muy obscuras. Observemos las especies más vigorosas: cuanto más númerosa sea, tanto más tendera a aumentar sin límite. No sabemos exactamente cuales son los obstáculos que se presentan ni siquiera en un solo caso. Y esto no sorprenderá a nadie que reflexione en nuestra ignorancia sobre este asunto, aun con respecto al género humano, incomparablemente mejor conocido que cualquier otro. Esta cuestión de los obstáculos al aumento ha sido tratada con autoridad por varios autores, y abrigo la esperanza de estudiarla extensamente en una obra futura,
especialmente con respecto a los animales bravíos de la América del Sur. Aquí solamente haré algunas observaciones, nada para llamar la atención del lector sobre algunos puntos principales. Huevos o animales muy nuevos parecen los que sufren más por lo general, pero no siempre ocurre así. En los vegetales hay una gran destrucción de semillas, pero por algunas observaciones que he hecho parece que los retoños sufren más por germinar en terreno ya espesamente poblado de otras plantas.
Los retoños, además, son destruidos en gran número por diversos enemigos, por ejemplo, en un pedazo de tres pies de largo y dos de ancho, excavada y despejada, donde no podía haber sofocación de otras plantas, marque los retoños de nuestras hierbas nativas a medida que surgían, y de 397 no menos de 295 fueron destruidos, principalmente por babosas o insectos. Si se deja crecer un césped que siempre ha sido segado – y el caso seria el mismo con el pasto comido por el ganado-, las plantas más vigorosas matan siempre a las menos vigorosas, aunque adultas; y así de veinte especies que crecían en un trozo de hierba segadas (tres pies por cuatro) nueve especies perecieron porque se permitió a las otras crecer libremente.
La cantidad de alimento para cada especie establece, por supuesto, el límite extremo hasta el cual puede aumentar; Pero muy a menudo no es la dificultad de conseguir alimento, sino el hecho de servir de presa a otros animales lo que determina el número medio de una especie. Así, parece haber pocas dudas de que la cantidad de perdices, gallinetas y liebres que hay en todo el campo relativamente grande depende principalmente de la destrucción que se haga de las alimañas. Si no se matara una sola cabeza de caza durante los próximos veinte años, en Inglaterra, y si al mismo tiempo no se destruyeran las sabandijas, probablemente habría menos caza que ahora, aunque cientos de miles de animales son cazados actualmente cada año.
Por otra parte, en algunos casos, como el del elefante, ninguno es destruidos por animales de presa, pues aun el tigre de la India se atreve muy pocas veces a atacar a un elefantito protegido por su madre.
El clima desempeña función importante para determinar el número medio de una especie, y temporadas periódicas de frío o sequía extremas parecen ser los obstáculos más eficaces. He calculado (principalmente por la gran disminución del número de nidos en la

65
primavera) que el invierno de 1854-55 destruyo cuatro quintos de los pájaros de mis propias tierras; y es una destrucción formidable si recordamos que en las epidemias del hombre el diez por ciento constituye una mortalidad extraordinaria. A primera vista, la acción del clima parece ser completamente independiente de la lucha por la existencia, pero en cuanto al clima obra reduciendo el alimento, provoca la lucha más intensa entre los individuos, sean de la misma especie o de especies distintas, que viven de la misma clase de comida. Aun en los casos que el clima, por ejemplo mediante el frío extremo, obra directamente, serán los ejemplares menos vigorosos o los que han tenido menos alimentos durante el invierno que avanzaba los que más sufrirán. Cuando viajamos de Sur a norte o de una región húmeda a una seca, invariablemente vemos que algunas especies se vuelven cada vez más raras y finalmente desaparecen; y si el cambio de clima es pronunciado, nos sentimos tentados a atribuir todo el efecto a su actuación directa. Pero esta opinión es errónea; olvidamos que cada especie, aun donde abunda, sufre constantemente enormes destrucciones en algún periodo de su vida, provenientes de sus enemigos o de competidores para el mismo lugar y alimento; y si esos enemigos o competidores fuesen favorecidos en el menor grado por algún leve cambio de clima, aumentaran en número, y como cada zona ya esta repleta de moradores las otras especies tendrán que disminuir. Cuando viajamos hacia el sur y vemos que una especie disminuye, podemos estar seguros de que la causa reside tanto en el hecho de que otras especies son favorecidas como en el de que esta es perjudicada. Así pasa cuando viajamos hacia el Norte, aunque en grado algo menor, porque el número de especies de todas clases, y por lo tanto también de competidores, disminuye al avanzar hacia el Norte o al subir una montaña, tropezamos con ejemplares deformes, debido a la acción directamente dañosa del clima, con más frecuencia que cuando vamos hacia el Sur o descendemos una montaña. Cuando llegamos a las regiones árticas o a cimas cubiertas de nieve o desiertos absolutos, la lucha por la existencia se produce casi exclusivamente contra los elementos.
Que el clima obra principalmente en forma indirecta favoreciendo a otras especies es algo que vemos claramente en el número prodigioso de plantas que en nuestros jardines pueden soportar perfectamente nuestro clima, pero que nunca se vuelven naturales del todo
porque no pueden competir con nuestras plantas ni residir la destrucción de nuestros animales nativos.
Cuando una especie, debido a que circunstancias muy favorables, aumenta desmesuradamente en una región pequeña, las epidemias, por lo menos esto parece ocurrir generalmente con nuestros animales de caza- se producen a menudo y con eso tenemos un obstáculo independiente de la lucha por la vida. Pero algunas de estas llamadas epidemias parecen debidos a algunos parásitos que por alguna causa, probablemente debido en parte a la facilidad de propagarse entre los animales apiñados, han sido extraordinariamente favorecidos: y aquí se presenta una especie de lucha entre el parásito y su víctima.
Por otra parte, en muchos casos, gran cantidad de ejemplares de la misma especie, con relación al número de sus enemigos, es absolutamente necesaria para su preservación. De ese modo podemos cultivar abundancia de maíz y nabos en nuestros campos, porque las semillas son excesivas con relación al número de aves que se alimentan de ellas; y no pueden las aves, aunque tienen superabundancia de alimento en esa estación, aumentar su número en forma que guarde proporción con la provisión de semilla porque su multiplicación es contenida durante el invierno; pero quienquiera que lo haya ensayado sabe cual difícil es sacar simiente de pocas plantas de trigo u otras semejantes en un jardín: cuando lo he procurado he perdido siempre hasta la ultima semilla. Esta teoría de que es necesaria una gran cantidad de ejemplares de la especie para su preservación explica, me parece, ciertos hechos extraños de la naturaleza, como el de que algunas plantas muy raras sean extremadamente abundantes en los pocos sitios en los que existen, y el de que algunas plantas colectivas, es decir abundantes en individuos, hasta en el último limite de la región en que crecen. En tales casos creeríamos que una planta solamente puede existir donde las condiciones de vida sean favorables que permitan vivir a muchos ejemplares y salvar así a la especie de la destrucción completa.
Debo añadir los buenos efectos del cruzamiento variado y los malos efectos del cruzamiento entre próximos, la menor duda en muchos de los casos: pero no he de extenderme aquí sobre este tema.

66
COMPLEJAS RELACIONES DE LOS ANIMALES Y PLANTAS EN LA LUCHA POR LA EXISTENCIA.
Se registran muchos casos que muestran cuan complejos son los obstáculos entre seres orgánicos que tienen que luchar juntos en el mismo país. Daré solo un ejemplo, aunque, sencillo, me intereso, Staffordshire, en la propiedad de un pariente en que yo tenia antes medios de investigación, había un brezal grande extremadamente estéril que nunca había sido tocado por el hombre; cientos de acres del mismo carácter exactamente habían sido tocados veinticinco años antes, habían plantado en ellos abetos escoceses. El cambio de la vegetación nativa de las parte sembrada del brezal fue muy notable, se ve generalmente al pasar de un suelo a otro completamente diferente: no solo el número proporcional de brezos había cambiado totalmente sino que nueve, de estas plantas (sin contar hierbas, padañas) florecían en las poblaciones y no podían encontrar el brezal. El efecto cobre los insectos debe haber sido aun mayor, porque comúnmente se veían seis aves insectívoras en las plantadas que no se veían, otra, mientras esta era frecuentada por dos o tres aves insectívoras diferentes. Por esto vemos cuan poderoso ha sido el efecto de la introducción de un solo árbol, si haberse hecho nada mas, fuera del cerco para que no entrase el ganado. Pero la importancia de todo cercamiento pudo ser comprobada claramente por mí en Farnham, Surrey.
Allí hay brezales extensos, con unos cuantos grupos de viejos abetos escoceses sobre las distantes cumbres de las colinas: en los diez últimos años han sido cercados grandes espacios, y abetos reproducidos por si solos están brotando en multitud, y tan juntos unos de otros que no todos pueden vivir. Cuando comprobé que estos descendientes no habían sido sembrados ni plantados quede tan sorprendido ante su número que busque varios puntos de observación desde donde podía contemplar cientos de acres no cercados de brezal, y no encontré literalmente ni un solo abeto escocés, excepto los viejos grupos plantados. Pero mirando de cerca entre los tallos del brezal encontré multitud de retoños y árbolitos que habían sido perpetuamente roídos por el ganado. En una yarda cuadrada, sobre un punto situado a varios centenares de yardas de uno de los viejos grupos, conté treinta y un árbolillos, uno de ellos con veintiséis anillos de crecimiento que durante largos años había procurado alzar la cabeza por sobre los tallos del matorral sin conseguirlo.
No es de extrañar que tan pronto como fue cercada la tierra esta se cubriera espesamente de jóvenes abetos que crecían vigorosamente. Pero el brezal era tan estéril y tan extenso que nadie se habría imaginado nunca que el ganado lo registrara tan estrecha y eficazmente en busca de comida.
Vemos aquí que el ganado determina en absoluto la existencia del abeto escocés; pero en varias partes del mundo los insectos determinan la existencia del ganado. Quizá el Paraguay ofrezca el ejemplo más curioso de esto, porque ni los vacunos ni los caballos ni los perros se han vuelto salvajes aunque se encuentran copiosamente al Norte y al Sur en estado bravío; y Azara (naturalista español) y Rengger han demostrado que esto se debe a que en el Paraguay existe en gran número una mosca que pone sus huevos en el ombligo de esos animales, recién nacidos. El aumento de esas moscas, númerosas como son, debe de ser contenido habitualmente por algún medio, probablemente por otros insectos parásitos. En consecuencia, si en el Paraguay disminuyeran ciertos pájaros insectívoros, los insectos parásitos aumentarían probablemente; y esto disminuiría el número de las moscas, que visitan los ombligos, y entonces los vacunos y los caballos se volverían federales, lo cual modificaría mucho
(Como he observado en parte de Sudamérica) la vegetación, a su vez, ello efectuaría, como acabamos de ver en Staffrshire, a los pájaros insectívoros, y así sucesivamente en círculos cada vez mayores de complejidad. Y no es que la naturaleza las relaciones hayan de ser siempre tan sencillas. La batalla dentro de la batalla debe reproducirse continuamente con varia fortuna; y sin embargo, a la larga, las fuerzas están equilibradas con tanta perfección que la faz de la naturaleza permanece uniforme por largos periodos, aunque de seguro la menor insignificancia daría la victoria a un ser orgánico sobre otro. Sin embargo, esta profunda ignorancia y tan grande nuestra presunción, que nos maravillamos al enterarnos de la extinción de un ser orgánico, y como no vemos la causa invocamos cataclismos que hayan desolado el mundo o inventamos leyes sobre la duración de las formas de vida.
Me siento tentado a dar un ejemplos más de la forma en que plantas y animales remotos de la escala de la naturaleza están vinculados por una red de complejas relaciones. Tendré ocasión de mostrar más adelante que la exótica Lobelia fulgens (flor ornamental, de raíces medicinales que debe su nombre al botánico flamenco Lobel, planta silvestre de

67
cuba de hojas pectorales y antivenéreas) nunca es visitada en mi jardín por los insectos, y en consecuencia, debido a su estructura peculiar, nunca da semilla. Casi todas nuestras orquídeas necesitan indispensablemente la visita de los insectos para que remuevan sus masas de polen y las fecunden. Por experimentos que he realizado encuentro que los abejorros son casi indispensables para fecundar los pensamientos (viola tricolor), pues otras abejas no visitan estas flores. También he encontrado que las visitas de las abejas son necesarias para fecundar ciertas clases de trébol; por ejemplo veinte cabezas de trébol holandés (trifolium repens) rindieron 2290 semillas; pero otras veinte, protegidas de las abejas, no rindieron ninguna sola. Aun: cien cabezas de trébol rojo (trifolium pratense) produjeron 22700 semillas, pero el mismo número de cabezas protegidas no produjo ninguna. Solamente los abejorros visitan el trébol, pues otras abejas no pueden alcanzar el néctar. Se ha sugerido que las polillas podrían fecundar al trébol. Pero dudo que pudieran hacerlo en el caso del trébol rojo, pues su peso no seria suficiente para deprimir los pétalos. De ahí podemos inferir como muy probable que todo el género de los abejorros se extinguiera o volviese muy raro en Inglaterra, el pensamiento y el trébol rojo se harían rarísimos y desaparecerían del todo. El número de abejorros que hay en cada región depende en gran medida del número de ratones de campo que destruyen sus panales y nidos, y el coronel Newman, que ha estudiado largamente los hábitos de los abejorros, cree que “más de dos tercios de ellos son destruidos así en toda Inglaterra”. Ahora bien, el número de ratones depende de gran parte, como todo el mundo sabe, del número de gatos; Y el coronel Newman dice; “cerca de aldeas y ciudades pequeñas he encontrado que los nidos de abejorros son más númerosos que en otras partes, lo cual atribuyó a la cantidad de gatos que destruyen a los ratones”. Por lo tanto es completamente creíble que la presencia de algún animal felino de gran número determine en una región, primero por intermedio de los ratones y después las abejas ¡la frecuencia de ciertas flores en esa región!
En el caso de cualquier especie intervienen muchos obstáculos diferentes que obran en distintos periodos de vida y distintas estaciones del año; generalmente uno o varios obstáculos dados serán los más potentes, pero todos contribuirán a determinar el número medio y aun la existencia de las especies. En algunos casos
se puede demostrar que obstáculos muy diferentes obran sobre la misma especie en distintas regiones. Cuando miramos las plantas y arbustos que cubren una ribera enmarañada nos sentimos tentados a atribuir el número y clases proporcionales a la que llamamos casualidad. ¡Que opinión más equivocada! Todos han oído decir que cuando se tala un bosque norteamericano surge una vegetación muy distinta; pero se ha observado que antiguas ruinas indias en el sur de los Estados Unidos, que antes deben haber sido despejadas de árboles, exhiben ahora la misma hermosa variedad y proporción de todas clases que la selva virgen circundante. ¡Que lucha debe haberse desarrollado durante largos siglos entre las diversas clases de árboles que cada año disceminan sus semillas por millares! ¡Que guerra entre insecto e insecto, entre insectos, caracoles y otros animales con bestias de presa, todos procurando aumentar, todos alimentándose unos de otros, o de los árboles, de sus semillas y sus retoños, o de las plantas que alfombraban antes el suelo, entorpeciendo así el crecimiento de los árboles! Arrójese un puñado de plumas y todas caerán al suelo de acuerdo con leyes definidas; pero ¡qué simple es el problema de dónde ha de caer cada una comparado con el de la acción y reacción de las innumerables plantas y animales que han determinado, en el curso de los siglos, el número y las clases proporcionales de los árboles que ahora crecen en las antiguas ruinas indias!
La dependencia de un ser orgánico de otro, como de un parásito de su presa se produce generalmente entre animales alejados en la escala de la naturaleza. A veces pasa lo mismo con aquellos de los que se podía decir estrictamente que luchan por su existencia, por ejemplo, las langostas y los cuadrúpedos que viven de hierba. Pero la lucha será casi invariablemente más severa entre individuos de la misma especie, porque frecuentan las mismas regiones, necesitan el mismo alimento y están expuestos a los mismos peligros. En caso de variedades de las misma especie, la lucha por lo general será casi igualmente intensa, y a veces vemos que la contienda se decide pronto: por ejemplo, si diversas variedades de trigo son sembradas juntas, y las semillas mezcladas son plantadas de nuevo, algunas de las variedades, las que mejor se adaptan al suelo o al clima o son más fecundas por naturaleza vencerán a las otras y rendirán más simiente, de modo que en pocos años suplantaran a las demás variedades. Para mantener una provisión mixta de variedades tan próximas como las

68
arvejas dulces de varios colores hay que cosecharlas cada año por separado y mezclar en su debida proporción las semillas, porque de otro modo las clases más débiles disminuirán constantemente en número hasta desaparecer. También para eso con las variedades de ovejas; se ha afirmado que ciertas variedades de montaña harán morir de hambre a otras variedades de ovejas montañosas, de modo que no pueden ser mantenidas juntas. Igualmente se producido ese resultado por tener juntas variedades distintas de la sanguijuela medicinal. Hasta puede dudarse que las variedades de cualquiera de nuestras plantas o animales domésticos tengan tan exactamente la misma fuerza, hábitos y constitución que pudieran mantenerse las proporciones originales de un conjunto mixto (impidiendo los cruzamientos) durante una media docena de generaciones, si se les permitiera luchar entre si como los seres en estado de naturaleza, y s las semillas o las crías no fueran preservadas anualmente en la debida proporción.
LA LUCHA POR LA CIDA ES MÁS INTENSA ENTRE INDIVIDUOS Y VARIEADES DE LA MISMA ESPECIE.
Como las especies del mismo género suelen tener, aunque de ningún modo la tienen en todos los casos, mucha semejanza de hábitos y constitución y siempre de estructura, la lucha será generalmente más intensa entre ellas, si entran en competencia, que entre las especies de géneros distintos. Lo vemos en la reciente difusión por parte de los Estados Unidos, de una especie de golondrina que ha hecho disminuir otras especies. El reciente aumento del tordo en ciertas partes de Escocia ha ocasionado la disminución del zorzal (de la familia de los tordos.) Es le turdus musicus de los zoólogos. ¡ Cuan a menudo oímos decir de una especie de rata ha ocupado el lugar de otra especie en los climas más diferentes! En Rusia la pequeña cucaracha asiática ha desplazado en todas partes a su gran congénere. En Australia la abeja de colmena importada esta exterminando rápidamente a la pequeña abeja nativa, que no tiene aguijón. Una especie de mostaza según se ha observado, suplanta a otra especie, y así en muchos casos distintos. Podemos ver confusamente porque la competencia ha de ser más encarnizada entre formas relacionadas que ocupan casi el mismo lugar en la economía de la naturaleza; pero probablemente en ningún caso podríamos decir con precisión porque una especie ha sido
victoriosa sobre la otra en la gran batalla de la vida.
Un corolario de la mayor importancia puede deducirse de las observaciones anteriores, a saber, de la estructura de todo ser orgánico esta relacionada, del modo más esencial aunque frecuentemente oculto, con la de todos los otros seres orgánicos con los que entra en la competencia por la comida o la residencia, o de los que tienen que escapar, o que les sirve de presa. Esto es evidente en la estructura de los dientes y garras del tigre; en la de las patas y garfios del parásito que se prende al pelo el tigre. Pero en la semilla hermosamente emplumada del diente de león y en las patas chatas y peludas del escarabajo acuático la relación parece al principio limitarse a los elementos, el aire y el agua. Sin embargo, no cabe duda de que la ventaja de las semillas emplumadas esta en la más estrecha relación con el hecho de que la tierra ya se haya espesamente alfombrada de otras plantas, de modo que las semillas puedan ser ampliamente distribuidas y caigan en terreno no ocupado. En el escarabajo de agua la estructura de las patas, tan bien adaptada para la zambullida, le permite competir con otros insectos acuáticos, cazar sus propias presas y librarse de ser presa de otros animales.
La cantidad de elemento nutritivo depositado dentro de las semillas de muchas plantas no parece tener, a primera vista, relación con otras plantas. Pero por el vigoroso conocimientos de plantas jóvenes nacidas de tales semillas, como arvejas y habas cuando son sembradas en medio de hierba larga puede sospecharse que la utilidad principal de la sustancia nutritiva que hay en la semilla es favorecer el crecimiento de la planta nueva que tiene que competir con otras plantas ya en vigoroso crecimiento a su alrededor.
Mírese una planta que se encuentra en el centro de la región que abarca su especie, ¿por qué no duplica o cuadruplica su número? Sabemos que puede soportar perfectamente un poco más de frío o calor, de humedad o sequía, porque en cualquiera otra parte aparece en regiones ligeramente más cálidas frías, más húmedas o secas. En este caso vemos claramente si deseamos dar con la imaginación a esta planta el poder de aumentar en número, tenemos que ofrecerle alguna ventaja sobre sus competidoras o sobre los animales que la comen.
En los confines de la zona geográfica que abarca, un cambio de constitución con respecto al clima seria evidentemente una

69
ventaja para nuestra planta; pero tenemos razones para creer que muy pocas plantas o animales abarcan zonas tan extensas que sean destruidas únicamente por el rigor del clima. Solamente cuando llegamos a los últimos confines de la vida, en las regiones árticas o al borde de un desierto total, cesa la competencia.
La tierra puede ser extremadamente fría o seca, pero todavía habrá competencia entre unas pocas especies o entre los ejemplares de la misma por los sitios más cálidos o más húmedos.
Por esto podemos explicarnos que cuando una planta o animal son colocados en nuevo país, entre nuevos competidores, generalmente las condiciones de su vida cambiarían de modo esencial aunque el clima sea exactamente el mismo que en su región anterior. Para que su número aumente en su nueva región, tendríamos que modificarlos en sentido diverso que en su país natal, porque tendríamos que darle alguna ventaja sobre un conjunto diferentes de competidores o enemigos.
Es conveniente imaginarse una ventaja cualquiera que deberíamos dar una especie sobre otra. Probablemente en ningún caso sabríamos que hacer. Esto debiera bastar para convencernos de nuestra ignorancia sobre las relaciones mutuas de todos los seres orgánicos, convicción tan necesaria como difícil de adquirir. Todo lo que podemos hacer es tener bien presente que cada ser orgánico se esfuerza por aumentar en progresión geométrica; que cada uno de ellos, en diferentes periodos de su vida, durante alguna estación del año, durante cada generación o con intervalos, tiene que luchar por su vida y ser destruido en gran cantidad. Cuando reflexionamos sobre esta lucha, podemos consolarnos pensando que la guerra en la naturaleza no es incansante, que no siente el miedo, que la muerte generalmente es rápida, y que los vigorosos, los sanos y los felices sobreviven y se multiplican.
CAPITULO IV
LA SELECCIÓN NATURAL O LA SUPERVIVENCIA DE LOS MÁS APTOS.
SELECCIÓN NATURAL; SU PODER COMPARADO CON LA SELECCIÓN DEL HOMBRE; SU PODER SOBRE CARACTERES DE ESCASA IMPORTANCIA; SU INFLUENCIA EN TODAS LAS EDADES Y EN LOS DOS SEXOS.
¿Cómo obrara la lucha por la existencia, brevemente tratado en el último capitulo, con respecto a la variación? ¿Puede aplicarse al estado de naturaleza el principio de selección, que hemos visto ser tan poderoso en manos el hombre? Creo que lo veremos obrar del modo más eficaz. Téngase presente el infinito número de variaciones ligeras y diferencias individuales que se manifiestan en nuestros productos domésticos, y en menor grado en los que se hallan en estado de naturaleza, así como la fuerza de la tendencia hereditaria. En la domesticidad, puede decirse con justicia que toda la organización se vuelve en cierto modo plástica. Pero la variabilidad que encontramos casi universalmente en nuestros productos domésticos no es producida directamente, como bien han observado Hooker y Asa Gray, por el hombre; el no puede ni originar variedades ni impedir que ocurran; puede, si, preservar y acumular las que ocurren. Expone sin querer los seres orgánicos a nuevas y cambiantes condiciones de vida, y se produce la variabilidad; pero cambios similares de condiciones podrían ocurrir y efectivamente ocurren en estados de naturaleza. Tengamos también presente cuan infinitamente complejas y estrechamente adaptadas son las relaciones mutuas de todos los seres orgánicos entre si y con sus condiciones físicas de vida; y en consecuencia que diferencias infinitamente variadas de estructura podrían ser útiles a cada ser bajo condiciones alteradas de vida ¿Puede entonces parecer improbable, en vista de que han ocurrido indudablemente variaciones útiles al hombre, que otras variaciones útiles a de algún modo a cada ser en la grande y compleja batalla de la vida ocurran en el curso de muchas generaciones sucesivas? Y si ocurren ¿podemos dudar (recordando que nacen muchos más ejemplares de los que pueden sobrevivir) de que los ejemplares que tengan alguna ventaja, por leve que sea, sobre otros tendrán las mejores probabilidades de vivir y dejar descendientes semejantes?. Por otra parte, podemos estar seguros de que cualquier variación que sea perjudicial en el más ínfimo grado seria rígidamente destruida. Esta preservación de diferencias y variaciones individuales favorables y la destrucción de las perjudiciales es lo que yo he llamado selección natural o supervivencia de los más aptos. Las variaciones que no son útiles ni dañosas no serian afectadas por la selección natural y quedarían ya como un elemento fluctuante, como vemos quizá en ciertas especies polimorfas, o finalmente se fijarían debido a la naturaleza del organismo y de las condiciones.

70
Varios escritores han interpretado mal o formulado objeciones a la expresión selección natural. Algunos han llegado a imaginar que la selección natural provoca la variabilidad, cuando por el contrario implica solamente la preservación de esas variaciones que surgen y son beneficiosas para el ser en sus condiciones dadas de vida. Nadie se opone a que los agricultores hablen de los potentes efectos de la selección del hombre, y en este caso las diferencias individuales dadas por la naturaleza, que el hombre elige por algún motivo, tienen necesariamente que ocurrir primero. Otros han alegado que el termino selección implica una elección consciente por parte de los animales que resultan modificados, ¡y hasta se ha argüido que, como las plantas no tienen voluntad, la selección no es aplicable a ellas! En el sentido literal de la palabra no cabe duda de que selección natural es una expresión falsa; pero ¿quién se ha opuesto nunca a que los químicos hablen de las afinidades electivas de los diversos elementos? Y sin embargo no se puede decir estrictamente que un ácido elija la base con que mejor se combina. Se ha dicho que hablo de la selección natural como de un poder o deidad activa; pero ¿quién se opone a que un autor hable de la fuerza de gravedad como de la que gobierna los movimientos de las plantas? Todos saben lo que se quiere decir y aparece implícitamente en esas expresiones metafóricas; son casi indispensables para la brevedad. Del mismo modo, es difícil evitar la personificación de la naturaleza; pero por naturaleza solo entiendo la acción sumada y el producto de muchas leyes naturales, y por leyes la secuencia de sucesos tal como ha sido comprobada por nosotros. Familiarizándose un poco con el tema, se olvidaran estas objeciones superficiales.
Comprenderemos mejor el curso probable de la selección natural tomando el caso de un país que sufre algún ligero cambio físico, por ejemplo, de clima. Las cantidades proporcionales de sus moradores cambiaran casi de inmediato, y probablemente algunas especies se extinguirán. Podemos concluir, de lo que hemos visto sobre la intima y compleja manera en que están vinculados los habitantes de cada país, que cualquier cambio en las cantidades proporcionales de sus moradores, independientemente del cambio del propio clima, afectara seriamente a todos. Si el país fuera abierto en sus fronteras, seguramente inmigrarían nuevas formas, y esto también perturbaría seriamente las relaciones de algunos de los habitantes anteriores. Recuérdese cuan poderosa ha
demostrado ser la influencia de un solo árbol o mamífero introducido. Pero en el caso de una isla o país parcialmente rodeado por barreras en el que no podrían entrar libremente formas nuevas y mejor adaptadas, tendríamos ciertamente en la economía de la naturaleza que serian ocupados mejor si algunos de los moradores originales se modificaran de uno u otro modo; pero si la zona hubiese estado abierta a la inmigración, esos mismos lugares habrían sido capturados por los intrusos. En tales casos, modificaciones ligeras de que algún modo favorecieran a los individuos de alguna especie adaptándolos mejor a sus condiciones alteradas, tenderían a preservarse y la selección natural tendría amplio campo para la obra de mejoramiento.
Tenemos buenas razones para creer, como se ha demostrado en el capitulo I, que los cambios en las condiciones de vida producen una tendencia al aumento de variabilidad, y en los casos mencionados, las condiciones han cambiado, y esto evidentemente seria favorable a la selección natural porque daría mejores probabilidades de que ocurran variaciones provechosas. Si ellas no ocurren la selección natural no puede hacer nada. Bajo el termino “variaciones” nunca debe olvidarse que se alude a simples diferencias individuales. Así como el hombre puede obtener un gran resultado con sus animales y plantas domesticas agregando en algún sentido determinado diferencias individuales, lo mismo podría lograr la selección natural, pero de un modo mucho más fácil porque tiene incomparablemente más tiempo para la selección. Y no creo yo que sea necesario un gran cambio físico, como por ejemplo en el clima, ni un grado inusitado de aislamiento que impida la inmigración para que queden nuevos lugares desocupados que a la selección natural corresponde llenar mejorando algunos de los diversos habitantes. Pues como todos los moradores de cada país están luchando entre si con fuerzas exquisitamente equilibradas, una modificación levisima en la estructura o costumbres de una especie bastaría con frecuencia para darle una ventaja sobre las otras; y nuevas modificaciones de la misma clase aumentarían aun la ventaja mientras la especie continuara bajo las mismas condiciones de vida y aprovechara medios similares de subsistencia y defensa. No se puede mencionar ningún país en que todos los habitantes nativos estén ahora tan perfectamente adaptados unos a otros y a las condiciones físicas bajo las cuales viven que ninguno de ellos pudiera mejorar o adaptarse mas. Pues en todos los países,

71
productos nativos han sido vencidos por productos importados hasta el punto de que han permitido a algunos extraños tomar firme posesión de su tierra. Y como los extraños han vencido así en cada país a algunos de los productos nativos, podemos concluir sin temor que estos podrían haberse modificado beneficiosamente para resistir mejor a los intrusos.
Si el hombre puede producir y ha producido ciertamente grandes resultados con sus medio metódicos o inconscientes de selección ¿qué no podrá hacer la selección de la naturaleza? El hombre solo puede obrar sobre caracteres externos y visibles. La naturaleza, si se me permite personificar la preservación natural o supervivencia de los más aptos, no se cuida de las apariencias, salvo en cuando son útiles a un ser. Puede obrar sobre cada órgano interno, sobre cada matriz de diferencia constitucional, sobre todo el mecanismo de la vida. El hombre elige solamente para su propio bien: la naturaleza solamente para bien del ser afectado. Todo carácter seleccionado e plenamente ejercitado por ella, como lo implica el hecho de que lo haya elegido. El hombre mantiene a los productos nativos de muchos climas en el mismo país; rara vez hace ejercitarse cada carácter seleccionado de un modo especial y adecuado; alimenta a una paloma de pico largo y a una de pico corto con la misma comida; no hace ejercitarse al cuadrúpedo de lomo largo o patas largas de ninguna manera especial; somete al mismo clima ovejas de lana corta y de lana larga. No permite que los machos más vigorosos luchen por las hembras. No destruye rígidamente a todos los animales inferiores, sino que protege a todos, en cada estación, hasta donde puede. A menudo empieza la selección con algunas forma semimonstruosa; o por lo menos con alguna modificación lo suficientemente destacada para llamar la atención o ser evidentemente útil. En estado de naturaleza, las más leves diferencias de estructuras o constitución bien podrían alterar la balanza delicadamente equilibrada en la lucha por la existencia, y por ello ser preservadas. ¡ Cuan fugaces son los deseos y esfuerzos del hombre! ¡Cuán breve su tiempo! Y en consecuencia, ¡cuan pobres sus resultados comparados con los de su naturaleza durante periodos geológicos enteros! ¿Podemos maravillarnos entonces de que los productos de la naturaleza sean mucho más genuinos en sus caracteres que los productos del hombre; que estén infinitamente mejor adaptados a las más complejas condiciones de vida y lleven claramente el sello de una elaboración mucho más elevada?
Metafóricamente, puede decirse que la selección natural escudriña día por día y hora por hora en todo el mundo las más leves variaciones, rechazando las que son malas, preservando y aumentando las que son buenas, trabajando en silencio e insensiblemente, dondequiera y cuando quiera que se presente la oportunidad, en el mejoramiento de cada ser orgánico en relación con sus condiciones orgánicas e inorgánicas de vida. No percibimos nada de estos lentos cambios en marcha hasta que la mano del tiempo ha marcado un transcurso de edades, y en nuestro conocimiento de edades geológicas transcurridas en el lejano pasado es tan imperfecto que solamente vemos que las formas de vida son ahora muy diferentes de lo que fueron antes. Para que se efectúe en una especie un alto grado de modificación, la variedad, una vez formada, deberá nuevamente, quizá después de un largo intervalo, variar o presentar diferencias individuales del mismo carácter favorable que antes; y estas deberán ser preservadas a su vez, y así sucesivamente, paso a paso. Dado que diferencias individuales de la misma clase se repiten perpetuamente, difícilmente podrá considerarse que esta sea una suposición injustificada. Pero su verdad solo puede ser apreciada viendo hasta donde concuerda y explica los fenómenos generales de la naturaleza. Por otra parte, la creencia ordinaria de que el grado de variación posible es una cantidad estrictamente limitada, también es una simple suposición.
Aunque la selección natural puede obrar solamente por intermedio y para beneficio de cada ser, ciertos caracteres y estructuras que tendríamos propensión a considerar insignificantes, pueden ser afectados así. Cuando vemos que son verdes los insectos que comen hojas y moteados de gris los que comen cortezas; que la perdiz de los Alpes es blanca en invierno, la perdiz de Escocia tiene el color del brezal, hemos de creer que esos tintes son útiles para tales aves e insectos en cuanto los preservan del peligro. La gallineta, si no fuera destruida en algunos periodos e sus vida, aumentaría infinitamente en número; se sabía que padece mucho de aves de presa, y los gavilanes se guían por la vista hacia su presa, hasta el punto de que en el continente se previene a las personas que no tengan palomas blancas pues son las más expuestas a destrucción. De ahí que la selección natural pueda operar dando el color adecuado a cada clase de gallineta y manteniendo ese color, una vez adquirido, firme y constante. Tampoco debemos pensar que la destrucción ocasional de un animal de

72
cualquier color determinado produciría poco efecto; debemos recordar cuan esencial es destruir en un rebaño de ovejas blancas todo cordero que tenga la menor traza de negro. Hemos visto como el color de los cerdos que se alimentan de la “raíz de pintura” En Virginia determina si han de vivir o morir. En las plantas, la vellosidad del fruto y el color de su carne son considerados por los botánicos como caracteres de la más trivial importancia; sin embargo, nos dice un excelente horticultor, Downing, que en los Estados Unidos las frutas de cascara lisa sufren mucho más de cierto escarabajo, el curculio, que las que tienen pelusa; que las ciruelas purpúreas sufren mucho más de cierta enfermedad que las ciruelas amarillas; mientras otra enfermedad ataca a los duraznos amarillos que a los de otro color. Si con ayuda del arte esas leves diferencias tienen mucha importancia en el cultivo de diversas variedades, en estado de naturaleza, donde los árboles tendrían que luchar con otros árboles y con un ejercito de enemigos, tales diferencias determinarían eficazmente cual variedad de alguna fruta, si lisa o vellosa, amarilla o purpúrea, habrá de prosperar.
Observando muchos pequeños puntos de diferencia entre las especies que hasta donde nos permite juzgar nuestra ignorancia, parecen completamente faltos de importancia, no debemos olvidar que el clima, el alimento, etc., han producido sin duda algún efecto directo. También es necesario recordar que, debido a ley de correlación, cuando varía una parte y las variaciones se acumulan por selección natural, han de producirse otras modificaciones, a menudo de la naturaleza más inesperada. Así como vemos en la domesticidad que las variaciones que aparecen en cualquier periodo particular de vida tienden a reaparecer en la descendencia en el mismo periodo – por ejemplo, en la forma, tamaño y sabor de las semillas de las muchas variedades de nuestras plantas culinarias y agrícolas; en las etapas de oruga y capullo de las variaciones de gusano e seda; en los huevos de las gallinas y en el color del plumón de sus pollos; en los cuernos de nuestros ovinos y vacunos cuando son casi adultos- del mismo modo en estado de naturaleza la selección natural podría actuar, y modificar a seres orgánicos en cualquier edad, mediante acumulación de variaciones provechosas para esa edad y mediante su herencia a la edad correspondiente.
Si aprovecha a una planta el que sus semillas sean cada vez más ampliamente dispersadas por el viento no veo mayor dificultad para que
esto se produzca por selección natural de la que veo para que un replantador de algodón aumente mejore por selección el vellón de sus algodoneros. La selección natural puede modificar y adaptar las larvas de un insecto a una veintena de contingencias totalmente distintas de las que afectan al insecto adulto, y esas modificaciones, por la correlación, pueden afectar su estructura el adulto. Recíprocamente, las modificaciones en el adulto pueden afectar la estructura de larva, pero en todos los casos la selección natural asegurara que no sean perjudiciales, pues si lo fuesen la especie se extinguiría.
La selección natural modificara la estructura de los hijos con respecto a los padres, y de los padres con respecto a los hijos. En los animales gregarios, adaptara la estructura de cada ejemplar, al beneficio de toda la comunidad, si la comunidad se beneficia con el cambio seleccionado. Lo que no puede hacer la selección natural es modificar la estructura de una especie sin dar alguna ventaja, para beneficio de otra especie; y aunque en la historia natural se pueden encontrar afirmaciones en ese sentido, no logro encontrar ni un caso que resista la investigación. Una estructura utilizada una sola vez en la vida de un animal, si es muy importante para el, podría ser modificada hasta cualquier grado por la selección natural; por ejemplo, las grandes mandíbulas que tienen algunos insectos, utilizadas exclusivamente para abrir el capullo, o la dura punta del pico en pájaros no salidos de la cascara aun, que sirve para romperla. Se ha afirmado que de las mejores palomas volteadoras de pico corto son más las que mueren en el huevo que las que logran salir de él. De modo que los criadores ayudan en el acto del nacimiento, ahora bien, si la naturaleza tuviera que hacer muy corto el pico de una paloma adulta, para ventaja de la propia ave, el proceso de modificación seria muy lento y se produciría simultáneamente la más rígida selección de todas las crías que dentro del huevo tuvieran el pico más duro y fuerte, porque las de pico débil morirían irremisiblemente; o bien podrían resultar seleccionados cascarones más delicados y fáciles de quebrar, ya que el espesor de la cascara del huevo varia, según se ha notado, como cualquier otra estructura.
Quizá sea oportuno observar aquí que en todos los seres debe de producirse mucha destrucción fortuita que tenga poca o ninguna influencia sobre el curso de la selección natural. Por ejemplo, gran número de huevos o semillas son devorados anualmente, y estas

73
podrían ser modificadas por selección natural solamente si variaran de alguna manera que las protegiera de sus enemigos. Sin embargo, muchos de esos huevos o semillas, si no hubieran sido destruidos, habrían dado quizá ejemplares mejor adaptados a sus condiciones de vida que cualquiera de los resultaron sobrevivientes. De este modo, también un gran número de animales y plantas adultos, sean o no lo mejor adaptados a sus condiciones, deben ser destruidos anualmente por causas accidentales que de ningún modo serian mitigadas por ciertos cambios en la estructura o constitución que en otros sentidos habrían sido beneficiosos para la especie. Pero por intensa que sea la destrucción de los adultos, si el número de los que pueden existir en cualquier distrito no es totalmente reprimido por tales causas, o también, aunque la destrucción de huevos o semillas sea tan grande que solamente subsista la centésima o milésima parte, si hay variabilidad en dirección favorable, los ejemplares mejor adaptados de los que sobrevivan tenderán a propagar sus iguales en mayor número que los menos aptos. Si el número fuese totalmente sofocado por las causas recién indicadas, como habrá ocurrido a menudo, la selección natural seria impotente en ciertas direcciones favorables; pero esta no es objeción valida contra su eficacia en otras circunstancias y en otros sentidos, porque estamos lejos de tener razones para suponer que son muchas las especies que se modifican y mejoran al mismo tiempo y en la misma región.
SELECCIÓN SEXUAL
Puesto que en la domesticidad aparecen con frecuencia particularidades en un sexo que quedan hereditariamente unidas al mismo, igual sucederá, sin duda, en la naturaleza.
De esta modo se hace posible que los dos sexos se modifiquen, mediante selección natural, en relación con sus diferentes costumbres, como es muchas veces el caso, o que un sexo se modifique con relación al otro, como ocurre comúnmente. Esto me lleva a decir algunas palabras acerca de lo que he llamado selección sexual. Esta forma de selección depende, no de una lucha por la existencia en relación con otros seres orgánicos o con condiciones externas, sino de una lucha entre los individuos de un sexo – generalmente, los machos- por la posesión del otro sexo. El resultado no es la muerte del competidor desafortunado, sino que deja poca o ninguna descendencia. La selección sexual
es, por tanto, menos rigurosa que la selección natural. Generalmente, los machos más vigorosos, lo que están mejor adaptados a su situación en la naturaleza, dejaran más descendencia; pero en mucho casos la victoria depende no tanto del vigor natural como de la posesión de armas especiales limitadas al sexo masculino. Un ciervo sin cuernos, un gallo sin espolones, tendrán pocas probabilidades de dejar númerosa descendencia. La selección sexual, dejando siempre criar al vencedor, pudo, seguramente dar valor indomable, longitud a los espolones, fuerza al ala para empujar la pata armada de espolón, casi del mismo modo que lo hace brutal gallero mediante la cuidadosa selección de sus mejores gallos.
Ignoro hasta que punto, en la escala de los seres naturales, llega esta ley del combate; se ha descrito que los cocodrilos riñen, rugen y giran alrededor -–omo los indios en una danza guerrera- por la posesión de las hembras. Se ha observado que los salmones machos pelean durante todo el día; los ciervos voladores (llamados también escarabajos cornudos, son coleópteros cuyo macho tiene dos mandíbulas muy desarrolladas y parecidas a las astas del ciervo.) Machos a veces llevan heridas de las enormes mandíbulas de los otros machos; el inimitable observador Fabre ha visto muchas veces los machos de ciertos himenopteros riñendo por una hembra determinada que esta posada al lado, espectadoras en apariencia indiferente de la lucha, la que se retira después con el vencedor. La guerra es quizá más severa entre los machos de los animales polígamos, y parece que estos cuentan muy frecuentemente con armas especiales. Los machos de los carnívoros están siempre bien armados, aun cuando estos y otros pueden disponer de medios especiales de defensa mediante la selección natural, como la melena del león o la mandíbula ganchuda del salmón macho, pues tan importante puede ser para la victoria el escudo como la espada o la lanza.
Entre las aves, la contienda es muchas veces de carácter pacifico. Todos los que se han ocupado de este asunto creen que entre los machos de númerosas especies existe gran rivalidad por atraer con su canto a las hembras. El tordo rupestre de Guayana, las aves del paraíso y algunas otras se reúnen, y los machos sucesivamente, despliegan con el más minucioso cuidado y exhiben de la mejor manera su esplendoroso plumaje; además ejecutan extraños movimientos ante las hembras, que, asistiendo como espectadoras, escogen al fin el compañero más atractivo. Lo

74
que han prestado mucha atención a las cautivas saben perfectamente que estas, con frecuencia, tienen preferencias y aversiones individuales; R. Heron ha descrito como un pavo real manchado era sumamente atractivo para todas sus pavas. Si el hombre es capaz en corto tiempo de dar hermosura y porte elegante a sus gallinas bantam conforme a su estándar o tipo de belleza, no hay ninguna razón legítima para dudar de que las aves hembras, eligiendo durante miles de generaciones los machos más hermosos y melodiosos según sus tipos de belleza, puedan producir un efecto señalado. Algunas leyes muy conocidas respecto al plumaje de las aves machos y hembras, en comparación del plumaje de los polluelos, pueden explicarse, en parte, mediante la selección sexual sobre variaciones que se presentan en diferentes edades y se transmiten solo a los machos, o a los dos sexos, en las edades correspondientes.
Así es que, cuando los machos y hembras tienen las mismas costumbres generales, pero difieren en conformación, color o adorno, estas diferencias han sido producidas principalmente por selección sexual, es decir: mediante individuos machos que han tenido en generaciones sucesivas alguna ligera ventaja sobre otros machos, en sus armas, medios de defensa o encantos, que han transmitido solamente a su descendencia masculina. Sin embargo, no quisiera atribuir todas las diferencias sexuales a esta acción, pues en los animales domésticos vemos surgir en el sexo masculino y quedar ligadas a él particularidades que evidentemente no han sido acrecentadas mediante selección por el hombre. El mechón de filamentos en el pecho del pavo salvaje no puede tener ningún uso, y es dudosos que sea ornamental a los ojos de la hembra; realmente, si el mechón hubiese aparecido en estado domestico se le habría calificado como monstruosidad.
EJEMPLOS DE LA ACCION DE LA SELECCIÓN NATURAL O SUPERVIVENCIA DE LOS MÁS APTOS.
A fin de explicar como obra, según creo, la selección natural pediré permiso para dar uno o dos ejemplos imaginarios. Tomemos el caso de un lobo que se ceba en varios animales, alcanzando algunos por la astucia, otros por la fuerza y otros por la velocidad; y supongamos que su presa más ligera, un gam, por ejemplo, hubiera aumentado en número por algún cambio en el país, o que hubiera decrecido otra especie durante la estación del
año en que el lobo estaba más apremiado por la necesidad de comida. Bajo tales circunstancias los lobos más delgados tendrían las mejores probabilidades de sobrevivir y ser así preservados o seleccionados, siempre que conservasen bastante robustez para dominar a su presa en este u otro periodo del año en que se vieran obligados a cazar otros animales. No veo más razones para dudar de que ese seria el resultado que para dudar que el hombre pueda mejorar la velocidad de sus galgos mediante una selección metódica y cuidadosa o con esa clase de selección inconsciente que se produce porque cada hombre procura conservar sus mejores perros, aunque no piense en modificar la raza. Añadiré que, según Mr. Pierce, hay dos variedades de lobo en las montañas de Catskill, Estados Unidos: una que tiene forma ligera, parecida al galgo y persigue a los gamos; otra más maciza, con patas más cortas, que ataca preferentemente a los rebaños de los pastores.
Debe observarse que en el ejemplo anterior hablo de los lobos individualmente más flacos y no de ninguna variación muy marcada que se haya preservado. En ediciones anteriores de esta obra hablaba yo a veces como si la última alternativa hubiese ocurrido con frecuencia. Veía la gran importancia de las diferencias individuales y esto me llevo a tratar de lleno los grandes resultados de la selección inconsciente efectuada por el hombre, que depende de la preservación de todos los ejemplares más o menos valiosos y de la destrucción de los peores. Veía también que la preservación, en estado de naturaleza, de cualquier desviación ocasional de estructura, tal como una monstruosidad, seria un caso raro, y que aunque se preservarse al principio se perdería generalmente por el intercruzamiento ulterior con ejemplares normales. Sin embargo, hasta que leí un competente y útil articulo publicado en la North British Review (1867) no aprecie cuan raro es que se conserven las variaciones individuales, sean leves o muy marcadas. El autor toma el caso de un par de animales que producen durante su vida doscientos hijos, de los cuales, por causas diversas de destrucción, solamente dos, por termino medio, alcanzan a procrear.
Este es un cálculo más bien extremado para la mayoría de los animales superiores, pero de ningún modo para los organismos inferiores. Después demuestra que si naciera un solo individuo que viajara de alguna manera, dándole el doble de probabilidades de vida que a los otros ejemplares, aun así las

75
probabilidades serian en su mayoría contrarias a que sobreviviera. Suponiendo que lo lograra y procreara y que la mitad de sus hijos heredara la variación favorable todavía, como dice el articulista, los hijos apenas tendrían, un poco más de probabilidades de vivir y procrear; y esta probabilidad iría disminuyendo en las generaciones sucesivas. Creo que no puede discutirse la exactitud de estas observaciones. Por ejemplo, si alguna ave de alguna clase pudiera procurarse alimento con más facilidad por tener el pico encorvado y si naciera una con el pico encorvado y si naciera una con el pico muy encorvado que por ello prosperase, habría con todo muy pocas perspectivas de que ese único ejemplar perpetuase su forma con exclusión de la forma común; pero ha juzgar por lo que hemos visto en la domesticidad, difícilmente podrá dudarse que ese resultado se produciría por la preservación, durante muchas generaciones, de ejemplares con el pico más o menos encorvado y por la destrucción de un número todavía mayor de los que tuviesen el pico recto.
Sin embargo, no debe olvidarse que ciertas variaciones muy marcadas, que nadie clasificaría como meras diferencias individuales, se repiten a menudo debido a que una organización semejante es afectada de modo parecido, de lo cual podrían sacarse muchos ejemplos entre nuestros productos domésticos. En tales casos, si el ejemplar que varia no transmitiera realmente a sus descendientes el carácter recién adquirido. Indudablemente les transmitiría, mientras se mantuviesen las condiciones existentes, una fuerte tendencia a variar del mismo modo.
Tampoco puede haber dudas de que la tendencia variar del mismo modo ha sido a menudo tan fuerte que todos los ejemplares de la misma especie han resultado modificados similarmente sin ayuda de ninguna forma de selección. O puede haber sido afectada así solamente la tercera, quinta o décima parte de los ejemplares, de lo cual también podrían darse varios ejemplos. Por eso Graba calcula que aproximadamente un quinto de las crías de las islas Feroe pertenece a una variedad tan marcada que antes se le clasificaba como especie distinta bajo el nombre de uria lacrymans. En casos semejantes, si las variaciones fueran beneficiosas, la forma original pronto seria suplantada por la forma modificada, debido a la supervivencia de los más aptos. Tendré que volver sobre los efectos del cruzamiento para eliminar variaciones; pero aquí puede observarse que la mayoría de los animales y
plantas se mantienen en su propia tierra y no viajan innecesariamente; lo vemos hasta en las aves migradoras que casi siempre vuelven al mismo punto. En consecuencia cada variedad recién formada seria el principio generalmente local, como parece ser la regla con las variedades de estado de naturaleza; de modo que ejemplares similarmente modificados no tardarían en existir juntos en un cuerpo reducido y a menudo procrearían juntos. Si la nueva variedad tuviera éxito en la batalla por la vida, difundiría lentamente desde una región central, compitiendo y venciendo a los ejemplares no cambiados, en un círculo cada vez mayor.
Quizá valga la pena dar otro ejemplo más completo de la acción de la selección natural. Ciertas plantas segregan un jugo, al parecer para eliminar algo dañoso a la savia; así lo hacen por ejemplo, ciertas leguminosas con glándulas colocadas en la base de las estipulas o el laurel común con glándulas situadas en la cara posterior de la hoja. Este jugo, aunque exiguo en cantidad, es sorbido afanosamente por los insectos, pero sus visitas no benefician en nada a la planta. Ahora supongamos que el jugo o néctar fuera segregado desde el interior de las flores de cierto número de plantas de cualquier especie. Los insectos, al buscar el néctar, se empolvarían de polen, y a menudo lo transportarían de una flor a otra. Las flores de dos ejemplares distintos de la misma especie resultarían cruzadas así, y el acto del cruzamiento como puede probarse, da origen a vigorosos vástagos que en consecuencia tendrían las mejores probabilidades de prosperar y sobrevivir. Las plantas que producen flores con las mayores glándulas o nectáreos, las que segregan más néctar, serian las más visitadas por los insectos y por lo tanto las cruzadas con más frecuencia; y así a la larga ganarían a la ventaja y formarían una variedad local. También en las flores que tuvieran estambres y pistilo, en relación con el tamaño y hábitos de los insectos particulares que las visitan, dispuestos en tal forma que favoreciesen de algún modo el transporte de polen, resultarían igualmente favorecidas. Podríamos haber tomado el caso de insectos que visitan a las flores para recolectar polen en lugar de néctar, y como el polen solo tiene el fin de fecundar, su destrucción parece ser una simple perdida para la planta; pero si un poco de polen fuera llevado, al principio de vez en cuando, y luego habitualmente de flor a flor y se produjese así un cruzamiento, aunque nueve décimas partes del polen resultan destruidas todavía quedara una gran ventaja

76
para la planta despojada así, y los ejemplares que produjesen más y más polen y tuvieran las antenas más y más grandes, serian seleccionados.
Cuando nuestra planta, por el proceso mencionado, largamente proseguido, resultase muy atrayente para los insectos, éstos sin intención de su parte llevarían regularmente polen de flor a flor, y que efectivamente lo hacen así podríamos probarlo con muchos hechos notables. Expondré solamente uno, que ilustra igualmente un paso para la separación de los sexos de las plantas. Algunos agrifolios producen solamente flores machos que tienen cuatro estambres productores de una cantidad más bien pequeña de polen, y un pistilo rudimentario; otros agrifolios producen solamente flores hembras; estas tienen pistilo de tamaño cabal y cuatro estambres con antenas encogidas, en las que no se descubre un solo grano de polen. Habiendo encontrado un árbol hembra de sesenta yardas, exactamente, de un árbol masculino, puse los estigmas de veinte flores, tomadas de ramas distintas, bajo el microscopio, y en todos ellos, sin excepción había unos pocos granos de polen, y muchos en algunos. Como durante varios días el viento había soplado del árbol hembra al macho, el polen no podía haber sido transportado de ese modo. El tiempo había sido frío y agitado, y por consiguiente, no favorable a las abejas; sin embargo cada flor hembra que examine había sido eficazmente fecundada por las abejas, que habían volado de uno a otro árbol en busca de néctar. Pero volvamos a nuestro caso imaginario: tan pronto como la planta se hubiese vuelto muy atrayente para los insectos de modo que el polen fuera regularmente transportado de flor en flor, podría comenzar otro proceso. Ningún naturalista duda de la ventaja de lo que se ha llamado división fisiológica del trabajo; por lo tanto podemos creer que habría sido ventajoso para una planta producir solamente estambres en una flor o en toda la planta, y solo pistilos en otra flor o en otra planta. En plantas cultivadas y puestas bajo otras condiciones de vida, a veces los órganos hembras se vuelven más o menos impotentes; ahora bien, si suponemos que esto ocurre aunque sea en grado muy leve, en el estado de naturaleza, entonces, como el polen ya es llevado regularmente de flor a flor y como una separación más completa de los sexos de nuestra planta habría sido beneficiosa de acuerdo con el principio de la división, los ejemplares manifiestan aumento de esa tendencia serian continuamente
favorecidos o seleccionados, hasta que al final podría producirse una separación completa de los sexos. Llevaría demasiado espacio mostrar los varios pasos, a través del dimorfismo y otros medios, por lo cual parece estar ahora desarrollándose la separación de los sexos en plantas de varias clases; pero puedo añadir que algunas de las especies de agrifolio en la América del Norte se hallan, según Asa Gray, exactamente en la condición intermedia, o como lo dice él, son más o menos dioicamente polígamas.
Volamos ahora a los insectos que se alimentan de néctar; podemos suponer que la planta que hemos visto aumentar lentamente el néctar por una selección continuada es una planta común; y que ciertos insectos necesitan principalmente su néctar para alimentarse. Podría dar muchos ejemplos para demostrar, cuanto se afanan las abejas por ganar la base de ciertas flores cuando con un poco más de trabajo podrían entrar por su boca. Teniendo en cuenta estos hechos puede creerse que, bajo ciertas circunstancias, diferencias individuales en el largo de la probóscide etc., demasiado leves para que nosotros las apreciemos, podrían aprovechar a una abeja u otro insecto, de modo que ciertos ejemplares podrían obtener su alimento con más raidez que otros; y así a las comunidades que pertenecieran prosperarían y producirían muchos enjambres dotados por herencia de las mismas peculiaridades. Los conductos tubulares de la corola del trébol rojo común y el encarnado (trifolium pratense etincarnatum) no parecen a primera vista diferir en longitud; sin embargo la abeja de colmena puede chupar fácilmente el néctar de un trébol encarnado, pero no de un trébol rojo común, que es visitado solamente por abejorros; de modo que todos los campos de trébol rojo ofrecen en vano abundante provisión de néctar precioso a la abeja de colmena. Que este néctar gusta mucho a dicha abeja es seguro; pues he visto repetidamente, aunque solo en otoño, muchas abejas de colmena sorbiendo las flores por agujeros practicados en la base del conducto tubular por abejorros. La diferencia en el largo de la corola en las dos clases de trébol que determina las visitas de la abeja de colmena debe ser insignificante, pues se me ha asegurado que cuando se ha segado el trébol rojo, las flores de la segunda cosecha son algo menores y estas son visitadas por muchas abejas de colmena. No se si esta afirmación es exacta ni si se puede confiar en otra declaración publicada, a saber, que la abeja ligur, considerada a menudo como una simple variedad de la abeja d colmena común, y que

77
se cruza fácilmente con ella, puede alcanzar y sorber el néctar del trébol rojo. Así, en un país que abundara esta clase de trébol seria una gran ventaja para la abeja de colmena tener un aguijón ligeramente más largo o hecho de un modo distinto. Por otra parte, como la fecundidad del trébol depende enteramente de que la abeja visite sus flores, si los abejorros se volviesen raros en cualquier país podría ser una gran ventaja para la planta tener una gran corola más corta o más dividida para que las abejas de colmena pudiera chuparla. Por eso puedo comprender que una flor y una abeja se transformaran lentamente, juntas o una después de otra, y se adaptaran recíprocamente del modo más perfecto por la preservación continua de todos los ejemplares que presentasen ligeras desviaciones de estructura mutuamente favorables. Se perfectamente que esta doctrina de selección natural, ilustrada en los ejemplos anteriores, esta expuesta a las mismas objeciones que fueron formuladas primero contra las nobles opiniones de Sir Charles Lyell sobre los cambios modernos de la tierra, como ejemplos para geología; pero rara vez calificamos a los agentes naturales que todavía operan de triviales o insignificantes cuando nos sirven para explicar la excavación de los valles más profundos o la formación de largas líneas de rocas terrestres. La selección natural obra solamente mediante la preservación y acumulación de pequeñas modificaciones hereditarias, cada una provechosa para el ser preservado; y así como la geología moderna ha casi desterrado opiniones como la excavación de un gran valle por una sola onda diluvial, así la selección natural desterrara la creencia en la creación continuada de nuevos seres orgánicos o de una modificación súbita y grande de su estructura.
CRUZAMIENTO DE LOS INDIVIDUOS.
Debo hacer aquí una breve disgresión. En el caso de animales y plantas de sexos separados, es obvio por supuesto, que dos ejemplares deberían siempre (con excepción de los curiosos y no muy claros casos de partenogénesis)unirse para cada nacimiento; pero cuando se trata de hermafroditas esto esta lejos de obvio. Sin embargo hay razones para creer que entre los hermafroditas, dos ejemplares, sea en ocasiones o habitualmente, concurren para la reproducción. Esta opinión fue sugerida hace largo tiempo, con vacilación, por Sprengel, Knight y Kölreuter. Pronto veremos su importancia; pero aquí debo tratar el tema
con extremada brevedad, aunque tengo preparados los materiales para una amplia discusión. Todos los animales vertebrados, todos los insectos y algunos otros grandes grupos de animales, se aparean para cada nacimiento. Investigaciones modernas han disminuido mucho el número de los presuntos hermafroditas, y de los hermafroditas verdaderos no pocos se aparean, es decir, se unen regularmente dos ejemplares para cada reproducción, que es lo único que nos interesa. Puede preguntarse que razón hay para suponer en estos casos que dos ejemplares concurran siempre a la reproducción. Como aquí es imposible entrar en detalles, he de confiarme solamente en algunas consideraciones generales. En primer ligar, he reunido un gran cuerpo de hechos y realizado tantos experimentos que demuestran, de acuerdo con la creencia casi universal de los criadores, que en los animales y plantas un cruzamiento entre variedades distintas o entre ejemplares de la misma variedad, pero de otra corriente, da vigor y fecundidad a la descendencia; y por otra parte, que el intercruzamiento estrecho disminuye mucho el vigor y la fecundidad, que estos hechos solos me inclinan a creer que es la ley general de la naturaleza que ningún ser orgánico se fecunda a sí mismo para una serie perpetua de generaciones, sino que de vez en cuando, quizá con largos intervalos de tiempo, es indispensable un cruzamiento con otro ejemplar.
Fundándonos en la creencia de que esta es una ley de la naturaleza, creo que podemos comprender varias clases amplias de hechos como los siguientes, que son inexplicables con otro punto de vista. Todo cultivador de híbridos sabe cuan desfavorable para la fecundación de una flor es estar expuesta a la humedad; sin embargo, ¡Qué multitud de flores tienen las anteras y estigmas expuestos a la intemperie! Si fuese indispensable un cruzamiento ocasional, a pesar de que las propias anteras y pistilo de la planta están tan cerca unas de otro que casi aseguran la autofecundación, la necesidad de dejar la más completa libertad de entrada para el polen de otro ejemplar explica que los órganos estén expuestos así. Muchas flores, por otra parte, tienen sus órganos de fructificación estrechamente encerrados, como en la gran familia de las papilionaceas o de las arvejas; pero casi invariablemente estas presentan notables y extrañas adaptaciones en relación con la visita de los insectos. Tan necesarias son las visitas de las abejas para muchas flores papilonaceas, que su fecundidad disminuye mucho si se impide esas visitas.

78
Ahora bien; apenas es posible que los insectos vuelen de flor en flor sin llevar polen de unas a otra, con gran beneficio para la planta. Los insectos obran como una brocha delicada, y para asegurar la fecundidad basta tocar con el mismo pincel las anteras de una flor y luego el estigma de otra; pero no debe suponerse que las abejas producirían así una multitud de híbridos entre distintas especies, porque si el polen propio de la planta y el de la otra especie son colocados sobre el mismo estigma, el primero es tan superior en potencia que invariablemente destruye del todo, como lo ha demostrado Gärtner, la influencia del polen extraño. Cuando los estambres de una flor se tienden súbitamente hacia el pistilo o se mueven lentamente uno tras otro hacia él, este mecanismo parece destinado solamente a asegurar la autofecundación; y no cabe duda de que es útil para ese fin; pero a menudo se necesita la intervención de insectos para que los estambres se tiendan hacia adelanta, como ha demostrado Kölreuter que ocurre en el bérbero; y en este mismo genero, que parece tener un mecanismo especial para la autofecundación, se sabe bien que si se plantan cerca formas o variedades estrechamente vinculadas, apenas se puede producir retoños puros, de tal modo se cruzan naturalmente entre sí. En muchos otros casos, lejos de ser favorecida la autofecundación, hay mecanismos especiales que impiden eficazmente que el estigma reciba polen de su propia flor, como podría demostrarse por las obras de Sprengel y otros, así como con mis propias observaciones: por ejemplo, en la lobelia fulgens hay un mecanismo realmente hermoso y complejo mediante el cual los granitos de polen, infinitamente númerosos, son barridos de las anteras unidas de cada flor antes de que el estigma de esa flor este pronto para recibirlo, y como ésta nunca es visitada, al menos en mi jardín, por los insectos, nunca da semilla, aunque clocando polen de una flor en el estigma de otra yo no produzco cantidad de retoños. Otra especie de lobelia que es visitada por las abejas produce libremente semillas en mi jardín. En muchisimos casos mas, aunque no hay dispositivo mecánico para impedir que el estigma reciba polen de la misma flor, con todo, como ha demostrado Sprengel y más recientemente Hildebrand y otros, y como puedo confirmarlo yo mismo, o bien las anteras revientan antes de que el estigma de la flor este pronto para la fecundación, o bien el estigma esta pronto antes de que este disponible el polen de esa flor, de modo que esas llamadas plantas dicomagas tienen en
realidad sexos separados y deben cruzarse habitualmente. Así pasa con las plantas recíprocamente dimórficas a que aludimos antes. ¡Cuán extraños son estos hechos! ¡Cuán extraño es que el polen y la superficie estigmática de la misma flor, aunque colocados tan próximos como para el propio fin de la autofecundación, sean en tantos casos mutuamente inútiles! ¡Cuán sencillo es explicar esos hechos con la hipótesis de que un cruzamiento ocasional con ejemplar distinto es beneficioso o indispensable! Si se permite echar semilla a unas cuantas variedades de repollo, rábanos, cebollas y algunas otras plantas, cerca unas de otras, gran mayoría de los retoños producidos así resultaran, según he encontrado yo, mestizos. Por ejemplo, saqué 233 retoños de repollo de varias plantas de diversas variedades, que crecían cerca unas de otras, y de estas solamente 78 reprodujeron fielmente su clase, y algunas todavía no del todo. Sin embargo el pistilo de cada flor de repollo esta rodeado no solamente por sus propios seis estambres sino también por los de muchas otras flores de la misma planta, y el polen de cada flor llega fácilmente a su propio estigma sin necesidad de insectos, pues he observado que las plantas cuidadosamente protegidas de los insectos producen el mismo número completo de vainas. Entonces ¿cómo ocurre que tan vasto número de retoños sean mestizados? Debe de ser porque el polen de alguna variedad distinta ha tenido efecto preponderante sobre el polen de la propia flor, y esto es parte de la ley general de que sea beneficioso el intercruzamiento de ejemplares distintos de la misma especie cuando se cruzan especies distintas el caso es inverso, pues el polen propio de una planta casi siempre prepondera sobre el polen extraño; pero sobre este tema volveré en un capitulo futuro.
En el caso de un gran árbol cubierto de flores numerales, puede objetarse que raras veces podría ser llevado el polen de árbol a árbol y a lo sumo podría serlo de una a otra flor del mismo; y que las flores de un mismo árbol solamente en un sentido muy limitado pueden ser consideradas como ejemplares distintos. Creo que esta objeción es valedera, pero que la naturaleza ha obrado ampliamente contra ella al dar a los árboles una fuerte tendencia a producir flores de sexos separados. Cuando los sexos son separados, aunque las flores machos y hembras se produzcan en el mismo árbol, el polen ha de ser llevado regularmente de flor a flor, y esto dará mejores oportunidades y de que ocasionalmente sea llevado el polen de árbol a árbol. Que los

79
árboles pertenecientes a todos los órdenes tienen los sexos separados con más frecuencia que otras plantas, lo he observado en este país; y a pedido mío el Dr. Hooker clasifico los árboles de Nueva Zelandia y el Dr. Asa Gray de los Estados Unidos, y el resultado fue el que yo preveía. En cambio el Dr. Hooker me informa que la regla no se cumple en Australia; pero si la mayoría de los árboles australianos son dicógamos, el mismo resultado se produciría que si dieran flores con sexos separados. He hecho estas pocas observaciones sobre los árboles simplemente para llamar la atención sobre el asunto.
Volviendo por un momento a los animales: varias especies terrestres son hermafroditas, como los moluscos terrestres y lombrices, pero todas ellas se aparean. Hasta ahora no he encontrado un solo animal terrestre que pueda fecundarse a sí mismo. Este hecho notable, que presenta un contraste tan pronunciado con lo que ocurre en las plantas terrestres, es inteligible suponiendo que un cruzamiento ocasional sea indispensable: porque debido a la naturaleza del elemento fecundador no hay medios análogos a la acción de los insectos y del viento entre las plantas, por lo cual pudiera efectuarse un cruzamiento ocasional entre animales terrestres sin la concurrencia de dos ejemplares. Como en el caso de las flores, todavía no he podido, después de consultar con una de las más altas autoridades, es decir el profesor Huxley, descubrir un solo animal hermafrodita con los órganos de reproducción tan perfectamente encerrados que pudiera demostrarse que seria materialmente imposible su acceso desde fuera o la influencia ocasional de un ejemplar distinto. Durante largo tiempo me pareció que los cirrópodos presentan, bajo este punto de vista, un caso muy difícil; pero por una casualidad feliz he podido probar que dos ejemplares, aunque ambos sean hermafroditas que sé autofecundan, se cruzan a veces.
Debe de haber sorprendido a la mayoría de los naturalistas como una extraña anomalía que tanto entre los animales como entre las plantas algunas especies de la misma familia y aun del mismo genero, aunque concuerdan estrechamente entre sí en toda su organización, son hermafroditas y algunas unisexuales. Pero si en realidad todos los hermafroditas se cruzan ocasionalmente, la diferencia entre ellos y las especies unisexuales es muy pequeña en cuanto a esa función. Por esas diversas consideraciones y por muchos hechos especiales que he reunido pero que no puedo exponer aquí parece que
en los animales y plantas un cruzamiento ocasional entre ejemplares distintos es una ley muy general, sino universal, de la naturaleza.
CIRCUNSTANCIAS FAVORABLES PARA LA PRODUCCION DE NUEVAS FORMAS POR
SELECCIÓN NATURAL.
Este es un tema extremadamente intrincado. Una gran variabilidad, término bajo el cual se incluyen siempre las diferencias individuales, será evidentemente favorable. El gran número de ejemplares, al dar mejor oportunidades dentro de cualquier periodo determinado, para la aparición de variaciones provechosas, compensara la menor cantidad de variabilidad en cada ejemplar y creo que constituye un importante factor de éxito. Aunque la naturaleza concede grandes periodos de tiempo para el trabajo de la selección, no concede un periodo indefinido; pues como todos los seres orgánicos se esfuerzan por apoderarse de cada lugar en la economía de la naturaleza, si una especie no es modificada y mejorada en grado correspondiente al de sus competidores, será exterminada. Si no son heredadas variaciones favorables, al menos por algunos de los descendientes, nada puede hacer la selección natural. La tendencia a la reversión podrá contener o impedir a menudo el trabajo; pero si esta tendencia no ha impedido al hombre formar por selección númerosas razas domesticas, ¿por qué prevalecerá contra la selección natural?
En el caso de la selección metódica, un criador elige con algún objetivo definido, y si se permitiera a los ejemplares entrecruzarse libremente, su obra fracasaría en absoluto. Pero cuando muchos hombres, sin tratar de alterar la raza, tienen un tipo de perfección casi igual y todos procuran obtener y criar con los mejores animales, el mejoramiento ha de seguir lentamente pero con seguridad a este proceso de selección, a pesar de que no hay separación de los ejemplares elegidos. Así ocurrirá en el estado de naturaleza, porque dentro de una zona cerrada con algún lugar en la economía de la naturaleza que todavía no este ocupado completamente, todos los ejemplares que varíen en la buena dirección, aunque en grado distinto, tendrán tendencia a prosperar. Pero si la zona fuese grande sus diversos distritos presentarán casi seguramente condiciones distintas de vida, y entonces, si la misma especie sufre modificación en diferentes distritos, las variedades recién formadas se entrecruzarán en los confines de cada una de ellas. Pero

80
veremos en el sexto capitulo que las variedades que habitan distritos intermedios serán suplantadas, a la larga, por alguna de las variedades adyacentes. El intercruzamiento afectara principalmente a esos animales que se unen para cada nacimiento, que viajan mucho y no crían muy rápidamente. De ahí que entre los animales de esa naturaleza, por ejemplo las aves, las variedades, por lo general, se limitaran a países separados, y así he encontrado que ocurre efectivamente. En los organismos hermafroditas que se cruzan solo ocasionalmente y del mismo modo en los animales que se unen para cada nacimiento, pero viajan poco y pueden aumentar con ritmo apresurado, una nueva y mejorada variedad podría formarse en cualquier punto determinado y mantenerse allí en cantidad y luego extenderse, de modo que los ejemplares de la nueva variedad se cruzarían principalmente entre sí. Por este principio, los cultivadores prefieren siempre reservar semillas de grandes masas de plantas, porque así disminuyen las probabilidades de intercruzamiento.
Hasta en los animales que se unen para cada nacimiento y que no se propagan rápidamente no debemos suponer que el libre intercruzamiento eliminaría siempre los efectos de la selección natural; pues puedo presentar un considerable cuerpo de hechos que demuestran que dentro de la misma zona dos variedades del mismo animal pueden permanecer por largo tiempo distintas, debido a que frecuentemente diferentes lugares, crían en temporadas ligeramente distintas, o porque los ejemplares de cada variedad prefieren aparearse entre sí. El intercruzamiento desempeña un papel muy importante en la naturaleza al mantener a los ejemplares de la misma especie o de la misma variedad, genuinos y uniformes en sus caracteres. Evidentemente obrará así de un modo mucho más eficaz con esos animales que se unen para cada nacimiento, pero como ya hemos dicho tenemos razones para creer que un cruzamiento ocasional se produce en todos los animales y plantas. Aún cuando estos ocurran solamente con largos intervalos de tiempo, los hijos producidos así ganaran tanto en vigor y fecundidad sobre los hijos de los que se fecundan a sí mismos durante más tiempo, que tendrán mejores oportunidades de vivir y propagarse; y así a la larga la influencia de los cruzamientos, aun producidos con largos intervalos, será grande. Con respecto a seres orgánicos muy inferiores que no se propagan sexualmente ni aparean y que no pueden intercruzarse, la uniformidad de
caracteres podrá darse a los hijos modificados solamente porque la selección natural ha preservado variaciones similares favorables. El aislamiento tambien constituye un factor importante en la modificación de las especies por selección natural. En una zona cerrada o aislada, siempre que no sea muy grande, las condiciones orgánicas de vida serán por lo general casi uniformes, de modo que la selección natural tendera a modificar todos los ejemplares que varíen de la misma forma. El intercruzamiento con habitantes de los distritos de los alrededores será también impedido así. Moritz Wagner ha publicado recientemente un interesante ensayo sobre este tema, y ha demostrado que el servicio prestado por el aislamiento al impedir los cruzamientos entre variedades recién formadas es probablemente mayor aun de lo que yo había supuesto. Pero por las razones ya dichas no pudo convenir de ningún modo con este naturalista en que la migración y el aislamiento sean elementos necesarios para la formación de especies nuevas. La importancia del aislamiento es igualmente grande para impedir, después de cualquier cambio físico en las condiciones, tales como el clima, elevación de la tierra, etc., la inmigración de organismos mejor adaptados; Y así quedaran abiertos nuevos lugares en la economía natural del distrito para que sean llenados mediante la modificación de los viejos moradores.
Finalmente, el aislamiento dará tiempo para que una nueva variedad sea mejorada lentamente y esto a veces puede tener la mucha importancia. Empero, si una zona aislada es muy pequeña, ya porque esta rodeada por barreras ya porque tiene condiciones físicas muy peculiares, el número total de los habitantes será exiguo, y esto retardara la producción de nuevas especies por selección natural, porque habrán disminuido las probabilidades de que surjan variaciones favorables.
El mero transcurso del tiempo en sí mismo no hace nada en pro ni en contra de la selección natural. Esto lo digo porque se ha afirmado erróneamente que yo supongo que el factor tiempo desempeña un papel de importancia suprema en la modificación de las especies, como si todas las formas de vida sufrieran cambios naturales por alguna ley interna. El transcurso del tiempo solo es importante, y su importancia en este sentido es grande, en cuanto da mejor oportunidad de que surja una variación beneficiosa y sea seleccionada, acumulada y fijada. Igualmente tiende a aumentar la acción directa de las condiciones físicas de vida en relación con la constitución

81
de cada organismo. Si nos volvemos hacia la naturaleza para probar la verdad de estas observaciones y miramos alguna pequeña zona aislada, como una isla oceánica, aunque el número de especies que la habitan sea pequeño, como veremos en nuestro capitulo sobre la distribución geográfica, de esas especies, una gran proporción son endémicas, es decir han sido producidas allí y en ninguna otra parte del mundo. Por eso a primera vista una isla oceánica parece haber sido muy favorable para la producción de nuevas especies. Pero así podemos engañarnos, porque para comprobar que una zona pequeña aislada o una zona grande abierta, como un continente, haya sido más favorable para la producción de nuevas formas orgánicas deberíamos hacer la comparación en tiempos iguales, lo cual no podemos. Aunque el aislamiento es de gran importancia para la producción de nuevas especies, en general me inclino a creer que la amplitud de la zona es todavía más importante, especialmente para la producción de especies que sean capaces de resistir durante largo periodo y de extenderse ampliamente. En toda una zona grande abierta, no solamente habrá mejores probabilidades que surjan variaciones favorables por el gran número d ejemplares de la misma especie que viven allí, sino que las condiciones de vida son mucho más complejas por el gran número de especies que ya existen, y si algunas de esas especies son modificadas y mejoradas, otras tendrán que serlo en grado correspondiente o si no resultaran exterminadas, también, cada nueva forma, tan pronto como haya mejorado mucho, podrá extenderse sobre la zona abierta y continua, y así entrara en competencia con otras muchas formas. Además, grandes zonas, aunque ahora sean continuas, habrán existido no pocas veces debido a las antiguas oscilaciones del nivel, como divididas en muchas partes, de modo que generalmente los buenos efectos del aislamiento, hasta cierto punto, se habrán producido. Finalmente deduzco que aunque zonas pequeñas aisladas sean en algunos sentidos muy favorables para la producción de nuevas especies, el curso de modificaciones habrá sido más rápido en las zonas amplias; y lo que es más importante, las nuevas formas producidas en esas zonas amplias, que ya habían vencido a muchos competidores, serán las que se extenderán más y darán origen al mayor número de nuevas variedades y especies. Así desempeñaran un papel más importante en la historia de los cambios del mundo orgánico. De acuerdo con este punto de vista, quizá podamos entender algunos
hechos a los que aludiremos nuevamente en nuestro capitulo sobre la distribución geográfica; por ejemplo, el hecho de que los productos de ese continente más pequeño que es Australia rindan ahora más pronto que los de la gran zona europeo - asiática. Así también ocurre que los productos continentales han resultado en todas partes muy bien naturalizados en las islas. En una isla pequeña, la carrera será menos severas y habrá menos modificaciones y menos exterminio. Con eso podremos entender como es que la flora de la isla de Madera, según Oswald Heer, se parece en cierto modo a la extinta flora terciaria de Europa. Todas las cuencas de agua dulce sumadas forman una forma pequeña en comparación con la del mar o la tierra, en consecuencia, la competencia entre los productos de agua dulce será menos intensa que en otras partes; nuevas formas habrán sido producidas así más lentamente y las viejas formas serán exterminadas con más lentitud. Y es en el agua dulce donde encontramos siete géneros de los peces ganoideos, restos de un orden en otro tiempo preponderante; y en el agua dulce encontramos algunas de las formas más anómalas que se conocen ahora en el mundo, como el ornitórrinco y los lepidosiros que, como los fósiles, vinculan hasta cierto punto a ordenes que actualmente están muy apartados en la escala de la naturaleza. Estas formas anómalas pueden ser amados fósiles vivos; han resistido hasta hoy porque han habitado una zona limitada y porque han estado sometidas a una competencia menos variada y por consiguiente menos severa. Para resumir, hasta donde lo permite la extremada complicación del tema, las circunstancias favorables o desfavorables para la producción de nuevas especies por selección natural: saco la conclusión de que para los productos terrestres una gran zona continental que haya sufrido mucha oscilaciones de nivel habrá sido la más favorable para la producción de muchas formas nuevas de vida, aptas para soportar por largo tiempo y extenderse ampliamente. Mientras la zona existió como un continente, sus habitantes habrán tenido númerosos ejemplares y clases y habrán estado sometidos a intensa competencia. Cuando después se ha dividido en grandes islas separadas, todavía habrán existido muchos ejemplares de la misma especie en cada isla: el intercruzamiento en los confines de la región propia de cada especie nueva habrá quedado impedido: después de cambios físicos de cualquier clase, la inmigración habrá quedado suprimida, de modo que habrá sido

82
necesario llenar nuevos lugares en la economía de cada isla mediante la modificación de los viejos habitantes, y siempre habrá habido tiempo para que las variedades de cada una resultaran bien modificadas y perfeccionadas. Cuando por una nueva elevación, las islas se convirtieron otra vez en una zona continental, se habrá producido nuevamente intensa competencia: las variedades más favorecidas o modificadas habrán tenido la posibilidad de extenderse; habrá habido mucha extinción de las formas menos mejoradas y los números proporcionales de los diversos habitantes del continente reconstituido habrán cambiado nuevamente, y una vez más habrá existido amplio campo para que la selección natural mejorarse aun a los habitantes y produjese así especies nuevas. Reconozco plenamente que la selección natural obra generalmente con extremada lentitud. Solo puede obrar cuando en la economía natural de un distrito quedan lugares que pueden ser ocupados mejor por la modificación de algunos de sus habitantes. La apertura de tales lugares dependerá a menudo de cambios físicos, que generalmente ocurren muy lentamente, y de que se impida la inmigración de formas mejor adaptadas. Al modificarse algunos de los viejos habitantes, las relaciones mutuas serán perturbadas, y esto creara nuevos lugares prontos para ser llenados por formas mejor adaptadas; pero todo esto ocurrirá con lentitud. Aunque todos los ejemplares de la misma especie difieran levemente unos de otros, a menudo pasara mucho tiempo antes que puedan producirse diferencias de carácter favorable en las diversas partes de la organización. El resultado será con frecuencia muy demorado por el libre intercruzamiento. Muchos dirán que estas diversas causas sobran para neutralizar el poder de la selección natural. Yo no lo creo así. Pero si creo que la selección natural obrara generalmente de un modo muy lento, solo con largos intervalos, y sobre pocos de los habitantes de la misma región. Creo además que estos resultados lentos e intermitentes concuerdan bien con lo que nos dice que la geología sobre la proporción y el modo de que han cambiado los habitantes del mundo. Por lento que sea el proceso de la selección, si el hombre, a pesar de su debilidad, puede hacer mucho con la selección artificial, no veo limite para el grado de cambio, para la belleza y complejidad de las coadaptaciones entre todo los seres orgánicos, unos con otro y con sus condiciones físicas de vida, que puedan haberse efectuado en el largo curso del tiempo por el poder de la naturaleza para
seleccionar, es decir por la supervivencia de los más aptos.
EXTINCION CAUSADA POR LA SELECCIN NATURAL
Este asunto será tratado con más amplitud en nuestro capitulo sobre la geología; pero aquí debemos aludir a él por estar íntimamente relacionado con la selección natural. La selección natural obra solamente mediante la preservación de variaciones que en cierto sentido son ventajosas y por consiguiente se mantienen. Debido a la alta proporción geométrica del incremento de todos los seres orgánicos, cada zona está ya plenamente poblada, y de ahí se sigue que así como las forman favorecidas aumentan en número, generalmente las menos favorecidas disminuyen y se vuelven raras. La rareza, como nos enseña la geología, es la precursora de la extinción. Podemos que cualquier forma que es representada por pocos ejemplares correrá buen riesgo de completa extinción durante las grandes fluctuaciones en la naturaleza de las estaciones, o por un aumento temporal en él número de sus enemigos. Pero podemos ir más lejos, pues a medida que producen nuevas formas, y a menos que admitamos que formas específicas pueden seguir disminuyendo indefinidamente en número, muchas formas viejas habrán de extinguirse. Que él número de formas específicas no ha aumentado indefinidamente nos lo dice claramente la geología, y breve procuraremos demostrar porque él número de especies en todo el mundo no se ha vuelto inconmensurablemente grande.
Hemos visto que las especies que son más númerosas en ejemplares tienen las mejores probabilidades de producir variaciones favorables en un periodo determinado. Tenemos pruebas de esto en los hechos expuestos en el segundo capitulo para demostrar que las especies comunes y definidas, o dominantes, son las que ofrecen a menudo el número mayor de variedades registradas. Pues las especies raras serán modificadas con mucho menos rapidez o menos mejoradas en cualquier periodo; en consecuencia, serán vencidas en la carrera de la vida por los descendientes modificados y mejorados de las especies más comunes. De estas diversas consideraciones se sigue inevitablemente que como el curso del tiempo se forman nuevas especies por selección natural, otras se volverán más y más raras y finalmente se extinguirán. Las formas que se haya en competencia, más estrechas con las

83
que sufren modificación y mejoramiento serán naturalmente las más perjudicadas. Y hemos visto en él capitulo dedicado a la lucha por la existencia que son las formas más estrechamente vinculadas a variedades de la misma especie y especies del mismo genero o géneros relacionados entre sí, las que, por tener casi la misma estructura, constitución y hábitos, entran generalmente en la competencia más severas unas con otras; en consecuencia, cada nueva variedad o especie, durante el proceso de su formación, hará la presión más intensa sobre sus parientes más próximos y tendera a exterminarlos. Vemos el mismo proceso de exterminación entre nuestras producciones domesticas, en la selección por el hombre de las formas mejoradas. Podrían darse muchos ejemplos curiosos para demostrar con cuanta rapidez nuevas razas de vacunos, ovejas y otros animales o variedades de flores toman el lugar de clases inferiores y más antiguas. En Yorkshire se sabe históricamente que los antiguos vacunos negros fueron desplazados por los de cuernos largos y que éstos “fueron desalojados por los de cuernos cortos” (cito estas palabras de un escritor agrícola) “como por pestilencia mortífera”.
DIVERGENCIA DE CARACTERES.
El principio que designado con éste termino es de alta importancia y explica varios hechos significativos. En primer lugar, las variedades, hasta las muy marcadas, aunque tengan cierto carácter de especies –como demuestran las dudas irremediables que en muchos casos se presentan sobre como clasificarlas-, difieren por cierto entre sí mucho menos que la especies verdaderas y distintas. Sin embargo, según mi opinión, las variedades son especies en proceso de formación o como las he llamado, especies incipientes. Entonces ¿cómo es que las diferencias menores entre las variedades resultan transformadas en las grandes diferencias entre las especies? Que esto ocurre habitualmente debemos inferirlo de que la mayoría de las innumerables especies que hay en toda la naturaleza presentan diferencias bien marcadas, mientras que las variedades, los presuntos prototipos y padres de futuras especies bien definidas, presentan diferencias leves y pocos terminantes. La mera casualidad, como pudiéramos llamarla, quizá fuese causa de que una variedad difiera en algún carácter de sus troncos paternos, y los descendientes de esta variedad difieran nuevamente en los mismos caracteres y en mayor grado; pero esto por sí solo nunca explicaría en grado
habitual y tan grande de diferencia como el que se encuentra entre las especies del mismo genero.
Como acostumbro siempre, para este asunto he buscado luz entre nuestras producciones domesticas.
Aquí encontraremos algo análogo. Se admitirá que la producción de razas tan diferentes como los vacunos Shorthorn y Hereford, los caballos de carrera y de tiro, las diversas razas de palomas, etc., nunca habrían podido surgir de la mera acumulación casual de variaciones durante muchas generaciones sucesivas. Por ejemplo, en la practica un criador de palomas se interesa al notar que una de ellas tiene el pico más bien largo; otro por pico corto, y según el principio reconocido de que los aficionados no admiran un termino medio sin que gustan de los extremos, ambos seguirán (como ha ocurrido realmente con las subrazas de la volteadora) eligiendo y haciendo reproducir aves de picos más y más largos, o de picos más y más cortos. Podemos suponer también que en un periodo anterior de la historia, los hombres de una nación o comarca necesitaron caballos más veloces, mientras que los de otras necesitaban caballos más grandes y corpulentos. Las primeras diferencias serian muy leves, pero en el curso del tiempo, debido a la continua selección de caballos más veloces en un caso y de caballos más fuertes en el otro, las diferencias resultarían mayores y serian señaladas como características de dos subrazas distintas. Finalmente después de transcurrir siglos, esas subrazas se convertirían en dos razas bien establecidas y distintas. A medida que aumentan las diferencias, los animales inferiores, con caracteres intermedios, ni muy veloces ni muy fuertes, no habrían sido utilizados para procrear y por lo tanto habrían tendido a desaparecer. Aquí pues, vemos en los productos del hombre la acción de lo que puede llamarse el principio de la divergencia de caracteres, tanto unos de otros, como de los caracteres paternos. Pero puede preguntarse, ¿cómo es posible que e aplique un principio análogo en la naturaleza? Creo que es posible y que se aplica con la mayor efectividad (aunque paso mucho tiempo antes de que yo viese cómo), por la simple circunstancia de que cuanto más diferenciados se vuelvan los descendientes de cualquier especie en estatura, constitución y hábitos, tanto mejor podrán llenar lugares númerosos y muy diversos en la economía de la naturaleza y aumentar en número gracias a ello.

84
Podemos discernir claramente esto en animales con hábitos simples. Tómese el caso de un carnívoro cuadrúpedo del cual él número que puede soportar algún país haya llegado hace tiempo al termino medio completo. Si se deja obrar su poder natural para aumentar (no sufriendo el país ningún cambio en las condiciones) solamente podrá conseguirlo si sus descendientes modificados toman lugares hasta entonces ocupados por otros animales; algunos de ellos, por ejemplo, pudiendo alimentarse de nuevas clases de presas, vivas o muertas; algunos habitando nuevas localidades, trepando a los árboles, frecuentando el agua, y quizá unos volviéndose menos carnívoros. Cuanto más diferenciados en hábitos y estructura se vuelvan los descendientes de nuestro carnívoro, más lugares podrán ocupar. Lo que se aplica a un animal se aplicara a todos los animales a través del tiempo, es decir, si varían, pues de otro modo la selección natural no podrá hacer nada. Así ocurrirá con las plantas. Se ha demostrado experimentalmente que si un trozo de terreno es sembrado de una especie de hierba y otro trozo semejante lo es con varios géneros distintos de hierba, podrá criarse mayor número de plantas y cosecharse más cantidad de heno en el segundo caso que no en el primero. Lo mismo se ha probado cuando una variedad y unas cuantas variedades mezcladas de trigo se plantan en espacios iguales de tierra. Por eso, si alguna especie de hierba hubiera de seguir variando y las variedades que difiriesen entre sí del mismo modo, aunque levemente, como ocurre entre las distintas especies y géneros de hierbas, fuesen continuamente seleccionadas, un número mayor de ejemplares de esta especie, inclusive sus descendientes modificados, conseguiría poder vivir en el mismo pedazo de tierra. Y sabemos que cada especie y cada variedad de hierba siembra anualmente semillas innumerables, y puede decirse que así se esfuerza en lo posible para aumentar. En consecuencia, en el curso de muchos miles de generaciones, las variedades más distintas de cualquier especie de hierba tendrían mejores posibilidades de prosperar y aumentar en número, suplantando así a las variedades menos distintas. Y las variedades, una vez muy diferenciadas entre si, toman la categoría de especies. La verdad del principio de que la mayor cantidad de vida puede ser sostenida por una gran diversificación de estructura se ven en muchas circunstancias naturales. En una zona extremadamente pequeña sobre todo si sé esta libremente abierta a inmigración, y donde la rivalidad
entre ejemplar y ejemplar ha de ser muy intensa, siempre encontramos mucha diversidad en sus habitantes. Por ejemplo, he comprobado que un pedazo de césped de tres pies por cuatro, que había estado expuesto durante muchos años exactamente a las mismas condiciones, sostenía veinte especies de plantas, y éstas pertenecían a dieciocho géneros y ocho ordenes, lo cual demuestra cuanto diferían esas plantas entre sí. Lo mismo pasa con las plantas e insectos que viven en islitas uniformes o en pequeños estanques de agua dulce. Los agricultores comprueban que pueden obtener mayor cantidad de alimento mediante la rotación de plantas pertenecientes a los ordenes diferentes; la naturaleza efectúa lo que podría llamarse una rotación simultanea. La mayoría de los animales y plantas que viven muy cerca en torno a una pequeña sección de tierra podrían vivir de ella (suponiendo que su naturaleza no fuese peculiar en algún sentido) y quizá se esfuerzan todo lo posible para vivir allí; pero se ha visto que cuando entran en competencia muy estrecha, las ventajas d la diversificación de la estructura, con las diferencias correspondientes de hábitos y constitución, determinan que los habitantes que así se estorban unos a otros pertenecen, por regla general, a lo que llamamos ordenes y géneros distintos. El mismo principio se ve en la aclimatación de las plantas a tierras extranjeras, por obra del hombre. Podría haberse esperado que las plantas que consiguiesen aclimatarse en algún país serian las más estrechamente relacionadas con las indígenas, pues a estas se les considera por lo general como especialmente creadas y adaptadas para su propio país. Quizá también podría haberse esperado que las plantas aclimatadas perteneciesen a pocos grupos más especialmente adaptados a ciertas regiones de su nueva patria. Pero el caso es muy diferente, y Alfonso de Candolle, en su grande y admirable obra, ha señalado con acierto que las flores ganan por aclimatación, y en comparación con el número de los géneros y especies nativos, mucho más en nuevos géneros que en nuevas especies. Para dar un solo ejemplo: en la ultima edición del Manual de la flora del norte de los Estados Unidos, del Dr. Asa Gray, se enumeran 260 plantas aclimatadas que pertenecen a 162 géneros. Vemos así que esas plantas aclimatadas son de naturaleza muy diferenciada. Se diferencian además, en gran parte, de las indígenas, pues de los 162 géneros aclimatados, no menos de 100 géneros no son allí autóctonos, y de este modo una gran adición proporcional se ha

85
hecho a los géneros que viven ahora en los Estados Unidos.
Considerando la naturaleza de las plantas o animales que en algún país han luchado con esto contra las indígenas y se han aclimatado, podemos obtener alguna idea rudimentaria de la firma en que los seres nativos tendrían que modificarse para ganar alguna ventaja sobre sus compatriotas, y por lo menos podemos inferir que les seria provechosa una diversificación de su estructura que llegase hasta constituir nuevas diferencias genéricas. La ventaja de la diversificación de estructura en los habitantes de la misma región es, en verdad, la misma que la de división fisiológica del trabajo entre los órganos del mismo cuerpo individual, asunto también aclarado por Milne Edwards. Ningún filosofo duda de que un estomago adaptado a digerir solamente materia vegetal o solamente carne, saca más alimento de esas sustancias. Del mismo modo en la economía general de un país, cuanto más y mejor estén diversificados los animales y plantas para diferentes hábitos de vida, tanto mayor será el número de ejemplares capaces de sostenerse allí. Un conjunto de animales cuya organización sea poco diversa no podrá competir con un conjunto mejor diversificado en su estructura. Por ejemplo, puede dudarse de que los marsupiales australianos, que están divididos en grupos que difieren poco entre sí y representan débilmente, como han observado Mr. Waterhouse y oros, a nuestros mamíferos carnívoros, rumiantes y roedores, pudieran competir con estos órdenes bien desarrollados. Entre los mamíferos australianos vemos el proceso de diversificación en una fase temprana e incompleta de desarrollo.
PROBABLES EFECTOS DE LA ACCION DE LA SELECCIÓN NATURAL, POR LA DIVERGENCIA
DE CARACTERES Y LA EXTINCION, SOBRE LOS DESCENDIENTES DE UN ANTEPASADO
COMÚN
Después de las consideraciones anteriores, que hemos abreviado mucho, podemos suponer que los descendientes modificados de una especie prosperan mucho más a medida que se diversifiquen en su estructura y puedan así arraigar en los lugares ocupados por otros seres. Ahora veamos como tiende a obrar este principio del beneficio que se obtiene de la divergencia de los caracteres, combinados con los principios de selección y extinción naturales.
El diagrama que insertamos nos ayudara a comprender este problema algo complicado. Supongamos que las letras de la A hasta la L representan las especies de un genero grande en su propio país; y que esas especies se parecen unas a otras en grado desigual, como ocurre tan a menudo en la naturaleza y como lo indicamos en el diagrama colocando las letras a distancias desiguales. He dicho un genero grande porque, como vimos en el segundo capitulo, por termino medio varían más las especies de los géneros mayores que las de los géneros menores, y las especies que varían en los géneros grandes presentan mayor número de variedades. También hemos visto que las especies más comunes y más ampliamente difundidas varían más que las especies raras y restringidas. Sea (A) una especie común, ampliamente difundida, y muy variada, perteneciente a un genero grande en su propio país. Las líneas punteadas y ramificadas de largo desigual que parten de (A) pueden representar a su variada descendencia. Suponemos que las variaciones son extremadamente leves, pero del carácter más vario, y que no aparecen simultáneamente, sino a menudo después de largos intervalos: tampoco suponemos que persistan por periodos iguales. Solamente las variaciones que de algún modo son provechosas serán preservadas o seleccionadas naturalmente. Y aquí entra la importancia del principio que es beneficiosa la divergencia de caracteres, porque él conducirá generalmente a que sean preservadas y acumuladas por selección natural las más diferentes o divergentes variaciones (representadas por las líneas punteadas exteriores). Cuando una línea de puntos llega a una de las líneas horizontales y es marcada allí por una letra numerada, se supone que se ha acumulado suficiente cantidad de variación para constituir una variedad muy bien definida, como la que seria digna de registrarse en una obra sistemática. Los intervalos entre las líneas horizontales del diagrama pueden representar cada uno mil o más generaciones. Después de mil generaciones se supone que la especie (A) ha producido dos variedades bien marcadas, a saber: (a1) y (m1). Estas dos variedades seguirán generalmente expuestas a las mismas condiciones que hicieron variables a sus padres y la tendencia a variar es por si misma hereditaria; en consecuencia, tenderán igualmente a variar y por lo común del mismo modo que sus padres. Además, esas dos variedades, siendo solamente formas sólo modificadas de un modo leve, tenderán a heredar las ventajas que hicieron a su padre

86
(A) más númeroso que la mayoría de los otros habitantes del mismo país; participaran también e esas ventajas generales que hicieron que el genero al que pertenecía la especie padre fuese un genero grande en su propio país. Y todas esas circunstancias son favorables para la producción de nuevas variedades. Entonces, si esas dos variedades son variables, las más divergentes de sus variaciones serán generalmente preservadas en las próximas mil generaciones. Y después de este intervalo se supone en el diagrama que la variedad (a1) ha producido a la variedad (a2), la cual, debido al principio de la divergencia, diferirá de (A) más que la variedad (a1). Se supone que la variedad (m1) ha producido dos variedades: (m2) y (s2), que difieren entre si y difieren todavía más de su padre común (A). Podemos continuar este proceso, por pasos semejantes, durante cualquier lapso de tiempo; algunas de las variedades, después de cada mil generaciones, producirán solamente una variedad, pero de condición más y más modificada, produciendo algunas dos o tres variedades, y otras no produciendo ninguna. Así las variedades o descendientes modificados del padre común (A) seguirán por lo general aumentando en número y divergiendo en caracteres. En el diagrama ese proceso esta representado hasta la generación Nº 10.000, y en forma condensada y simplificada, hasta la generación Nº 14.000.
Pero debo observar aquí que no supongo que el proceso vaya siempre de un modo tan regular como lo he representado en el diagrama, aunque él mismo es un poco irregular, ni tampoco que siga sin solución de continuidad; mucho más probable es que cada firma permanezca por largos periodos inalterados y luego sufra nuevamente una modificación. Tampoco supongo que sean preservadas siempre las variedades más diversas: a menudo podrá resistir una forma intermedia por largo tiempo, y podrá producir o no producir más de un descendiente modificado, pues la selección natural obra siempre según la naturaleza de los lugares que todavía están desocupados o imperfectamente ocupados por otros habitantes, y esto dependerá de relaciones infinitamente complejas. Pero por regla general, cuanto más diversificados en estructura puedan llegar a ser descendientes de una especie, más lugares podrán ocupar y más aumentara su progenie modificada. En nuestro diagrama, la línea de sucesión esta interrumpida a intervalos regulares por pequeñas letras numeradas que señalan las formas sucesivas que se han vuelto
suficientemente distintas para ser registradas como variedades. Pero esas interrupciones son imaginarias y podrían haber sido insertadas en cualquier parte, después de intervalos suficientes para permitir la acumulación de un grado considerable de variación divergente.
Como todos los descendientes modificados de una especie común y ampliamente difundida, perteneciente a un genero grande, tenderán a participar de las mismas ventajas que dieron éxito al padre en la vida, generalmente seguirán multiplicándose y divergiendo en caracteres: esto es representado en el diagrama por varias ramas divergentes que parten de (A). La descendencia modificada de las ramas posteriores y más mejoradas de las líneas genealógicas tomaran a menudo, es probable, el lugar de ramas anteriores y menos mejoradas, destruyéndolas: esto esta representado en el diagrama por algunas ramas inferiores que no llegan a las líneas horizontales superiores. En algunos casos, no cabe duda de que el proceso de modificación se limitara a una sola línea genealógica y no aumentara quizá el número de descendientes modificados: aunque así habrá aumentado tal vez el grado de modificación. Este caso seria representado en el diagrama si todas las líneas que parten de (A) fueran suprimidas, exceptuando la que va de (a1) a (a10). Del mismo modo, el caballo ingles de carrera y el ventor inglés ha ido lentamente divergiendo en carácter, según parece, de sus troncos originales, sin que hayan dado origen a nuevas ramas o razas.
Después de 10.000 generaciones se supone que la especie (A) a producido tres formas: (a10), (f10) y (m10), las que por haber divergido en caracteres durante las generaciones sucesivas vendrán a diferir ampliamente, pero quizá en grado desigual, unas de otras y de su padre común. Si suponemos que el grado de cambio entre cada línea horizontal es excesivamente pequeño, estas tres forma podrán ser todavía solo variedades bien marcadas; pero nos basta suponer que los pasos de ese proceso de modificación sean más númerosos o amplios para convertir esas tres formas en especies dudosas o por lo menos mal definidas. Así el diagrama ilustra los pasos por los cuales las diferencias pequeñas que distinguen a las variedades aumentan, transformándose en las diferencias mayores que distinguen a las especies. Siguiendo el mismo proceso durante mayor número de generaciones (como muestra el diagrama en forma abreviada y simplificada) obtenemos ocho especies

87
marcadas por letras que van de (a14) a (m14), todas descendientes de (A). Así creo yo que se multiplican las especies y se forman los géneros. En un genero grande probable que más de una especie varíe. En el diagrama he supuesto que una segunda especie (I) haya producido, por pasos análogos, después de diez mil generaciones, dos variedades bien marcadas (w10 y z10) o dos especies, según el grado de cambio que suponemos representado entre las líneas horizontales. Después de catorce mil generaciones, seis nuevas especies, marcadas por las letras que van de (n14 a z14), han sido producidas según nuestra suposición. En cualquier genero, las especies que ya son muy diferentes unas de otras en sus caracteres tenderán generalmente a producir el mayor número de descendientes modificados, pues éstos tendrán la mejor oportunidad de apoderarse de nuevos lugares, ampliamente diversos, en la economía de la naturaleza: por eso en el diagrama he elegido la especie extrema (A) y la casi extrema (I) como las que han variado ampliamente y dado origen a nuevas variedades y especies. Las otras nueve especies. Las otras nueve especies (señaladas con letras mayúsculas) de nuestro genero original, pueden seguir durante largos pero desiguales periodos dando descendientes inalterados, y así lo muestra el diagrama con la línea de puntos desigualmente prolongadas hacia arriba.
Pero durante el proceso de modificación representado en el diagrama, otro de nuestros principios, el de la extinción, habrá desempeñado un papel importante. Como en cada país plenamente poblado la selección natural obra necesariamente por el hecho de que la forma seleccionada tiene alguna ventaja en la lucha por la vida sobre otras formas, habrá en los descendientes mejorados de una especie una tendencia constante a suplantar y exterminar en cada etapa de descendencia a sus predecesores y su progenitor original. Por eso todas las formas intermedias entre los estados anteriores y posteriores, es decir entre los estados menos mejorados de la misma especie, así como la propia especie madre, tenderán a extinguirse. Ello ocurrirá probablemente en muchas líneas posteriores y mejoradas. Empero si la descendencia modificada de una especie entra en algún país distinto o se adapta rápidamente a alguna región completamente nueva en la que no entren en competencia descendientes y progenitores, unos y otros podrán seguir existiendo. Entonces, si se supone que nuestro diagrama representa un grado considerable de modificación, la especie
(A) y todas las variedades anteriores se habrán extinguido así, siendo reemplazadas por ocho especies nuevas (a14 hasta m14), i la especie (I) será reemplazada por seis especies nuevas (n14 a z14).
Pero podemos ir aun más lejos. Se había supuesto que las especies originales de nuestro género se parecían unas a otras en grado desigual, como ocurre generalmente en la naturaleza, estando la especie (A) relacionada más de cerca con B, C, y D que las otras especies; Y la especie (I) más cerca de G, H, K, y L que las otras. Habíamos supuesto también que estas dos especies, (A) e (I) eran muy comunes estaban muy difundidas, de modo que originalmente habrían de tener alguna ventaja sobre la mayoría de las otras especies del genero. Probablemente sus descendientes modificados, que son catorce en la generación Nº 14.000, habrán heredado algunas de esas ventajas; así también han cambiado y mejorado de manera diversa en cada fase de la genealogía, de modo que se han adaptado a muchos lugares relacionados en la economía natural de su país. Parece por lo tanto extremadamente probable que hayan quitado los lugares – y por consiguiente exterminado- no solamente a sus padres (A) e (I) sino también algunas de las especies originales que estaban más estrechamente relacionadas con sus padres. En consecuencia, muy pocas de las especies originales habrán prolongado su descendencia hasta la generación Nº 14.000. podemos suponer que solamente una, (F), de las dos especies (E y F) que estaban menos estrechamente relacionadas con las otras nueve especies originales, ha hecho llegar sus descendientes hasta esta avanza etapa genealógica. Las nuevas especies de nuestro diagrama, descendidas de las once especies originales, serán ahora quince. Debido a la tendencia divergente de la selección natural, el grado extremo de diferencia de caracteres entre las especies (a14 y z14) será mucho mayor que entre las más distintas de las once especies originales. Además las nuevas especies estarán relacionadas entre sí de un modo muy diferente. De los ocho descendientes de (A), los tres señalados con (a14, q14 y p14) estarán estrechamente relacionados por haberse desprendido recientemente de (a10, b14 y f14) por haber divergido en un periodo anterior de (a5), serán en cierto grado distintas de las otras tres especies nombradas primero; Y por último, (o14, e14 y m14) estarán más estrechamente relacionadas entre sí, pero por haber divergido al empezar el proceso de modificación, serán muy

88
diferentes de las otras cinco especies y podrán constituí un subgénero o genero distinto. Los seis descendientes de (I) formaran dos subgéneros o géneros. Pero como la especie original (I) difería ampliamente de (A), estando casi en el limite extremo del genero original, los seis descendientes de (I) diferían considerablemente, debido a la sola herencia, de los ocho descendientes de (A), además, se supone que los dos grupos han seguido divergiendo en direcciones distintas. También las especies intermediarias (esto es muy importante) que vinculan a las especies originales (A) e (I) se habrán extinguido todas, menos (F) y no habrán dejado descendientes.
De ahí que las seis nuevas especies descendidas de (I) y los ocho descendientes de (A) tendrán que ser clasificadas como géneros muy distintos y aun como subfamilias distintas. Así ocurre, en mi opinión, que dos o más géneros son producidos por una descendencia, con modificaciones, desde dos o más especies del mismo genero. Y se supone que las dos o más especies madres descendían de alguna especie de algún genero anterior. En nuestro diagrama esto es indicado por las líneas de guiones que están debajo de las letras mayúsculas, convergiendo en subramas hacia abajo en dirección a un punto solo; este punto representa a una especie, el presunto progenitor de nuestros varios subgéneros y géneros nuevos. Vale la pena reflexionar por un momento sobre el carácter de la nueva especie, (F14), que se supone que no ha divergido mucho en caracteres sino que ha retenido la forma de (F), inalterada o alterada solamente en grado leve. En este caso, sus afinidades con las otras catorce especies nuevas serán de carácter curioso e indirecto. Habiendo descendido de una forma que estaba entre las especies madre(A) a (I), que ahora suponemos extintas y desconocidas, tendrá en cierto modo carácter intermediario ente los dos grupos descendidos de esas dos especies. Pero como esos dos grupos han seguido divergiendo en caracteres del tipo de sus padres la nueva especie (F14) no será indirectamente intermedia entre ellas, sino más bien entre tipos de los dos grupos, y todo naturalista puede recordar casos semejantes. Hasta ahora hemos supuesto que cada línea horizontal del diagrama representa mil generaciones, pero cada una podría representar un millón y más; también puede representar una sección de los estratos sucesivos de la corteza terrestre, inclusive los restos extintos. Cuando lleguemos a nuestro capitulo sobre geología, tendremos que
referirnos nuevamente a esta tema, y creo que entonces veremos que el diagrama arroja luz sobre las afinidades de seres extintos, que aunque generalmente pertenecen a los mismos ordenes, familias o géneros que los que viven ahora, son a menudo, en algún grado, de carácter intermedio entre grupos existentes, y podemos entender este hecho por que las especies extinguidas han vivido en varias épocas remotas, cuando las líneas ramificadas de descendencia no habían llegado a diferir tanto. No veo razones para limitar el proceso de modificación, como lo he explicado ahora, a la formación de géneros solamente. Si suponemos en el diagrama que es grande el grado de cambio representado por cada grupo sucesivo de líneas punteadas divergentes, las formas marcadas desde (a14 hasta m14) constituirán tres géneros muy distintos. También tendremos dos géneros muy distintos que habrán descendido de (I) y que diferían mucho de los descendientes de (A). Esos dos grupos de géneros formaran así dos familias u ordenes distintos, según el grado de divergencia que se supone representado en el diagrama. Y las dos nuevas familias, u ordenes, descienden de dos especies del genero primitivo, y se suponen que estas descienden de una forma aun más antigua y desconocida. Hemos visto que en cada país son las especies que pertenecen a los géneros mayores las que más a menudo presentan variedades o especies incipientes. En verdad, esto era de esperar, porque como la selección natural obra gracias a que alguna forma tiene cierta ventaja sobre otras formas en la lucha por la existencia, obrara principalmente sobre las que ya tienen alguna ventaja, y la amplitud de cada grupo demuestra que sus especies han heredado de algún antepasado común cierta ventaja común. Por eso la lucha por la producción de descendientes modificados se librara principalmente entre los grupos mayores, todos los cuales procuran aumentar en número. Un grupo grande vencerá lentamente a otro grupo grande, disminuirá su número y reducirá así sus probabilidades de variar y mejorar más. Dentro del mismo grupo amplio, los grupos posteriores y más perfeccionados, por haberse ramificado y capturado nuevos lugares en la economía de la naturaleza, tenderán constantemente a suplantar y destruir a los subgrupos anteriores y menos perfeccionados. Finalmente, los grupos y subgrupos pequeños e incompletos desaparecerán. Mirando hacia el futuro, podemos predecir que los grupos de seres orgánicos que ahora son amplios y triunfantes y que son menos incompletos, es decir que

89
hasta ahora han sufrido menos extinción, seguirán incrementando por un largo periodo. Pero nadie puede predecir que grupos prevalecerán al fin, porque sabemos que muchos grupos que antes se habían desarrollado ampliamente se han extinguido. Mirando todavía más lejos en el futuro, podemos predecir que debido al aumento continuo y firme de los grupos mayores, multitud de grupos menores se extinguirán totalmente y no dejaran descendientes modificados, y que por lo tanto, de las especies que viven en cualquier periodo serán muy pocas las que hagan llegar descendientes a un porvenir remoto. Tendré que volver sobre este tema en el capitulo relativo a la clasificación, pero puedo añadir ahora que, en mi opinión, muy pocas de las especies más antiguas habrán perpetuado su descendencia hasta el día de hoy, y que como todos los descendientes de una misma especie constituyen una clase, podemos entender como existen tan pocas clases en cada división principal de los reinos animal y vegetal. Aunque pocas de las especies más antiguas han dejado descendientes modificados. Con todo, en periodos geológicos remotos, la tierra puede haber estado casi aun copiosamente poblada de especies de muchos géneros, familias y ordenes como en la actualidad.
SOBRE EL GRADO EN QUE TIENDE A AVANZAR LA ORGANIZACIÓN
La selección natural obra exclusivamente mediante la preservación y acumulación de variaciones que son beneficiosas bajo las condiciones orgánica e inorgánicas a que esta expuesta cada criatura en todos los periodos de vida. El resultado final es que cada criatura tiende a mejorar más y más con respecto a sus condiciones. Este mejoramiento conduce inevitablemente al progreso gradual en la organización del mayor número de seres vivos en todo el mundo. Pero aquí entramos en un tema muy intricado, porque los naturalistas no han definido todavía a satisfacción unos de otros que se entiende por progreso en la organización. Entre los vertebrados, el grado de inteligencia y cierta proximidad con el hombre en cuanto a la estructura entran naturalmente en juego. Podría pensarse que el grado de cambio por que pasan las diversas partes y órganos en su desarrollo desde el embrión a la adultez bastaría como punto de comparación; pero hay casos, como en ciertos crustáceos parasitarios, en que varias partes de la estructura se vuelven menos perfectas, de modo que el animal adulto no puede ser
calificado de más elevado que su larva. El patrón adoptado por von Baer parece ser el más aplicable y el mejor, a saber; el grado de diferenciación de las partes del mismo ser orgánico (en estado adulto, me inclinaría a añadir yo) y su especialización para funciones diferentes; o, como lo expresaría Milne Edwards, lo completo de la división del trabajo fisiológico. Pero veremos cuan obscuro es este problema si consideramos, por ejemplo los peces, entre los cuales algunos naturalistas clasifican como los más elevados a los que, como el tiburón, se aproximan más a los anfibios; mientras que otros naturalistas clasifican a los peces óseos o teleósteos comunes como los superiores porque son peces en un sentido más estricto y difieren más de las otras clases de vertebrados. Vemos todavía más claramente la obscuridad del problema volviendo a las plantas, en las cuales queda excluida, naturalmente, la cuestión de la capacidad intelectual; y algunos botánicos clasifican como superiores a las plantas que tienen todos los órganos, como sépalos, pétalos, estambres y pistilos plenamente desarrollados en cada flor; al paso que otros, probablemente con más verdad, consideran superiores a las plantas que tienen sus diversos órganos muy modificados y reducidos en número. Si tomamos como tipo de alta organización el alcance de la diferenciación y especialización de los diversos órganos de cada ser adulto (lo cual incluirá el desarrollo del cerebro para los fines intelectuales), la selección natural nos lleva claramente hacia ese tipo pues todos los fisiológicos reconocen que la especialización de los órganos es una ventaja para cada ser porque así cumplen mejor sus funciones; y por eso la acumulación de variaciones tendientes a la especialización esta dentro del objetivo de la selección natural. Por otra parte, si recordamos que todos los seres orgánicos procuran aumentar en alta proporción y apoderarse de cada lugar desocupado no muy bien ocupado de la economía natural, podemos comprender que la selección natural adapte gradualmente a un ser a una situación en que varios órganos serian superfluos o inútiles; en tales casos habrá una represión en la escala de lo organizado. En nuestro capitulo sobre la sucesión geológica estudiaremos con más detención si la organización en general ha procesado realmente desde lo más remotos periodos geológicos hasta la actualidad. Pero puede objetarse: si todos los seres orgánicos tienden a elevarse en la escala ¿Cómo es que existen aun en todo el mundo multitudes de formas muy inferiores; Y como es que en cada

90
clase amplia, algunas formas están mucho más avanzadas que otras? ¿Por qué las formas más desarrolladas no han suplantado y exterminado en todas partes a las inferiores? Lamarck, que creía en una tendencia innata e inevitable hacia el perfeccionamiento en todos los seres orgánicos parece haber sentido esa dificultad, tan vivamente que fue llevado a suponer que continuamente se producen formas nuevas y simples por generación espontanea. La ciencia no ha probado aun la verdad de esta opinión, sea lo que fuere lo que revele el porvenir. Según nuestra teoría, la supervivencia de organismos inferiores no ofrece dificultad, pues la selección o la supervivencia de los más aptos no implica necesariamente desarrollo progresivo: solo utiliza las variaciones que surgen y son beneficiosos para cada criatura en sus complejas relaciones de vida. Y puede preguntarse que ventaja seria para un infusorio, para un gusano intestinal, y aun para una lombriz, estar elevadamente organizados. Si no fuese una ventaja, tales formas serian dejadas por la selección natural sin mejoramiento o con poco mejoramiento, y quizá permaneciesen durante edades indefinidas en su actual condición inferior. Y la geología nos dice que algunas de las formas inferiores, como los infusorios y los rizópodos, han quedado durante periodos desmesurados casi en su estado actual. Pero suponer que la mayoría de las formas inferiores que existan ahora no han progresado en nada desde los primeros albores de la vida, seria imprudente, pues todo naturalista que haya disecado algunos de los seres ahora colocados muy bajo en la escala, ha de ver quedado sorprendido por su organización acabada y realmente maravillosa. Casi las mismas observaciones son aplicables cuando consideramos los diversos grados de organización dentro del mismo grupo grande; por ejemplo, en los vertebrados, la coexistencia de mamíferos y peces; entre los mamíferos, la coexistencia del hombre y el ornitorrinco; entre los peces; la coexistencia del tiburón y del pez lanza(anfioxo), pez este último que por la extremada sencillez de su estructura se aproxima a las clases invertebradas. Pero mamíferos y peces difícilmente entraran en competencia unos con otros, el progreso de toda la clase de los mamíferos o de ciertos miembros de esa clase, hasta el más alto grado, no los llevaría a ocupar el lugar de los peces. Los fisiólogos creen que el cerebro debe estar irrigado por sangre caliente para ser activo, y para esto se necesita respiración aérea, de modo que los mamíferos de sangre caliente, cuando habitan
en el agua se hallan en desventaja porque tienen que salir continuamente a la superficie para respirar. Entre los peces, los miembros de la familia de los tiburones no tenderían a suplantar al pez lanza; pues éste, como me dice Fritz Müller, tiene como único compañero y competidor en las estériles costas arenosas del sur de Brasil un anélido anómalo. Los tres órdenes inferiores d mamíferos, a saber: marsupiales, desdentados y roedores, coexisten en la América del Sur, y en las mismas regiones, con númerosos monos y probablemente se molestan poco unos a otros. Aunque la organización, en general, puede haber avanzado y estar avanzando aún en todo el mundo, la escala siempre ofrecerá muchos grados de perfección, pues el alto desarrollo de algunas clases enteras o de ciertos miembros de cada clase no conduce necesariamente a la extinción de esos grupos con los cuales no entran en competencia estrecha. En ciertos casos, como veremos después, formas inferiormente organizadas parecen haberse conservado hasta la actualidad por habitar regiones confinadas o peculiares, donde han estado sometidas a competencia menos severa y donde su escaso número ha retardado la oportunidad de que sugieran variaciones favorables.
Finalmente, creo que muchas formas inferiores organizadas existen aun en todo el mundo por varias causas. En algunos casos variaciones o diferencias individuales de naturaleza favorable pueden no haber surgido nunca para que la selección natural operase y las acumulase. Probablemente en ningún caso ha bastado el tiempo para que se llegase al último grado posible de desarrollo. En unos cuantos casos, ha habido lo que podemos llamar regresión. Pero la causa principal consiste en el hecho de que bajo condiciones de vida muy simples, una alta organización no serviría de nada, y posiblemente hasta resultaría desventajosa por ser más delicada y más expuesta a sufrir perturbaciones y daños. Mirando hacia el primer albor de vida, cuando todos los seres orgánicos, según podemos creer, presentaban la misma estructura simple, se ha preguntado como han surgido los primeros caos en el progreso o diferenciación de las partes. Probablemente Mr. Herbert Spencer contestaría que tan pronto como el simple organismo unicelular, por la división o por el crecimiento, llego ha estar formado por varias células, o adherido a alguna superficie de sostén, entraría en juego su ley de que “ las unidades homologas de cualquier orden se diferencian proporcionalmente a medida que se diferencian sus relaciones con las fuerzas

91
incidentes”. Pero como no tenemos hechos que nos guíen, resulta casi inútil especular sobre el tema. Es un error, empero, suponer que no habría lucha por la existencia, y en consecuencia selección natural hasta que hubieran nacido muchas formas: las variaciones de una sola especie que habiten una región aislada podrían ser beneficiosas y así toda la masa de ejemplares resultaría modificada, o bien surgirían dos formas distintas. Pero como observe al final de la introducción, nadie debe sorprenderse de que mucho quede sin explicar aun sobre el origen de las especies si tenemos en cuenta nuestra profunda ignorancia de las relaciones mutuas entre los habitantes actuales del mundo, y más aun entre los de edades anteriores.
CONVERGENCIA DE CARACTERES
Mr. H: C. Watson opina que he exagerado la importancia de la divergencia de caracteres (en la que él, empero, parece creer) y que la convergencia, como puede llamarse, ha desempeñado también su parte. Si dos especies pertenecientes a dos géneros distintos, aunque relacionados, hubieran producido gran número de formas nuevas y divergentes, es concebible que las mismas pudieran aproximarse tanto que todas habrían de ser clasificadas en el mismo genero, y así los descendientes de dos géneros distintos se convertirían en uno. Pero en la mayoría de los casos seria muy imprudente atribuir a la convergencia una similitud estrecha y general de estructura en los descendientes modificados de formas ampliamente diferentes. La forma de un cristal esta determinada solamente por las fuerzas moleculares y no es sorprendente que substancias desemejantes asuman a menudo la misma forma; pero en cuanto a los seres orgánicos debemos recordar que la forma de cada uno depende de una multitud de relaciones infinitamente complejas, a saber: de las variaciones que han surgido y que se deben a causas demasiado intrincadas para ser seguidas, de la naturaleza de las variaciones que se han preservado o seleccionado, y ésta depende a su vez de las condiciones físicas circundantes, y en mayor grado aun de los organismos que rodean a cada ser y con los que este ha entrado en competencia; por último, de la herencia (de por sí un elemento fluctuante) procedente de progenitores innumerables, la forma de todos los cuales ha sido determinadas por relaciones igualmente complejas. No es creíble que los descendientes de dos organismos que originalmente habían diferido
pronunciadamente convergieran siempre después de un modo tan próximo que los condujese a una casi identidad en toda su organización. Si esto hubiese ocurrido, encontraríamos que una misma forma, independientemente de las vinculaciones genéticas, reaparecería de formaciones geológicas ampliamente separadas, y el saldo de las pruebas reunidas es contrario a tal suposición. Mr. Watson a objetado también que la acción continua de la selección natural, junto con la divergencia de carácter, tendería a crear un número indefinido de formas específicas. En cuanto a las formas meramente inorgánicas, parece verisímil que un número suficiente de especies se adapten pronto a todas las diferencias considerables de calor, humedad, etc., Pero reconozco plenamente que las relaciones mutua de los seres orgánicos son más importantes, y así como él número de especies de cualquier país va aumentando, las condiciones orgánicas de vida deben volverse más complejas más y más complejas. En consecuencia, a primera vista parece que no habrá limite al grado de diversificación favorable de estructura y, por consiguiente, tampoco para el número de especies que podrían ser producidas. No sabemos que ni siquiera la zona más prolífica esté plenamente poblada de formas específicas: en el Cabo de Buena Esperanza y en Australia, que sostienen tan asombroso número de especies, muchas plantas europeas se han aclimatado. Pero la geología nos da muestra que desde una época temprana del periodo terciario, el número de especies de moluscos no ha aumentado mucho, o nada, ni tampoco el número de mamíferos desde la mitad de dicho periodo. Entonces, ¿qué es lo que obstruye el crecimiento indefinido del número de especies? La cantidad de vida (no me refiero al número de formas específicas) sostenida en una zona debe de tener un limite, puesto que depende tanto de las condiciones físicas; por consiguiente, si una zona fuese habitada por muchas especies, cada una, o casi cada una de ellas estará representada por pocos ejemplares, y tales especies estarían expuestas a exterminio por fluctuaciones accidentales en la naturaleza de las estaciones o en el número de sus enemigos. El proceso de exterminio en tales casos será rápido, mientras que la producción de nuevas especies siempre debe ser lenta. Imagínese el caso extremo de que hubiera en Inglaterra tantas especies como hay ejemplares, y el primer invierno severo o el primer verano seco exterminara miles y miles de especies. Las especies raras, y toda especie se volvería rara si el número de las

92
mismas, en cualquier país aumentara indefinidamente, han de presentar dentro de un periodo determinado pocas variaciones favorable, de acuerdo con el principio explicado a menudo; en consecuencia, el proceso de dar nacimiento a nuevas formas específicas se retardaría así. Cuando cualquier especie se vuelve muy rara, el intercruzamiento estrecho contribuirá a exterminarla; ciertos autores han pensado que esto se produce y explica la decadencia del bisonte de Lituania (se refiere a los uros o toros bravíos, de los que desciende el bisonte europeo), del ciervo rojo en Escocia, y de los osos en Noruega, etc. Finalmente, y me inclino a creer que este es el factor más importante, una especie predominante, que ya ha vencido a muchos competidores en su propia patria, tendera a difundirse y a suplantar a muchas otras. Alfonso de Candolle ha demostrado que las especies que se difunden por zonas más amplias tienden generalmente a difundirse por zonas muy amplias; en consecuencia, tenderán a suplantar y exterminar diversas especies en diversas zonas y así contender el incremento desmesurado de formas específicas en todo el mundo. El Dr. Hooker ha demostrado recientemente que en el rincón sudeste de Australia, donde parece haber invasores de diferentes puntos del globo, las especies australianas endémicas han disminuido mucho en número. No pretenderé decir cuanto peso ha de atribuirse a estas diversas consideraciones; pero en conjunto son factores que limitaran en cada país la tendencia a un aumento indefinido en las formas específicas.
RESUMEN DEL CAPÍTULO
Si bajo condiciones variables de vida los seres orgánicos presentan diferencias individuales en casi todas las partes de su estructura, y esto no puede negarse; si debido a su proporción geométrica de incremento hay una intensa lucha por la vida en alguna edad, estación o año, y esto tampoco puede negarse, entonces, considerando la infinita complejidad de las relaciones de todos los seres orgánicos entre sí y con sus condiciones de vida, por lo cual resulta ventajosa para ellos una infinita diversidad de estructura, constitución y hábitos, seria extraordinario que no hubiese surgido nunca variaciones útiles para el bienestar de cada ser, del mismo modo que han surgido muchas variaciones útiles al hombre. Pero si las variaciones útiles a un ser surgen alguna vez, seguramente los ejemplares caracterizados
por ellas tendrán las mejores probabilidades de ser preservados en la lucha por la vida, y debido al fuerte principio de la herencia, tenderán a producir descendientes caracterizados de la misma manera. Este principio de preservación, o la supervivencia de los más aptos, es lo que he llamado selección natural. Conduce al mejoramiento de cada criatura en relación con sus condiciones orgánicas e inorgánicas de vida; y en consecuencia, en la mayoría de los casos, a lo que puede considerarse como un progreso en la organización. Sin embargo, formas simples e inferiores resistirán largo tiempo si están bien adaptadas a sus sencillas condiciones de vida.
La selección natural, de acuerdo con el principio de que las cualidades se heredan a edades correspondientes, puede modificar el huevo, la semilla o el hijo con tanta facilidad como el adulto. Para muchos animales, la selección sexual habrá ayudado a la selección ordinaria asegurando a los machos más vigorosos y mejor adaptados el mayor número de hijos. La selección sexual también dará caracteres útiles a los machos solamente en su lucha o rivalidad con otros machos, y esos caracteres serán transmitidos a un sexo o a los dos, de acuerdo con la forma de herencia que prevalezca. Que la selección natural haya obrado realmente así para adaptar las diversas formas de vida a sus diversas condiciones y regiones, debe ser juzgado por el tenor y el saldo generales de las pruebas que daremos en los capítulos siguientes. Pero ya hemos visto como ella implica también extinción y la geología revela claramente cuanto a intervenido la extinción en la historia del mundo. La selección natural conduce también a la divergencia de caracteres, porque cuanto más difieren los seres orgánicos en su estructura, hábitos y constitución, tanto mayor será el número que pueda ser sostenido en la región; De lo cual vemos la prueba examinando a los moradores de cualquier lugar pequeño y los productos aclimatados en tierras extrañas. Por lo tanto, durante la modificación de los descendientes de cualquier especie y durante la lucha incesante de todas las especies para aumentar en número, cuanto más diversificados se vuelven los descendientes, mejores serán sus probabilidades de éxito en la batalla por la vida. Así las pequeñas diferencias que distinguen a las variedades de la misma especie tienden constantemente a aumentar hasta que igualan a las diferencias mayores entre las especies del mismo genero y aun de géneros distintos. Hemos visto que las especies comunes, muy difundidas y que

93
abarcan regiones más extensas, pertenecientes a los géneros mayores de cada clase, son las que más varían; y ellas tienden a transmitir a sus descendientes modificados esa superioridad que ahora las hace predominantes en su propio país. La selección natural, como lo hemos observado hace un instante, conduce a la divergencia de caracteres y a mucha extinción de las formas de vida menos mejoradas e intermedias. Por estos principios pueden explicarse la naturaleza de las afinidades y las distinciones generalmente bien definidas entre los innúmeros seres orgánicos de cada clase que hay en todo el mundo. Es un hecho realmente maravilloso –cuya maravilla tenemos propensión a olvidar porque nos es familiar -que todos los animales y plantas de todos los tiempos y lugares estén relacionados unos con otros en grupos, subordinados a grupos de la misma manera en que los contemplamos en todas partes, es decir, estando las variedades de la misma especie muy estrechamente relacionadas; las especies del mismo genero relacionadas menos estrechamente y en forma desigual, constituyendo secciones y subgéneros distintos mucho menos relacionadas, y los géneros relacionados en diferente grado, constituyendo familias, ordenes, subclases y clases. Los diversos grupos subordinados de cada clase no pueden ser clasificados en un renglón comun, pues parecen aglomerados en torno a ciertos puntos, los cuales a su vez lo están en torno a otros puntos y así sucesivamente en ciclos casi interminables. Si las especies hubieran sido creadas independientemente no se podría explicar esta forma de clasificación, pero se le explica por la herencia y la acción compleja de la selección natural que acarrea extinción y divergencia de caracteres, como hemos visto en el diagrama.
Las afinidades de todos los seres de la misma clase han sido representadas a veces con un gran árbol. Creo que este símil dice en gran parte la verdad. Las ramitas verdes de los renuevos representan a las especies que existen; y las ramas producidas en años anteriores pueden representar a la larga sucesión de especies extintas. En cada periodo de crecimiento, las ramitas nuevas habrán tenido a bifucarse en todo sentido y a recuperar y matar a los renuevos y ramas circundantes, del mismo modo en que especies y grupos de especies han dominado en todos los tiempos a otras especies, en la gran batalla por la vida. Los miembros divididos en grandes ramas, éstas en ramas cada vez menores, fueron alguna vez, cuando el árbol era joven, verdes renuevos; y esta
vinculación de los vástagos anteriores y los actuales por las ramas que se bifurcan bien puede representar a la clasificación de todas las especies extinguidas y vivas en grupos por subordinados a otros grupos. De los muchos renuevos que florecían cuando el árbol era un mero arbusto, solamente dos o tres, transformados ahora en grandes ramas, sobreviven y sostienen a otras ramas; del mismo modo entre las especies que vivieron en viejos periodos geológicos, muy pocas han dejado descendientes vivos y modificados. Desde que creció el árbol, muchas ramas se han secado y desprendido, y estas ramas caídas, de diversos tamaños, pueden representar a ordenes, familias y géneros enteros que ahora no tienen representantes vivos y que solo conocemos como fósiles. Así como acá y allá vemos una fina rama desviada que brota desde muy bajo en el tronco y que por alguna suerte ha sido favorecida y llega hasta lo alto, viva aun, del mismo modo de vez en cuando un animal, como el ornitorrinco o el pez de légamo, que en cierto modo vincula con sus afinidades dos grandes ramas de vida y que al parecer se ha salvado de la fatal competencia por vivir en un lugar protegido. Como los brotes con su crecimiento, dan origen a nuevos brotes, y éstos si son vigorosos, se ramifican y superan en todas partes a muchas ramas más débiles, del mismo modo tengo la creencia que ha ocurrido por generación en el gran árbol de la vida, que llena con sus ramas muertas y rotas la corteza de la tierra y cubre la superficie con sus bellas e interminables ramificaciones.

94
LA MÁS BELLA HISTORIA DEL MUNDO
Hubert Reeves
Jöel De Rosnay
Yves Coppens
Dominique Simonnet
ESCENA 3
LA EXPLOSIÓN DE LAS ESPECIES
Las células, demasiado tiempo solitarias, se tornan solidarias. Se despliega un mundo lleno de colores: nacen las especies, mueren, se diversifican. La vida crece y se multiplica.
LA SOLIDARIDAD DE LAS CÉLULAS
En esta etapa de nuestra historia, la tierra esta poblada de células que viven apaciblemente en los océanos y que muy bien pudieron continuar así...
Pero llega un momento en que se ven obligadas a evolucionar. Las primeras células, que proliferan, se envenenan con los desechos que ellas mismas arrojan al entorno. Desde un comienzo la vida muestra una tendencia natural a agrupar a los individuos. Las sociedades “celulares” poseen ventajas evolutivas evidentes. Están mejor protegidas, sobreviven mejor que las células aisladas.
¿Cómo se van a constituir?
El comportamiento de una ameba, el dycostelium, que hoy vive todavía, nos puede ayudar a saberlo. Se alimenta de bacterias. Si se la priva de alimento y de agua, emite una hormona de ansiedad. Se le unen otras amebas y se aglomeran en una colonia de cerca de un millar de individuos que se desplazan en busca de alimento. Si no lo encuentran, de fijan, desarrollan un tallo con espora y así se quedan indefinidamente, en plena sequía. Si aparece agua, las esporas germinan y producen amebas independientes que se marchan cada una por su lado...
Los vólvox, pequeñas células provistas de flagelos, se comportan del mismo modo: en un mismo medio pobre en sustancias nutritivas, secretan una especie de gelatina, se pegan unas con otras y se desplazan en la misma dirección, con los flagelos en la parte exterior, de un modo coordinado, como formando una sola y la misma entidad.
¿Así se constituyeron los primeros organismos multicelulares?
Es probable que una lógica semejante de socialización haya actuado en los comienzos de la vida. Las primeras asociaciones de células utilizan una especie de cañería central, una suerte de cloaca general para evacuar desechos. Otras tienen forma ahusada, adelante poseen un sistema de coordinación y atrás o a los costados un sistema de propulsión. De este modo se mantienen juntas.
¿A qué se parecen estos primeros conjuntos de células?
Están compuestos por varios miles de individuos y forman pequeñas jaleas transparentes; son los primeros organismos marinos, gusanos, esponjas, medusas primitivas. Esta transformación ocurre en un lapso de sólo algunos cientos de miles de años. La evolución se acelera.
LA DIVISIÓN DEL TRABAJO
Estos nuevos ensamblajes son muy distintos de los anteriores.
Sí, la materia suele estar hecha de apilamientos e átomos idénticos unos a otros. En el mundo viviente, las células se diferencian según el lugar que ocupan en la estructura. Algunas se van a especializar en la locomoción, otras en la digestión y otras en la acumulación de energía. Poco a poco, reproduciéndose en el curso de las generaciones, estos organismos transmiten sus propiedades a su descendencia.
¿Y se puede seguir explicando este fenómeno por la mera urgencia de sobrevivir?
Sí, el organismo compuesto por células especializadas resiste mejor que un conjunto de células idénticas, pues puede responder de distintos modos a agresiones del entorno, lo que le concede mejores posibilidades de supervivencia. Los sistemas monolíticos siempre han terminado por desaparecer.
¿Pero qué empuja a estas células a asociarse? Desde luego, no se dicen “esto nos conviene para sobrevivir”...
¡Por supuesto que no! Las células no saben, es obvio, que les interesa sobrevivir. Pero poseen mecanismos de aproximación que las invita a ligarse a sus semejantes, e intercambian sustancias unas con otras. El juego de esta comunicación química y de los pequeños cambios que efectúan sus genes

95
termina por especializarlas. Se establece entonces una topografía en el grupo de células. Una medusa, por ejemplo, posee un sistema de contracción para desplazarse y un sistema sensorial que le permite dirigirse hacia el alimento. El plan del conjunto esta contenido en cada una de las células. Basta con una para que vuelva a empezar la organización.
A pesar de todo, las células que permanecieron solitarias consiguieron sobrevivir y algunas lo han hecho hasta hoy. ¿Por qué no se reagrupan éstas?
Porque estaban bien adaptadas a su entorno. Es el caso de los paramecios y las amebas: una sólida membrana las protege y están equipadas con cilios vibrátiles que les permiten desplazarse con facilidad; disponen de manchas fotosensibles que les señalan la luz y de enzimas eficaces que dirigen toda suerte de presas. Una bacteria posee hasta una especie de olfato: receptores químicos que comunican con su flagelo y la guían hacia los medios con mayor abundancia de alimento, como si sintieran el olor de la comida.
¡VIVA EL SEXO!
¿Y como van a continuar su evolución los organismos de varias células?
El árbol de la vida se desarrolla en tres grandes ramas a partir de los seres pluricelulares más simples, como las algas, las medusas y las esponjas: la de los champiñones, los helechos los musgos, las plantas de flor; la de gusanos, los moluscos, los crustáceos, los arácnidos, los insectos, y la de los peces, los reptiles, los procordados, luego las aves, los anfibios, los mamíferos...
Y después viene una invención mayor: el sexo. Hasta entonces las células se reproducían, en el sentido propio del termino: de manera idéntica. Con el sexo, dos seres vivos procrean un tercero que es distinto de ellos dos. ¿Quién fue el astuto que lo invento?
Según algunas hipótesis, el sexo habría nacido del... canibalismo: al comerse unas a otras, las células habrían integrado los genes de otras especies, que luego se habrían mezclado. Este fenómeno existe en las bacterias: algunas, bautizadas más y menos, se aparean e intercambian su material genético. Enseguida, a medida que los organismos se tornan más complejos, se van a dotar de células especializadas en la reproducción, las células germinales, que
incluyen, cada una, la mitad de los genes de su organismo. La sexualidad se generaliza.
Y desde ese momento el mundo viviente se hace más y más variado.
Es una revolución. La naturaleza puede combinar genes gracias a la sexualidad. Estalla la diversidad. Comienza la gran aventura de la evolución biológica; va ha experimentar innumerables ensayos fracasados, pistas que no llevan a ninguna parte, especies que no sobreviven... la naturaleza pone a prueba en gran escala: si la especie inventada no se adapta, desaparece.
¿Por qué la sexualidad se estabilizo entre dos? ¿Por qué no entre tres?
La mezcla de genes pone en juego, con los dos filamentos del ADN, un proceso de duplicación. Para combinar dos pares de cromosomas en un huevo fecundado, se necesita una maquinaria biológica extremadamente compleja. Y lo seria aun más si tuviera que mezclar tres patrimonios genéticos. Si hubo especies que inventaron una sexualidad de este tipo, no han sobrevivido.
LA MUERTE NECESARIA
Y se produce otro fenómeno decisivo: la introducción del tiempo en el organismo, es decir el envejecimiento y, en ultima instancia, la desaparición del individuo, la muerte. ¿No se pudo prescindir de esto?
La muerte es tan importante como la sexualidad: vuelve a poner en circulación los átomos, las moléculas, las sales minerales que necesita la naturaleza para continuar desarrollándose. La muerte realiza un gigantesco reciclaje de unos átomos cuyo número es constante desde el Big Bang. Gracias a ella, la vida animal se puede regenerar.
¿Estaba presente desde los primeros organismos?
Sí, también envejecen las medusa. Las células no dejan de reproducirse en todos los seres vivientes, pero poseen un oscilador químico, una especie de reloj biológico interno que limita a la cantidad de sus reproducciones: entre cuarenta y cincuenta. Cuando llegan a esta fase, un mecanismo programado en sus genes las conduce a una especie de suicidio. Mueren. Sólo las células cancerosas eluden esta fatalidad: se reproducen indefinidamente, sin especializarse ni diferenciarse como lo hacen las células embrionarias.

96
Pero su inmortalidad provoca la muerte del organismo a que pertenecen... ¿Se puede decir que la muerte es una necesidad de la vida?
Totalmente. Pertenece a la lógica de lo viviente. A medida que las células se dividen, multiplican los errores de sus mensajes genéticos y estos se acumulan en el curso del tiempo. Finalmente hay tantos errores que el organismo se degrada y muere. Es un fenómeno ineluctable. La muerte no es, por cierto, un regalo para el individuo, pero sí lo es para la especie: le permite conservar su nivel óptimo de desempeño.
¿Qué más puede hacer la evolución una vez que conoce el sexo y la muerte?
Perfeccionarse. El mundo viviente va a seleccionar un modo de fabricar energía; utilizando los azucares como alimento, va a enriquecer su metabolismo y desarrollar músculos, lo que le permitirá actuar, nadar, volar, correr, conquistar el mundo. Simultáneamente, los captores, que son los sentidos, coordinan las actividades del organismo. Aparecen tres grandes novedades: el sistema inmunetario, que asegura protección contra parásitos o virus; el sistema hormonal que permite el control de los ritmos biológicos y de la reproducción sexuada, y el sistema nervioso, que rige la comunicación interna.
¿Cuándo aparece este último?
Los primeros organismos, medusas, peces primitivos, necesitan coordinar sus células para reproducirse. Cuentan por lo tanto con canales especializados por donde circula la información. Un gusano, que apenas esta compuestos por algunos miles de células, posee fibras nerviosas que convergen en su cabeza, ganglios. En el curso de la evolución, este dispositivo se va a ramificar para formar una red de neuronas interconectadas que se reunirán en un cerebro. De hecho, los tres sistemas, nervioso, hormonal e inmunitario, aparecen apenas los animales salen del agua.
EL REGALO DE LAS LÁGRIMAS
¿Qué los impulsa a salir del agua?
Las especies pululan en los océanos. Reina la competencia. Aventurarse a tierra firme parece ventajoso para conseguir alimento, pero volviendo al océano para poner los huevos. El primero que experimento esta formula fue sin duda un pez extraño, el ictiostega. Posee grandes aletas, vive en
pequeñas lagunas y saca del agua de vez en cuando sus ojos globulosos para buscar pequeños insectos. En el curso de las generaciones, los descendientes de esta especie se arriesgan más tiempo en tierra firme gracias a unas branquias que les permiten capturar oxigeno del aire, pero también gracias a las lagrimas: tienen que conservar húmedos los ojos para poder ver tan bien en tierra como en el agua. La especie mejora por sucesivas selecciones: las aletas se tornan más sólidas, aparece una cola. Sus descendientes serán los batracios y los anfibios. ¿No estaríamos aquí si este pez no hubiera tenido lagrimas!
¿La vida al aire libre favorece la evolución?
Sí, la comunicación es más inmediata en el aire, más rápida, más sencilla. Y mayor la accesibilidad del alimento. Sin embargo, el oxigeno es un veneno para la vida: Contribuye al nacimiento de radicales libres, moléculas desequilibradas, que inducen la destrucción celular y por lo tanto el envejecimiento precoz; pero es esencial para dotar de energía a los organismos y hacer avanzar la evolución.
¿Y como van a acelerar el perfeccionamiento de los organismos estas restricciones del medio?
Con la aparición del esqueleto, los animales se tornan más sólidos y se liberan del peso. La invención de los músculos les permite dejar de ser bolas de gelatina muelle como los gusanos de tierra o las medusas; ahora pueden ejercer presión mecánica sobre el entorno, soportar el peso de la grasa protectora y del cerebro. Todo se diversifica: el metabolismo, los sistemas de locomoción... durante este tiempo se seleccionan en las plantas los sistemas para captar la energía solar con las hojas y para transportar energía con la savia.
EL OLFATO DE LOS VEGETALES.
¿Y por que los vegetales no desarrollaron todas estas maravillas que aportaron los animales?
Con la excepción de las algas, que evolucionaron en la superficie de los océanos, los vegetales se ingeniaron un camino más económico gracias a su inmovilidad, que les permite gastar menos energía. Su modo de vida es sencillo: fotopilas para transformar directamente la energía solar en energía química, raíces para extraer sales minerales y agua... lo astuto es su sistema reproductor, que es móvil y utiliza variados medios.

97
También los vegetales han heredado una sexualidad muy rica y están adaptados maravillosamente bien. Basta, para comprobarlo, observar un champiñón al pie de una secoya gigante de varios miles de años de edad. O, sencillamente, basta mirar los triviales pinos de montaña.
¿Y en que se advierte que resultan de una buena adaptación?
Necesitan de una determinada temperatura para desarrollarse en el bosque. Tal como las margaritas de nuestro planeta maginario, los árboles sombríos y negros capturan mejor el débil resplandor solar, calientan el entorno inmediato y crean un microclima favorable para su crecimiento. Pero en el invierno se cubren de nieve y quedan blancos. Si se mantuvieran así demasiado tiempo, ya no podrían asegurar las condiciones propicias. Ahora bien, como tienen las ramas inclinadas hacia abajo y en punta, la nieve se sostiene menos tiempo; recuperan su color y se calientan rápidamente. La evolución ha mantenido el tipo de árbol que mejor resiste la intemperie. Por eso hay pinos en las montañas...
Y uno se maravilla por esa fantástica adaptación. Una pregunta ingenua: ¿Por qué los vegetales no desarrollaron un cerebro?
Seres inmóviles no necesitan funciones complejas de coordinación. No les impulsa la necesidad de huir o de luchar como los animales. Comenzamos, no obstante, a descubrir, en las plantas, una forma de sistema inmunitario, un sistema de comunicación e incluso un homologo del sistema nervioso. Los vegetales poseen sofisticados mecanismos que los protegen contra invasores: una suerte de “hormona” vegetal les permite, por ejemplo movilizar sus defensas. Se sabe, también, que los árboles se “avisan”, a distancia, la presencia de un agresor.
¿Avisan?
Sí, cuando están en presencia de animales predadores que les quieren comer las ramas bajas, algunos árboles emiten productos volátiles que transportados de árbol en árbol modifican la producción de proteínas y dan a las hojas un gusto desagradable. ¡Yo no iría tan lejos, sin embargo, como para decir que hay que hablar con las plantas de interior!
¿En todo caso se puede afirmar que los animales son los que han llegado más lejos en cuanto a complejidad?
Es verdad que el mundo animal, en efecto, demuestra mayor exuberancia que el mundo vegetal en su adaptación al medio: hay especies que corren, que excavan, que nadan, que vuelan, que se arrastran... los animales desarrollan innumerables trucos, desde las presiones del abejorro hasta los tentáculos del pulpo, inventan trampas, cebos, armas, garras, alas, picos, aletas, caparazones, tentáculos, veneno...
LA EXCLUSIÓN NATURAL
Cuando se dice “inventan”...
No inventan. El fenómeno de la “selección” elimina a los menos aptos. Consideremos por ejemplo los gorriones de gran pico que se alimentan exclusivamente de pequeños gusanos ocultos en agujeros de los árboles. Son tan númerosos y activos que terminan por eliminar a todos los gusanos que hay en la superficie de las cortezas. Sin alimento, la mayor parte muere. Pero unos pocos poseen, por una mutación ocurrida al azar, un pico en punta más largo que el de los otros. Sus descendientes pueden ir a buscar gusanos en agujeros más profundos y resisten mejor la escasez. Resultado: este linaje se impone. Con el curso de las generaciones, la mayoría de la especie va a poseer un pico más largo. No se puede decir, sin embargo, que los gorriones “inventaron” este recurso. En realidad es al revés: murieron los que no tuvieron la fortuna de una mutación que les dio un pico más largo.
En el proceso de evolución no hay, entonces, intencionalidad.
No. La evolución intenta miles de soluciones al mismo tiempo; unas tienen exito y otras no. Se conservan, por definición, las que permiten sobrevivir.
¿Y el medio no actúa directamente sobre la evolución?
Hoy se considera que quizás tiene alguna influencia en el comportamiento de las células, por intermedio de las mitocondrias, esas fábricas que en el interior de las células poseen planes genéticos independientes y son muy sensibles a los cambios. Pero esto no se transmite a la descendencia.
¿Entonces el principio de la selección natural sigue siendo pertinente hoy?
Sí, pero a condición de que no se vea en ello la noción de un entorno demiurgo que

98
decidiría lo que esta bien y lo que no. Esto se mantiene y esto se desecha. No. Hablemos, mejor de exclusión competitiva: en el curso de las generaciones, se excluye a las especies menos adaptadas. Para comprender mejor este fenómeno hay que contar con el curso del tiempo y pensar en una larga cadena de generaciones sucesivas que se modifican muy lentamente.
Una mayoría aplastante de soluciones, de especies inventadas por la naturaleza, desaparece. ¿No hay momentos en que la evolución haya intentado detenerse, en que el mundo viviente puede hallar estabilidad, como las margaritas de nuestro planeta?
No. La diversidad es enorme desde el comienzo de la vida. Retomando la metáfora de Hubert Reeves, hay demasiadas letras como para que solo formen una palabra única. ¿La estabilidad de alguna especie gastada ha podido establecerse en algunos pequeños asteroide, constituyendo una suerte de compromiso o armisticio de la evolución? Pero no en toda la Tierra, que tiene unas dimensiones, una geología, una biosfera, una relación entre lo mineral y lo orgánico y un entorno en cambio constante que obliga a las especies a modificar su adaptación y evolucionar.
Y eso ocupa algunos cientos de millones de años.
Sí. Esta selección actúa sobre millones de generaciones sucesivas. Los mecanismos sensoriales se afinan. Los comportamientos se diversifican. Algunas especies se asocian y forman un verdadero organismo colectivo. Un panal de abejas, por ejemplo, mantiene la temperatura gracias al movimiento de las alas de los insectos; está irrigado por las hormonas que resultan del frotamiento de los insectos. Cuando las abejas dejan del panal para buscar alimento, indican con una danza las fuentes más cercanas. Así el panal economiza energía; optimiza sus posibilidades de sobrevivencia. Lo mismo ocurre con las hormigas: mantienen a las larvas, ayudan a la reina, se reparten las tareas, un poco como las células del vólvox, y aseguran el equilibrio del organismo hormiguero. Si se quita el treinta por ciento de las obreras, el conjunto se va adaptar y restablecerá la proporción.
Pero las hormigas no son capaces de comportamientos autónomos.
E incapaces de planificar. Se comunican individualmente por las feromonas, pero también colectivamente por el entorno: una
hormiga joven va a aprender las redes, los caminos que han trazado sus congéneres.
El comportamiento simultaneo de miles de individuos conduce a una forma de inteligencia colectiva. La hormiga, por ejemplo, sabe escoger el camino más corto para traer alimento. Este modo de asociación ha tenido bastante éxito, porque las hormigas existen hace millones de años. Si el planeta experimentara una guerra nuclear, es probable que sobrevivan gracias al caparazón que les permite resistir las radiaciones y gracias a su modo de organización.
EL INFORTUNIO DE LOS DINOSAURIOS.
Un mundo de hormigas y bacterias... hermosa perspectiva. En el curso de este relato se puede apreciar que, como la del universo, la evolución de la vida ha sido, por lo menos, caótica.
Sí. Ha experimentado una aceleración constante, pero también crisis, caminos sin salida y periodos de grandes extinciones. Los dinosaurios reinaban en el planeta hace doscientos millones de años. Las especies jamas habían logrado conquistar, como ellos, todos los ambientes. Había pequeños, enormes, vegetarianos, carnívoros, corredores, voladores, anfibios... una diversidad formidable, que les permitió adaptarse a sus entornos.
Y sin embargo desaparecieron... ¿Entonces es estúpida la hipótesis de que eso se debió a su mala adaptación?
Totalmente. A finales del jurásico, hace sesenta y cinco millones de años, cayo en el Golfo de México, cerca de Yucatán, un enorme meteorito de cinco kilómetros de diámetro. El choque fue tal que repercutió al otro costado del planeta y provoco un resurgir de magma. Este golpe doble creó un incendio mundial, se abrasaron los bosques, se libero gas carbónico y polvaredas cubrieron la Tierra con un velo intenso. El planeta se oscureció, se produjo un frío terrible y, probablemente, un posterior efecto invernadero que condujo a un recalentamiento.
¿Solo sobrevivieron algunas especies?
Sí. Es el caso de los lémures, que son muy móviles, adaptables y están provistos de manos prensiles. Se refugiaron en las grietas de las rocas y originaron los linajes que condujeron a los mamíferos. Estos adquirieron una nueva ventaja para asegurar la

99
supervivencia de su descendencia; llevar el huevo internamente lo protege mucho más que si queda en el exterior. Pensemos en los batracios, que ponen miles de huevos que se dispersan, son comidos, se pierden...
LA SELECCIÓN EN LA CABEZA
¿En qué momento aparece verdaderamente el verdadero cerebro?
Desde los peces, y después con los vertebrados, los pájaros, los reptiles, los anfibios y el hombre, el cerebro no ha cesado de perfeccionarse por estratos sucesivos. Primero, el más primitivo: el de los reptiles, que coordina los instintos primarios de supervivencia, hambre, sed, el instinto sexual, el miedo, el placer que impulsa a la unión y el dolor que no se le puede disociar. Ante un intruso, el cerebro primitivo reacciona y conduce al organismo a producir un veneno o a saltar sobre el agresor... segundo estrato, en los pájaros: el mesoencéfalo, que conduce a mecanismos colectivos como el cuidado de los pequeños, la construcción del nido, la búsqueda de alimento, el reparto, el canto, las exhibiciones amorosas... el tercer estrato aparece enseguida en los primates y sobre todo en el hombre; la corteza cerebral que proporciona datos abstractos, la conciencia, la inteligencia.
Lo más asombroso es este principio de selección, que se encuentra por todas partes, en el universo, en la primera química de las moléculas, en los seres vivos y, si debemos creer al neurobiólogo Jean Pierce Changeux, en el interior mismo del cerebro cuando se desarrolla en un recién nacido.
El desarrollo del sistema nervioso también obedece, en efecto, al principio darwiniano de la selección. Cuando crece un pequeño animal, sus neuronas se relacionan mediante un sistema que obedece a un plan de orden genético. Pero el empalme entre neuronas solo subsiste si estas funcionan en un circuito, si el entorno las solicita.
Las neuronas visuales de un recién nacido no se conectan si a éste se le mantiene continuamente en la oscuridad. Hay, pues, de algún modo, una selección que solo mantiene los circuitos pertinentes. Aprender es eliminar.
Según el antropólogo Stephen J. Gould, cada suceso, por insignificante que sea, influye el curso de la historia. Como en La vie est belle, la película de Frank Capra, basta modificar una nadería para que todo cambie con toda una catarata de consecuencias. Si no hubiera
aparecido el pikaka, un gusano que esta en el origen mismo de nuestro linaje, o si hubieran sobrevivido los dinosaurios, no estaríamos aquí. No habría ningún sentido, según él, en la evolución. Esta no retendría a los mejor adaptados, sino a los más afortunados. La vida quizás fuera aconteciendo probable, pero el hombre un verdadero afortunado.
Si los lémures no hubieran sobrevivido ni podido alimentarse de bayas en sus agujeros mientras desaparecían los dinosaurios, no estrían aquí. No hay una intención oculta en esta historia. Pero el resultado es que aumentara la complejidad. Si existen planetas que hayan desarrollado en las mismas condiciones que la Tierra, no es improbable que esos seres existan y que no se diferencien de nosotros más que un avestruz de un cocodrilo: cuatro miembros, dos ojos, un cerebro, sistemas locomotrices. Y hay una fuerte posibilidad de que estén en el mismo punto evolutivo que nosotros... no se puede afirmar que exista una ley que impulse a la complejidad. Pero comprobamos que alguna cosa se organiza y conduce a una inteligencia cada vez mayor y más desmaterializada. Quizás la historia de la evolución es el artefacto de una conciencia que adquiere conciencia de sí misma.
LA MEMORIA DE LOS ORÍGENES
Solo el cerebro humano se interroga sobre sí mismo... ¿Esto le distingue de los demás?
No sólo eso. Es capaz de exteriorizar funciones en el medio. La herramienta prolonga la mano. El hombre puede hacer hoy todo lo que hacen los demás animales: correr como una gacela con un automóvil, volar como un águila con un ala delta, evolucionar bajo el agua como un delfín, avanzar bajo tierra como un topo... una mascara, anteojos, un paracaídas, alas ruedas... ha ampliado también sus funciones sensoriales mediante la escritura, que permite conservar la palabra y transmitir el pensamiento en el espacio y en el tiempo. Esto caracteriza al cerebro humano. No es sólo una masa muelle de neurona, ni una estación telefónica que agrupa los circuitos del cuerpo, ni siquiera un ordenador. Se extiende también al exterior, acoplado a otros cerebros humanos en el conjunto del planeta. Es una red fluida, en continua reorganización, que reconfigura sus neuronas en la acción y la reflexión.
En toda esta historia, se comprueba que la complejidad se desarrolla con la disposición de cosas simples: dos cuarks en el comienzo del

100
universo, cuatro átomos simétricos para el carbono, solo cuatro bases para los genes, dos moléculas semejantes para fundar los mundos animal y vegetal, dos individuos para el sexo... como si en cada etapa la naturaleza encontrara el camino más sencillo para progresar.
De algún modo... complejidad no es complicación. Es una repetición de elementos simples que se reproducen y proliferan. Hoy sabemos simular este fenómeno en una pantalla de ordenador: partiendo de una forma elemental vemos que se constituyen dibujos elaborados a los que llamamos con el hermoso nombre de “formas fractales”; parecen alas de mariposa, colas de hipocampos, montañas, nubes. La vida es así, repetitiva. El átomo esta en la molécula que está en la célula que esta en el organismo que está en la sociedad...
llevamos en nosotros entonces, las huellas de estos ajustes...
Exacto. Nuestro cerebro, con sus tres estratos, conserva la memoria de la evolución. También nuestros genes. Y la composición química de nuestras células es un fragmento pequeño del océano primitivo. Hemos guardado en nosotros el medio del que salimos. Nuestro cuerpo relata la historia de nuestros orígenes.

101
LA EVOLUCIÓN LETICIA DURAND SMITH
INTRODUCCIÓN
En este texto el maestro encontrara un resumen del desarrollo histórico de las ideas evolutivas, desde sus inicios hasta nuevas controversias planteadas a la síntesis evolutiva. Así mismo se aclaran conceptos como selección natural, adaptación y deriva genética. Entre otros.
A pesar que la evolución es un proceso que se desarrolla a lo largo de millones de años y difícilmente podemos apreciar, sus resultados y las ideas generadas en torno a ellas nos rodean desde pequeños. Es común por ejemplo, escuchar que nuestros parientes más cercanos son los monos, que el hombre es el animal más evolucionado o que los dinosaurios dominaron la Tierra hace mucho tiempo y desaparecieron después en una extinción masiva. Aunque algunas de estas ideas no son del todo correctas, forman parte de un conocimiento extendido y cotidiano que ya se encuentra en la división de la realidad de los alumnos cuando ingresan a secundaria. De esta forma, la evolución es un tema de importancia central en la enseñanza de biología en secundaria, ya que por un lado sirve de puente entre las ideas comunes a las que esta expuesto el alumno en su vida cotidiana y los contenidos del programa de biología, y por otro, permite que temas íntimamente relacionados con los procesos evolutivos sean abordados desde esta perspectiva, dejando de ser temas aislados o difíciles de explicar (como por ejemplo genética o estructura y función de los seres vivos), lo que ofrece la posibilidad de desarrollar una amplia interconexión entre los puntos del temario de biología para secundaria.
EVOLUCIÓN: IDEAS E HISTORIA
El hombre ha dependido siempre de su relación con otros seres vivos. De ellos obtiene alimento, vestido, protección, medicamentos y hasta compañía. Esta estrecha relación ha provocado la necesidad de conocer a los organismos, no solo para poder aprovecharlos sino para saciar la curiosidad que plantas y animales producen en el hombre. Cuando aun no existían los instrumentos, transportes y medios de comunicación que conocemos actualmente, la fascinación por lo vivo ya trabajaba en la mente de las personas, quienes producían
explicaciones y descripciones de sus observaciones. Así, los médicos recetaban el polvo del cuerno de unicornio como un eficaz desintoxicante pero decían que debía administrarse con cuidado ya que toda bebida a la que se le añadían limaduras del cuerno de este animal comenzaba a bullir y a calentarse en el acto; y los marineros al regresar de sus travesías afirmaban haber visto serpientes de mar de treinta metros de longitud, de color marrón, con cabellera en la espalda y ojos rojos que intentaban enroscarse en la embarcación y comer a sus tripulantes. Todas estas historias y muchas más han ido poco a poco desapareciendo, siendo reemplazadas por modelos reales, fruto de una actividad humana muy importante que también deriva de la imaginación: La ciencia.
Sin embargo, como ya se menciono, los alumnos al ingresar a secundaria poseen una carga de ideas previas sobre la naturaleza. La importancia de estas ideas, sean o no correctas, reside en que son un primer acercamiento al mundo que observan y que apartir de su modificación en el salón de clases puede producirse un aprendizaje significativo; Es decir, un aprendizaje que se relacione con una mayor cantidad de situaciones vividas por los estudiantes y permita explicar con mayor profundidad la realidad a la que están expuestos. De una u otra forma, la ciencia trabaja del mismo modo que la mente de los alumnos, a partir de metodológicas claras y de una serie de conocimientos ya acumulados permite explicar con mayor claridad los fenómenos naturales, modificando gradualmente las ideas y concepciones.
La teoría de la evolución biológica es resultado de una serie de ideas que se han forjado a lo largo de la historia y, como todo conocimiento científico, se ha ido modificando de acuerdo con los avances obtenidos. Para algunos griegos, como Anaximandro y Empédocles (600-500 a. C.), la vida se había originado a partir de las distintas mezclas posibles entre los cuatro elementos: fuego, aire, tierra y agua. Para Anaximandro, estos elementos dieron origen a los primeros seres inferiores, de los cuales surgió el hombre como un ser superior. Para Empédocles, los cuatro elementos habían originado una gran cantidad de formas de vida compuestas por partes de los diferentes animales, pero no fueron solo los que estaban conformados de la manera adecuada, es decir, como los conocemos, quienes sobrevivieron.
Un siglo después, Platón origino una idea de la vida que prevaleció durante mucho tiempo, la

102
de un mundo estatico e inmutable. Para Platón todo aquello que existía sobre la tierra eran meras reproducciones inexactas de una esencia perfecta e inalterable que existía únicamente en el mundo de las ideas. Para él, los organismos eran simples reproducciones de la esencia original, por lo que la variación no existía o estaba reducida a imperfecciones en torno a la esencia. Poco después de Platón, Aristóteles creó un sistema de clasificación para los organismos al cuál llamó Scala Naturae, en el cual estos se agrupaban en una jerarquía que distinguía entre los organismos superiores e inferiores.
Con el surgimiento del cristianismo se modifico la idea sobre la naturaleza, pro se mantuvo como base la concepción de un mundo inmutable, ya que la idea de la modificación era inconsistente con los planteamientos religiosos, pues llevaba directamente a dudar a cerca de la perfección de las creaciones de Dios, y por lo tanto de su propia perfección. Los trabajos de los naturalistas eran siempre adaptados o moldeados de acuerdo con las ideas religiosas, indicando que la adaptación y perfección de los organismos eran muestra del poder de Dios. Poco a poco la unión entre ciencia y religión fue siendo cada vez más conflictiva. Newton, Galileo y Descartes, entre otros, mostraban que era posible explicar fenómenos naturales a partir de ideas que podían estudiarse y comprobarse la idea de un mundo cambiante y cognoscible fue ganando más terreno.
A principios del siglo XIX comenzaron a forjarse las primeras ideas sobre el cambio en las especies. Específicamente fue Jean Baptiste Lamarck quien en 1809 propuso que los seres vivos están expuestos a una serie de cambios a lo largo del tiempo. En su texto Filosofa Zoológica, Lamarck plantea preguntas que intentaron ser resueltas más adelante:... desde los restos fósiles que se encuentran... un gran número de ellos pertenecen animales de los cuales no se conocen análogos vivientes y perfectamente semejantes... ¿Pueden éstas conchas fósiles pertenecer a especies perdidas? ¿Cómo se habrían perdido si el hombre no ha podido obrar su destrucción? ¿No seria posible, al contrario, que los individuos fósiles de los grupos que se trata pertenecieran a especies todavía existentes, pero que hubiera cambiado dando lugar a las especies vivas que no parecen sus vecinas?
Lamarck respondió estas preguntas a partir de su teoría de la adaptación por la herencia de caracteres adquiridos, mencionando que
aquellos caracteres que eran adquiridos durante la vida de un organismo podían ser heredados a sus descendientes y explicando que los seres vivos tienden siempre a una mayor complejidad y perfección regidos por fuerzas divinas. Hoy sabemos que la idea de los caracteres adquiridos no es correcta y que los organismos tampoco tienen un deseo de superación. Aunque Lamarck no llegó a formular una teoría contundente sobre la evaluación, su trabajo es sumamente importante, pues propone por vez primera el cambio dentro del mundo vivo y además reúne una gran evidencia en este sentido. Sin embargo, fue Charles Darwin quien se encargó de aclarar y proponer mecanismos que desterraron a los mitos de la evolución.
La vida de Charles Darwin (1089-1882) fue una extraña mezcla de aventuras personales en su juventud, que posteriormente se convirtieron en aventuras intelectuales que ocuparon el resto de su vida, inmersas en una existencia tranquila. Darwin fue capaz de producir una teoría que modifico para siempre la visión del hombre sobre la naturaleza y marco el rumbo definitivo de la biología, pero lo más importante de su vida fue su completa entrega al quehacer científico, muestra del gran entusiasmo y atracción que produce la ciencia, capaz de absorber el tiempo, las ideas y la vida de las personas, a cambio del placer de descubrir y conocer.
Se ha escrito mucho sobre Darwin, existen diversas biografías y ediciones de sus diarios de campo, así como recopilaciones de cartas personales, que por si mismas son de gran interés como testimonio histórica y ejemplo del intricado origen de las teorías y descubrimientos científicos.
Una de las principales etapas del viaje de Darwin fue su visita a las islas Galápagos. En estas islas observo que cada una poseia una especie particular de tortugas que los isleños podían reconocer a partir de las diferentes formas que representaban en sus caparazones. También notó que había variación en las especies de pinzones que habitan las islas. Cada isla estaba habitada por una especie de pinzón, semejantes ente si pero que ocupaban nichos ecológicos diferentes alimentándose algunas de insectos, otras de semillas, etc. Sin embargo aún era muy pronto para que Darwin relacionara las semejanzas entre estas especies con un origen común y sus diferencias con un aislamiento reproductivo y la diferenciación de nichos ecológicos, aunque en este momento es cuando comienzan a originarse a la mente de Darwin las primeras ideas sobre la

103
modificación de las especies y su mecanismo de evolución. Otras observaciones importantes que realizo durante su viaje fueron aquellas relacionadas con la presencia de conchas fósiles en zonas muy alejadas del mar y que guardaban gran parecido con las conchas actuales que habían observado en las costas.
Durante su travesía Darwin recibió por correo en diferentes puertos varios libros. Uno de ellos fue Ensayo sobre el principio de la población, escrito por Thomas Malthus. Este texto fue básico en la formulación de las ideas de Darwin sobre la selección natural. Estudiando poblaciones humanas Malthus se dio cuenta de que los organismos vivientes producían un número mayor de descendientes que los que pueden sobrevivir por la limitación de recursos, como el alimento o espacio. Por ello, Darwin supuso que debería existir algún mecanismo que regulara las probabilidades de sobrevivencia o mortalidad de los individuos.
Gracias a sus conocimientos sobre la cría de animales domésticos, Darwin fue capaz de relacionar el problema planteado por la sobrepoblacion con la variación individual.
Éstas y muchas más observaciones fueron las que sirvieron a Darwin como materia prima y evidencia para postular su teoría de la evolución por la selección natural, aunque ésta fue postulada 28 años después del día que Darwin zarpó en el Beagle. Los conocimientos adquiridos durante este viaje los enriqueció en Inglaterra durante muchos años, en los que se dedico a una cuidadosa labor de clasificación y revisión, que sentó la posibilidad de reflexionar sobre lo ya vivido y generar nuevos datos. Esta forma de trabajar clásica de un científico, le permitió dominar información especifica y al mismo tiempo enmarcarla dentro de un esquema teórico general conformado por la selección natural como fuerza evolutiva.
Si resumimos las ideas de Darwin podemos decir que las condiciones para que se desarrolle un proceso de evolución por selección natural son; la existencia de variabilidad individual, el carácter heredable de esa variación y la limitante ambiental que promueve la competencia e impide el establecimiento de un número infinito de individuos. Es importante señalar que los individuos presentan variabilidad en una gran cantidad de caracteres morfológicos, fisiológicos o conductuales, pero no todos están expuestos a selección natural, ya que ésta sólo actúa sobre aquellos caracteres que confieren ventajas o afectan el desempeño de
los individuos; es decir, favorece la reproducción de aquellos individuos con rasgos que aportan beneficios y elimina a individuos con características negativas. El hecho de que una característica tenga consecuencias positivas o negativas en la reproducción y supervivencia, o no las produzca, depende, obviamente, del ambiente que se desarrolle el organismo, porque no existen caracteres que por sí mismos sean buenos malos. Por ejemplo, en la ficha 38 del Libro para el maestro de Biología se menciona que el color de los insectos, que representan los frijoles o la fichas utilizadas en esa actividad, es una característica que tiene afectos sobre la sobrevivencia, pues su coloración hace que sean más o menos visibles a los depredadores, siendo por lo tanto un rasgo expuesto a la selección natural. Pero la facilidad con que los alumnos, que representan a los depredadores, encuentren uno u otro color de “insecto” dependerá del color del piso sobre el que las fichas sean arrojadas. Esto significa que la evolución por selección natural no depende únicamente de la existencia de diferentes características entre los individuos sino de un conjunto de interacciones entre estas características y las condiciones, tanto físicas como biológicas, que rodean al organismo.
Otro aspecto importante en la teoría Darwinista es la adaptación. Para Darwin, aquellos organismos con características más adecuadas para desarrollarse en un ambiente particular son los que sobreviven y dejan un mayor número de descendientes, a quienes heredan sus rasgos ventajosos. Esto provoca que la selección natural ajuste gradualmente a los organismos a las exigencias del ambiente en un proceso denominado adaptación. La adaptación es, por lo tanto, una consecuencia de la selección natural y no resultado de la creación divina, como lo proponían los griegos, ni de una voluntad de perfección presente en los organismos, como lo sostenía Lamarck.
Tampoco podemos hablar de organismos más o menos adaptados u organismos superiores o inferiores, ya que la propia existencia de cada individuo nos confirma que puede desarrollarse en un ambiente particular estando por lo tanto adaptado a él, y esta adaptación no se incrementa con el tamaño o la complejidad de su estructura y funcionamiento. ¿Por qué? Básicamente porque la evolución es un proceso que no tiene dirección definida y la selección natural no actúa a favor de soluciones particulares; simplemente permite la reproducción de

104
organismos que presenten características que los hace competitivamente más eficientes, esto es independiente de que estas características sean simples o complejas. En este sentido, es común pensar que los organismos de gran tamaño son superiores, como si una talla mayor fuera sinónimo de un mejor desempeño. Por el contrario, el tamaño puede ser una limitante para muchos organismos. Por ejemplo, algunos parásitos, como la “solitaria” puede tener varios metros de longitud, pero su ancho no puede sobrepasar uno o dos centímetros, pues el oxigeno y los alimentos penetran directamente por la superficie externa, y si su tamaño fuera mayor no alcanzarían a difundirse en todo el cuerpo. La capacidad de muchos insectos de volar enormes distancias se debe a su pequeño tamaño, ya que el peso que se levanta en vuelo se incrementa cúbicamente con relación al tamaño del organismo, así que si las abejas crecieran tanto como un gran mamífero o más, como se ha visto en algunas películas de ciencia-ficción, estarían condenadas a permanecer en la tierra y ni siquiera podrían sostenerse sobre su patas. Estos ejemplos nos demuestran que el tamaño, al igual que muchos otros caracteres, puede estar expuesto a la selección natural, de tal forma que en muchos organismos exceder cierta talla significa una disminución en su eficiencia.
LA SÍNTESIS EVOLUTIVA
Darwin propuso en su teoría de evolución por selección natural que aquellas características que aportaban ventajas a los individuos en términos de reproducción y sobrevivencia, deberían transmitirse de padres a hijos y de este modo extenderse dentro de la población, para ocasionar los cambios graduales que a largo plazo originarían nuevas especies, sin embargo, a pesar de que es claro que los hijos se parecen a sus padres, uno de los grandes problemas de Darwin fue que ignoraba cuál era el mecanismo que permitía la transmisión de caracteres de una generación a otra, por lo que existía una incomoda laguna en su teoría para la que nunca encontró una solución convincente.
Parte del problema de Darwin fue que no centró su atención en la herencia de un carácter en particular, de tal forma que sus esfuerzos por encontrar una explicación sobre la herencia de los caracteres se diluyeron en la complejidad de un análisis de los cambios generacionales sufridos por los organismos como un todo. Esto le permitió a Darwin
proponer únicamente explicaciones como las de las gémulas o la herencia por el uso y desuso, que nunca lo convencieron, ni a él ni a la comunidad científica. La solución al problema de la transmisión de los rasgos de una generación a otra surgió de los trabajos de Gregor Mendel, quien descubrió las leyes de la herencia. Mendel propuso que las características de los organismos eran transmitidas por unidades indivisibles, que hoy en día llamamos genes, que podían presentarse en formas dominantes o recesivas y se heredaban de manera independientes unas de otras.
Aunque Mendel fue contemporáneo de Darwin, sus trabajos no se conocieron sino hasta 1900, y fue también a principios de ese siglo cuando se realizaron otra serie de descubrimientos importantes para aclarar los procesos hereditarios. Morgan, con sus estudios sobre la mosca de la fruta, Drosophila melanogaster, estableció que las mutaciones que producían cambios importantes ocurrían con poca frecuencia. Una serie de investigadores dedicados a la genética de poblaciones, entre los que se puede mencionar a R.A. Fisher, J.B.S. Haldane y S. Wright, demostraron que las mutaciones pequeñas son más frecuentes que las macromutaciones y tienen un mayor efecto que el que se les había asignado hasta entonces. Otros, como G.G. Simpson, dedicado a la paleontología, unió la información del registro fósil a los postulados de la teoría de la genética de poblaciones y E. Mayr, dedicado al estudio de las aves, desarrollo la base de la taxonomía evolutiva moderna. Todos estos adelantos en la biología produjeron la necesidad de reconsiderar la teoría darwinista y evaluar su compatibilidad con estos nuevos hallazgos. La teoría de la evolución por elección natural planteada por Darwin fue capaz de sostenerse y asimilar los nuevos conocimientos de la genética y otras disciplinas, de tal forma que la propia teoría de la evolución se ha ido modificando y ha integrado nuevos conocimientos para convertirse en la base para la explicación de la biología. Aunque sabemos que en muchos aspectos Darwin no pudo ser lo suficientemente convincente, el cuerpo central de su teoría se mantiene como eje fundamental de la teoría moderna de la evolución o neodarwinismo.
El neodarwinismo, al igual que el darwinismo, sostiene la existencia de la selección natural como fuerza evolutiva y resultante de tres hechos básicos: sobrepoblación, variabilidad y herencia.

105
Según los neodarwinistas, el incremento en individuos con caracteres que promueven el éxito en la sobrevivencia y la reproducción, así como la eliminación de aquellos que no los tienen, conduce gradualmente al origen de nuevas especies, en un proceso denominado especiación, del que son parte importante el aislamiento y las modificaciones ambientales.
Sin embargo, el neodarwinismo propone además otros mecanismos para el surgimiento de nuevas especies. Por ejemplo en muchas plantas es muy frecuente la aparición de poliploidías, o individuos con más de un juego de cromosomas, lo que significa que de una generación a otra pueden aparecer mutantes que son física y genéticamente diferentes a sus progenitores. En este caso la aparición de nuevas especies nada tiene que ver con la acumulación gradual de rasgos beneficiosos.
Otro mecanismo por el cual pueden aparecer nuevas especies de forma azarosa es la deriva génica, que puede equiparse con lo que en estadística se conoce como un error de muestreo. Imagine que tenemos una población de insectos en donde unos son rojos, otros verdes y otros azules.
En algún momento ocurre una catástrofe, como un incendio en el bosque donde habitan o una inundación, de tal forma que solo sobrevive una pequeña parte de los insectos que se encontraban en algún tipo de refugio. Pero por azar esa pequeña población de insectos es únicamente de color azul, de tal forma que los genes que contienen la información para el color verde y amarillo se encuentran en una proporción tan baja entre los individuos sobrevivientes que se hace muy difícil lleguen a expresarse, con lo que obtenemos una nueva población de insectos que siempre son azules. A esto se le conoce como deriva génica y se dice que se parece a un error de muestreo porque siempre que realizamos alguna investigación o estudio trabajamos con una muestra o una parte de una población o conjuntos de objetos, ya que es imposible trabajar con la totalidad de la población o conjunto.
Una de las características que debe tener esta muestra para que nuestro estudio salga bien, es ser siempre representativa; es decir, debe incluir todas las clases de objetos que contiene nuestro conjunto original o toda la variabilidad presente en nuestra población. En el caso de la población de insectos de nuestro ejemplo, una muestra representativa seria aquella que incluyera individuos de los tres colores existentes, pero en un procesos de deriva génica esto no ocurre. Solo sobrevive
una parte de la población que no la representa en su conjunto, como en nuestro ejemplo, donde solo sobrevivieron los azules, reduciendo la variabilidad de la población y posibilitando el origen de nuevas especies.
Es muy importante comprender que la evolución actúa no solo por la selección natural sino también por procesos azarosos, como la deriva génica y la macromutaciones, en donde a diferencia de la selección natural las características de los individuos no se ajustan gradualmente a las exigencias del ambiente, sino que son cambios bruscos en la composición de una población o individuo, que si resultan favorables se conservan.
Otro punto importante del neodarwinismo o teoría sintética de la evolución es que, a diferencia de Darwin, quien pensaba en la selección natural como una tajante elección entre organismos “exitosos” y “fracasados”, la selección se considera como una fuerza que moldea a los organismos y solamente puede detectarse a lo largo de muchas generaciones y dentro de poblaciones muy grandes. Asimismo, la selección puede actuar en muchos aspectos de la vida de los organismos, siendo la lucha por la sobrevivencia mucho más amplia que un antílope que corre más rápido que un león o un venado con mayores astas que su oponente. La selección puede actuar en procesos como:
• Competencia dentro de la misma especie: por ejemplo, en contiendas por defender un territorio en aves que anidan en colonias.
• Competencia entre especies diferentes: por ejemplo, entre diferentes especies de insectos que se alimentan del néctar de un mismo tipo de flor.
• Atracción sexual: por ejemplo, habilidad de diferentes machos de una población por cortejar a una hembra.
• Fertilidad y fecundidad: por ejemplo, capacidad para criar un mayor número de descendientes.
• Parasitismo: por ejemplo, susceptibilidad y resistencia a los parásitos internos y externos.
De esta forma durante las primeras décadas de nuestro siglo muchos biólogos y naturalistas hicieron importantes contribuciones a la teoría de la evolución, llenando los huecos que Darwin no pudo cubrir y detallando mecanismos, hasta lograr que el neodarwinismo o “síntesis evolutiva”

106
fuera del conjunto de ideas unificador de la biología moderna.
NUEVAS IDEAS SOBRE LA SLECCIÓN NATURAL
Como vemos, el neodarwinismo es un cuerpo teórico aceptado en su mayor parte, sin embargo no ha estado exento de criticas y cuestionamientos.
Posteriormente a su postulación, se dice que la síntesis evolutiva se “endureció”, ya que muchos de sus defensores no permitían considerar, aparte de la selección natural y la adaptación, a ningún otro factor como importante dentro de la evolución orgánica, que consideraban era siempre un proceso gradual. Algunos autores han criticados esta posición y nos advierten sobre la gran importancia de otros factores, como el azar, dentro del proceso evolutivo; mencionan además que este puede darse también en forma brusca, originando no solo nuevas especies sin géneros o familias.
Las mutaciones son modificaciones en la información genética que producen el cambio de un aminoácidos –moléculas que forman las proteínas- por otro en una proteína. Actualmente existen técnicas que nos permiten determinar los tipos de aminoácidos que conforman una proteína y el orden o la secuencia que guardan dentro de ellas: así podemos comparar la secuencia de aminoácidos de una misma proteína en diferentes especies. Realizando este tipo de comparaciones M. Kimura encontró que el orden de los aminoácidos en ciertas regiones de una proteína como la hemoglobina, que se encuentran en la sangre, era distinto en animales, como el caballo, el hombre, el ratón, el tiburón y las gallinas, y que a pesar de estas diferencias la proteína mantenía su función. Estos estudios indican, a diferencia de lo que postulaban los neodarwinistas, que la mayor parte de estos cambios o mutaciones no tiene efectos positivos o negativos sobre el desempeño de los organismos, por lo menos a nivel molecular; Es decir, son mutaciones neutras. Si las mutaciones son neutras, los cambios de un aminoácido por otro no pueden mantenerse por selección natural pues, como ya vimos, ésta sólo puede actuar sobre características que afectan o disminuyen el desempeño de un organismo y no sobre caracteres neutros. Todo esto cuestiona el papel protagonista de la selección natural como fuerza evolutiva a nivel molecular.
Para S. J. Gould y N. Eldredge, el hecho de que en el registro fósil no sea continuo y nos muestre el cambio gradual de las especies, no es resultado de las restringidas condiciones que permiten la fosilización, lo cual ocasión, como lo proponen los neodarwinistas, un registro incompleto, sino de un verdadero proceso de evolución discontinua y brusca que contiene largos periodos de poco cambio en las especies, seguidos por periodos de cambios bruscos que originan especies totalmente diferentes a las ancestrales. Estas ideas ponen en entredicho el carácter gradual de la evolución y la idea que las especies se originan por un proceso lento de acumulación de características favorecidas por la selección natural por lo que aun queda mucho por hacer en el estudio de la evolución.
CUESTIONARIO
1. -EXPLIQUE EN QUE CONSISTE LA SELECCIÓN NATURAL.
2. -EXPLIQUE LA RELACION ENTRE SELECCIÓN NATURAL Y ADAPTACIÓN.
3. -¿POR QUÉ NO PODEMOS HABLAR DE ORGANISMOS MÁS O MENOS ADAPTADOS?
4. -¿CUÁLES SON LAS PRINCIPALES CRÍTICAS AL NEODARWINISMO?

107
PROBLEMAS ASOCIADOS A LA ENSEÑANZA DE LA EVOLUCIÓN EN LA ESCUELA SECUNDARIA:
ALGUNAS SUGERENCIAS FEDRO CARLOS GUILLÉN
INTRODUCCIÓN
Muchos estudios han demostrado que los estudiantes de secundaria tienen problemas para comprender los elementos esenciales de la teoría evolutiva. En este articulo se identifican algunos de estos problemas y se proponen opciones didácticas para la enseñanza de la evolución en la educación secundaria.
GUILLÉN, FEDRO CARLOS (1995), “PROBLEMAS ASOCIADOS A LA ENSEÑANZA DE LA EVOLUCION EN LA ESCUELA SECUNDARIA: ALGUNAS SECUNDARIAS”, EN CIENCIA, VOL. 46, NUM.2.
RESUMEN
En nuestro país, la enseñanza de la evolución en la escuela secundaria ha recibido tradicionalmente un tratamiento marginal. Los enfoques educativos institucionales han planeado modelos que parten de las estructuras individuales y culminan con grandes procesos biológicas. Sin embargo, la Secretaria de Educación Publica modificó este modelo y propuso dentro de los nuevos programas de biología para el ciclo 1993-1994, una estructura en que los procesos evolutivos adquieren muy especial relevancia y presentan como conceptos introductores para la comprensión de los procesos biológicos.
Diversos estudios han demostrado que en los estudiantes del nivel existen problemas para comprender los elementos esenciales de la teoría evolutiva. En este trabajo se caracterizan algunos de estos conceptos relacionados con los temas evolutivos.
I. INTRODUCCIÓN
Hasta 1992, los programas de biología para la educación secundaria elaborados por la Secretaria de Educación Publica (SEP), no reconocieron la importancia de la enseñanza de la evolución como un elemento introductorio a la comprensión de los procesos biológicos. El programa emergente para primer grado que se aplico en el ciclo de 1992-1993, dividía el curso en cinco
unidades: el mundo vivo y la ciencia que lo estudia; células, tejidos y órganos; Funciones biológicas; continuidad y transformación; herencia y evolución, salud humana. Para tratar de caracterizar el peso que recibió cada uno de los temas elegidos, se realizo un análisis de un libro de texto autorizado por la propia SEP: biología 1, de María Batalla Zepeda y Humberto Méndez Ramírez, editado por Kapeluz Mexicana. Varios han sido los autores que señalan la importancia del libro de texto en el ámbito escolar como una fuente primaria de información científica (Carrick, 1982; Hernández, 1994).
En Estados Unidos, nueve de cada diez maestros utilizan el libro de texto el 90% del tiempo (Yager y Penick, 1983 citados por Candela, 1991). El libro de texto permite al investigador realizar análisis puntuales acerca de las características de las propuestas educativas (Osborne y Wittrock, 1985; Novak y Gowin, 1988) y es usado con gran frecuencia por los estudiantes de los primeros cursos de secundaria (Tamir y Amir, 1987), en algunos casos de forma abusiva, obligando al alumno a realizar transcripciones sin sentido educativo (Hardie, 1987).
El análisis de los contenidos del libro elegido arroja resultados dignos de destacarse; de las 279 paginas que se han utilizado para cubrir los temas, 171 (61%) se han destinado a la Unidad II, CÉLULAS, TEJIDOS Y ORGANOS. Encontrarse, el resto de las unidades tiene un número notablemente más reducido (Unidad I, 38; Unidad III, 22 Unidad IV, 32; Unidad V, 16). Si consideramos, como un criterio de la importancia de cada tema el número de horas destinadas a estudiarlo, una lectura de estos datos indicaría que, desde la óptica de los autores y de las autoridades educativas, la estructura de los organismos es el teme prioritario en un curso de biología para estudiantes del primero de secundaria. Esto representa un problema grave ya que los procesos biológicos, precedentes necesarios de cualquier curso, son abordados de manera secundaria y en algunos casos son ignorados.
El tema de la evolución se inscribe dentro de la Unidad IV y es revisado en 3.5 páginas, es decir, el 1% del total del texto. El tratamiento que se le da es poco ordenado e inclusive presenta inexactitudes, como la comparación de la selección natural y las mutaciones como procesos análogos que producen evolución, cuando en realidad, el primero actúa sobre la variabilidad producida por el segundo.
En 1993, en los programas de la SEP se muestra un reconocimiento de la importancia

108
de establecer temas evolutivos como un antecedente necesario en la enseñanza de la biología en la secundaria. La evolución ocupa ahora por completo la segunda unida del primer curso y su antecedente inmediato es la caracterización de la biología como ciencia. Es en este contexto que se hace necesario estudiar y determinar los mecanismos de enseñanza para impartir este tema que permitan sugerir estrategias didácticas adecuadas con el fin de lograr un aprendizaje realmente significativo.
II. LOS PROBLEMAS EN LA ENSEÑANZA DE LA EVOLUCIÓN
A. ¿Porque enseñar evolución en secundaria?
T. Dobzhansky(1993) acuño una frase que refleja con claridad la importancia de la teoría evolutiva en el contexto biológico: “nada tiene sentido en biología si no es la luz de la evolución”.
Actualmente ningún estudioso de la naturaleza del conocimiento biológico cuestiona el papel de la evolución como el cuerpo teórico más global y unificado de la biología. La teoría de la evolución es la pieza fundamental de la biología (Reiss, 1985) y posee importancia científica y social indiscutible. Esta caracterización implicaría necesariamente la inclusión de tópicos evolutivos en el curriculum de los primeros niveles de enseñanza formal de la biología que se presentan en la enseñanza secundaria (Deadman y Kelly, 1978; Reiss, 1985; Engel y Wood, 1985 b). Para realizar el diseño curricular, sin embargo, es muy importante identificar las nociones y preconceptos que los estudiantes manejan sobre un tema. Con el propósito de identificar las ideas que los niños tienen sobre la evolución se han conducido diversas investigaciones (Deadman y Kelly 1978; Brumby, 1979; Engel y Wood, 1985b, Bishop y Anderson, 1990).
Los resultados han seguido una línea: la evolución, entendida como una relación entre los cambios ambientales y la producción de cambios en las características de los organismos mediante mecanismos genéticos, presentan una serie de conceptos que los alumnos encuentran difíciles de asimilar. Los alumnos aparentemente no son capaces de establecer las relaciones que existen entre sus nociones sobre evolución orgánica y las razones ofrecidas por los científicos que les son explicadas en la escuela. El modelo escolar enfatiza frecuentemente el detalle de los conceptos sin poner atención en los
procesos globales. Ante este panorama, existen dos opciones; seguir las ideas de Shayer (1974), quien sugiere que la evolución se enseñe por primera vez en el nivel de preparatoria, dada su aparente complejidad, en claro contraste, atender la propuesta de varios autores(Deadman y Kelly, 1978; Engel y Wood, 1985ª), quienes recomiendan, en lugar de la postergación del tema, la necesidad de instrumentar estrategias para impartirlo más efectivamente. Ante esta disyuntiva es necesario entender que una distinción importante es la que existe entre los temas de curriculum – entendidos como los procesos y criterios para seleccionar y ordenar el conocimiento, las destrezas y actitudes que se enseñan a un grupo determinado- de las estrategias de enseñanza entendidas como la selección de modos de enseñar y propiciar un ambiente de instrucción. En el primer caso, lo importante es la elección de conceptos, en el segundo la elección de actividades con un componente significativo. Esta diferenciación es importante, ya que permite evitar la suposición de que la falla en la estrategia para enseñar un concepto implica necesariamente la imposibilidad de enseñarlo en alumnos de ese nivel (Novak, 1976). En ese sentido me parece que el problema de la enseñanza de la evolución, más que de complejidad conceptual, es de estrategia didáctica. Es necesario entender que la enseñanza de la evolución no solo ofrece ventajas como un concepto estructurador. Enseñar evolución tiene beneficios que no son tan evidentes pero sí de gran importancia; Gougt (1978) sugiere que el estudio de las explicaciones darwinianas sobre la evolución es muy importante en el medio escolar, ya que permitirá comprender la naturaleza misma de la explicación científica. Darwin desarrolló una serie de deducciones ejemplares que comprendían supuesto que podían ser validos de manera empírica. Por ejemplo, el principio de la lucha por la existencia es valido, si se acepta la premisa demostrable del incremento poblacional ante una cantidad limitada de recursos. Enfatizar éste método de razonamiento en el ámbito escolar le permitiría al alumno iniciar la comprensión de principios de razonamiento esenciales que tienen una aplicación concreta en ámbitos no científicos.
Es posible que en lugar de retrasar la presentación del tema sea necesario incluirlo de manera temprana en el curriculum de la secundaria. Algunas evidencias de investigación (Engel y Wood, 1985ª) indican que los alumnos que llegan a la secundaria ala

109
edad de 11 años poseen ya una fuente importante de conocimiento sobre el tema adquirida por medios no-formales de educación, y señalan como un problema el hecho de que los estudiantes no tengan contacto formal con el tema hasta la preparatoria. Estos autores marcan una serie de errores conceptuales comunes que es necesario considerar en el diseño del curso, tales como la idea frecuentemente extendida de que un sexo contribuye más que otro a la transmisión de caracteres o de que las características adquiridas son heredables. Aparentemente una parte significativa de los conceptos biológicos esenciales tiene una forma intuitiva en el pensamiento de los niños. Algunos de estos conceptos pueden permanecer mucho tiempo y afectar la comprensión de los contenidos biológicos que se presentan en la escuela.
B. Las ideas de los niños sobre la evolución
Bonilla y Hernández(1993), en un estudio realizado con alumnos de primero de secundaria, encontraron que la mayoría de los términos fundamentales en los que se estructura la teoría sintética de la evolución no son reconocidos por los alumnos en su connotación biológica. Deadman y Kelly (1978) condujeron un estudio en el que investigaron los conceptos que manejaban los alumnos de secundaria con relación al tema de herencia y evolución. Encontraron que los alumnos reconocían procesos evolutivos únicamente en las poblaciones animales y no eran capaces de establecer las relaciones entre diferentes grupos.
En cuanto a las explicaciones acerca de la ocurrencia del fenómeno evolutivo, todos los niños ofrecieron ideas que pueden ser divididas en dos tipos básicos; naturalistas, en las cuales asocian los cambios con alguna necesidad; asociados a una fuerza interna que impulsa a los animales para ser mejores, o ideas ambientalistas, según las cuales los cambios en los cuales se asocian con cambios en el ambiente. En el momento de explicar como cambian los organismos, los alumnos frecuentemente ofrecieron explicaciones iguales al porque cambian. Sus argumentos tenían un componente lamarckiano en el sentido de que el uso repetido de algún miembro u órgano determinaría una mutación.
En cuanto al concepto de adaptación, prácticamente todos los estudiantes emplearon este termino para fundamentar sus explicaciones de la evolución. La adaptación
fue entendida como la relación entre la estructura del animal y el ambiente. En las explicaciones privó un argumento naturalista. Para los estudiantes la adaptación es resultado de una necesidad del animal. Aparentemente el concepto de preadaptación no les es familiar. Solo aquellos que manifestaron comprensión del valor adaptativo de ciertas estructuras animales incorporaron el concepto de sobrevivencia y erradicaron la idea de cambio por necesidad.
Todos los niños tuvieron claro que algunas especies primitivas se han reproducido mientras otras no; de hecho, emplearon términos como extinción y sobrevivencia, pero sin ligarlos de manera profunda con mecanismos de selección. No se reconoce el carácter intraespecífico de las adaptaciones y todas las explicaciones se refieren a especies diferentes. Algunos niños manejaban un concepto elemental de adaptación y enfatizaron más la sobrevivencia que la extinción.
Los estudiantes no entienden a la evolución como un proceso en el que se incluyan aspectos probabilisticos, y carecen por completo de la información acerca de las fuentes de variación en los organismos. Poseen únicamente la idea de que la herencia es la transmisión de caracteres de una generación a otra. Sus ideas acerca de la aparición de nuevas características se basan en la experiencia. Palabras como gen o cromosoma fueron utilizadas sin que aparentemente los estudiantes comprendieran su significado
Las características de la estructura conceptual sobre la evolución en los niños de secundaria es determinada por Deadman y Kelly (1978) en cuatro apartados:
a) Siete categorías (fenómeno evolutivo, causas de la evolución, mecanismos de la evolución, adaptación, selección, azar, herencia) alrededor de los cuales los estudiantes estructuran sus ideas.
b) El reconocimiento por parte de los alumnos de la importancia de explicar los procesos de evolución y herencia.
c) La comprensión elemental del concepto de herencia y su relación intergeneracional, así como la comprensión de que la evolución implica diferentes animales del pasado y del presente.
d) Las estructuras conceptuales concernientes a la adaptación y la selección

110
Asimismo se sabe que los niños entre seis y 13 años tienen sus propias teorías que tienden a flexibilizarse en la medida que aumenta la edad del estudiante. Una visión bastante común es la idea errónea propuesta por Lamarck, en el siglo pasado, de que los caracteres adquiridos son heredables (Kargbo y cols., 1980). Este hecho resulta interesante, ya que ejemplifica claramente como puede existir una analogía entre las concepciones que estuvieron vigentes en la historia del pensamiento y las percepciones han confirmado que el pensamiento lamarkiano esta presente en estudiantes desde la secundaria hasta el nivel licenciatura (Deadman y Kelly, 1979; Brumby, 1979; Bishop y Anderson, 1990). Angseesing (1978) argumenta que esta tendencia puede deberse al hecho de que la terminología de los libros de texto es confusa e inadecuada y sugiere que se experimente en el laboratorio de manera que se puedan confrontar la teoría de Lamarck con evidencias empíricas.
Un mecanismo para poder comprender el verdadero significado del pensamiento del estudiante es el proporcionarle oportunidades más estructuradas para presentar sus ideas. Uno de los problemas de la enseñanza de la adaptación es que poca gente tiene experiencias propias sobre el proceso. Es necesario reflexionar con el alumno sobre la variación visible de los organismos vivos y enfatizar las causas que lo originan.
Reif (1991) reflexiona sobre el hecho de que la ciencia está deliberadamente organizada para alcanzar metas especiales y es, en varios aspectos, diferente del conocimiento natural de la vida cotidiana; en este sentido frecuentemente los estudiantes no tienen claras las metas del conocimiento científico, lo que dificulta la apropiación de contenidos.
La mayoría de los estudiantes “cree” en la evolución debido al prestigio de la ciencia que avala la teoría más que avala la teoría más que en el entendimiento y razonamiento e la misma (Lucas, 1987). Por otro lado, la mayoría de la gente no parece entender el proceso evolutivo como ha sido descrito por los científicos, aun después de haberlo estudiado (Bishop y Anderson, 1990). En un trabajo realizado en Inglaterra para conocer el desempeño de los adultos con antecedentes de estudios científicos, contra los que no tenían, en cuanto biología elemental, se encontró que no había una diferencia significativa, lo que sugiere el poco efecto del trabajo escolar en los estudiantes. Estos autores encontraron además diferencias entre las ideas científicas y las que los alumnos
manejan. Una de las características de la teoría evolutiva es la distinción de dos procesos; por un lado, la aparición aleatoria de cambios en la estructura genética de una población, en función de mutaciones o recombinación genética, y por otro la sobrevivencia o extinción diferencial de los individuos en función de presiones ambientales (selección natural). En general la mayoría de los estudiantes no son capaces de reconocer la diferencia entre estos dos procesos, que entienden como uno solo.
Los estudiantes piensan que el ambiente causa los cambios en las poblaciones a través del tiempo. Los mecanismos que sugieren son de necesidad (el organismo “necesita” correr más rápido), de uso y desuso (el no uso de los ojos los convierte en disfuncionales), de adaptación (el color blanco del color de la piel de los osos ha cambiado como resultado de las presiones ambientales).
Estas implicaciones tienen un componente lamarckiano; esto se debe a la dificultad ya mencionada de separar los procesos aleatorios de mutación de los no aleatorios de selección, para los estudiantes basta la explicación de una función que frecuentemente confunden con el mecanismo evolutivo.
La variación en un componente esencial de la teoría evolutiva. En sentido estricto es el sustrato sobre el que actúa la selección natural. En los estudiantes ésta no es una noción clara, y entienden a la evolución como un proceso que homogeniza a las especies.
Las nuevas características se diseminan en una población debido a que los organismos que las poseen se reproducen con mayor frecuencia. Los estudiantes piensan que estos cambios se van dando en las mismas características de manera gradual entre una generación y otra.
El concepto de adaptación es entendido en su aceptación cotidiana, que es diferente de la que se utiliza en el contexto evolutivo. Los biólogos utilizan el termino adaptación refiriéndose a un fenómeno poblacional, donde los cambios se producen, a través de varias generaciones, debido a la acción de la selección natural. Los estudiantes interpretan el concepto de adaptación como un termino que se refiere a cambios individuales por medio de un esfuerzo propio, como cuando un perro se adapta a su nueva casa; en el momento que los alumnos escuchan en la escuela el termino adaptación, que se les presenta en un contexto evolutivo, inmediatamente lo refieren a su propia concepción, lo que tiende a reforzar

111
concepciones equivocadas de carácter naturalista (Bishop y Anderson, 1990). Estas ideas de los alumnos aparentemente se pueden modificar si los maestros las conocen y diseñan metodos para enfrentarlas.
Uno de los argumentos de los docentes para explicar su resistencia a impartir el tema, se basa en que la evolución no es una materia de carácter practico. Sin embargo, existen trabajos que atendiendo a este problema se han desarrollado para producir practicas viables en el ámbito escolar sobre selección natural (Allen y cols.), 1987) y sobre selección sexual (Adams y Greenwood, 1987).
Seria necesario que los docentes aplicaran cierta iniciativa para reproducir estas experiencias en el salón de clase.
Como una estrategia para transmitir adecuadamente el concepto de evolución, éste se debe tratar de entender de una manera cabal, determinando su reorganización jerárquica; es así como el docente contara con un elemento estructurado para transmitir los conceptos que se deriven de él.
Otros estudios han demostrado que los alumnos de la secundaria poseen una visión teleológica y antropocéntrica del concepto de adaptación biológica (Engel y Wood, 1985b). La presencia del pensamiento teleológico puede explicarse por el uso en clase o en libros de texto de frases como “el mejor adaptado” o “la supervivencia del más apto”, que intuitivamente transmiten una idea de mejoría en las poblaciones. Es necesario enfatizar el hecho de que esta mejoría es tan variable como las presiones de selección que actúan sobre los organismos. Es decir, como el ambiente es variable, no es posible conseguir un “producto acabado” en términos evolutivos, ya que las condiciones en que es apto pueden variar y determinar que sus características se vuelvan ineficaces para enfrentar las presiones ambientales. En un estudio, Jungwuith (1975) demostró que una proporción elevada de alumnos de secundaria acepta conceptos de adaptación y evolución desde las perspectivas antropocéntrica y teleológica. Esta aceptación es literal y no de manera metafórica lo que distorsiona su visión de los conceptos evolutivos.
III. ALGUNAS SUGERENCIAS.
Es necesario que los maestros posean una visión muy clara de las ideas que los alumnos tienen sobre la evolución. El siguiente temario representa el trabajo de varios autores y nos ofrece una línea de resultados que, creemos,
es necesario considerar para determinar las categorías docentes:
Es importante sondear la información que el alumno maneja, de tal manera que pueda contrastar las diferencias conceptuales con respecto a la información aceptada científicamente. En el caso de la evolución podemos suponer que:
El sentido común es lamarckiano. Los niños piensan que en los animales existe un deseo de mejorar y, en general, los caracteres adquiridos son heredables.
No se maneja el concepto de variabilidad.
No existe un concepto de mutación en los estudiantes que les permita explicarse la variabilidad.
Se piensa que la evolución ocurre a lo largo de la vida de un organismo.
El concepto de adaptación se interpreta en su acepción cotidiana, como un elemento positivo que ayuda a los animales a sobrevivir.
No existe la concepción de que la evolución ocurre también en los vegetales.
No se diferencian las fases del proceso evolutivo: la producción aleatoria de variabilidad y la selección no aleatoria de los organismos más aptos.
Conceptos como gen o cromosomas son manejados en el vocabulario dl alumno sin que se entienda su significado.
Se piensa que el uso o desuso favorece la aparición o desaparición de caracteres.
Se confunden las explicaciones de cómo y porque ocurre la evolución.
Existe una visión teleológica y antropocéntrica d la evolución.
Se piensa que los cambios ambientales determinan los cambios en las estructuras de los animales.
Se puede sugerir una posible explicación, dividida en dos líneas, del porque de los problemas de los niños para entender conceptos evolutivos:
a) Es más sencillo para la estructura conceptual del niño entender ciertos conceptos de una manera, aunque estos no tengan una validación científica. Intuitivamente, por ejemplo, es más fácil pensar que los organismos adquieren los elementos necesarios para vivir y los heredan, aunque esto no sea cierto.

112
b) Los medios de educación no formal, en un esfuerzo por simplificar las ideas, llevan a confusiones e interpretaciones equivocadas. “Solo los animales más fuertes o inteligentes sobreviven” es un ejemplo de lo que se menciona en un programa de televisión sobre evolución (Bishop y Anderson, 1990) o “el hombre viene del mono”, frase atribuida a Darwin que supuestamente resume su teoría (Guillen, 1992). Existen modelos cognitivos con base en el sentido común que son muy resistentes al reemplazo por modelos validados científicamente (Greene, 1990). La enseñanza del desarrollo histórico del pensamiento evolutivo es muy importante, ya que de esta manera pueden darse cuenta de la forma en que los conceptos similares a los que poseen se modificaron por la aparición de nueva evidencia (Engel y Wood, 1985ª).
A continuación presentare una serie de sugerencias puntuales para la enseñanza de la evolución en la secundaria.
• Es muy importante que se abunde en el diseño de actividades experimentales que permitan al alumno reconocer principios evolutivos básicos, como selección natural o herencia de caracteres adquiridos. Tradicionalmente la evolución ha sido catalogada como un cuerpo teórico que no puede demostrarse experimentalmente. W.J. Crozier explicaba en 1930 a sus alumnos de Harvard en su curso introductorio a la biología: “La evolución no es ciencia: ustedes no pueden experimentar con dos millones de años” (Smocovitis, 1992). En realidad este prejuicio aun existe en el ámbito escolar: frecuentemente los maestros piensan que no es posible contar con apoyo experimental para sustentar el tema. Sin embargo, existen muy diversas fuentes de experimentación que pueden demostrar principios evolutivos elementales, por ejemplo, sobre selección natural (Allen y cols., 1987) y selección sexual (Adams y Greenwood, 1987).
• Es necesario que los maestros puedan diseñar situaciones problema que le permite al alumno plantearse explicaciones acerca de los mecanismos de cambio. Angseesing (1978) sugiere que una de las formas que el alumno pueden erradicar el pensamiento de tipo Lamarckiano es mediante de la solución de problemas que le permitan detectar las fallas de la teoría propuesta por Lamarck,
por ejemplo: la polilla Biston betularia se presenta en dos variedades con colores distintos: claro y oscuro. En una zona urbana el 90% de los individuos son oscuros mientras que en una zona rural son menos del 10%. Se pueden ofrecer cuatro hipótesis:
a) El color se determina genéticamente y las diferentes frecuencias de las dos formas en las dos áreas se deben a que varían en su habilidad para sobrevivir en cada ambiente.
b) El color es determinado genéticamente y las diferentes frecuencias de las dos formas se deben al azar.
c) Cada polilla tiene habilidad de cambiar de color para parecerse a su ambiente
d) El color es determinado genéticamente y las diferentes frecuencias de las dos formas en las dos áreas se deben a que cada polilla busca un fondo que no contraste con su propio color.
Un Biólogo decide distinguir entre estas hipótesis liberando un número determinado de polillas marcadas para luego recapturarlas.
a) ¿Es necesario que libere polillas de ambos colores?
b) Si lo hace, ¿debería marcar a los dos tipos de polillas de manera diferente?
c) ¿Debería liberar polillas en las dos localidades?
d) ¿Existe alguna ventaja en liberar sólo polillas oscuras en un ambiente claro?
* La conceptualización del proceso mediante algunas herramientas como mapas conceptuales (MC) presentan relaciones significativas entre proposiciones que se articulan a través de unidades semánticas de manera esquemática. Los MC permiten llamar la atención sobre las ideas más importantes en un proposición, así como las relaciones entre los significados de los conceptos. En nuestro caso, parten de la premisa de que la biología puede ser enfocada como un sistema conceptual.
Los MC deben ser jerárquicos, con los conceptos más generales en la parte superior y los conceptos más específicos en la parte inferior. Un mismo concepto puede ser manejado de diferentes maneras en distintos MC.

113
Los MC le permiten al alumno reconocer las conexiones entre los diferentes tópicos del conocimiento. El profesor los puede utilizar para determinar las rutas que se siguen para organizar los significados y además deslindar la información relevante de la trivial. Un aspecto muy importante es que los MC son un instrumento de evaluación de la significatividad del aprendizaje, aspecto muy difícil de medir valiéndose de otras técnicas. En particular, el análisis conceptual de un área del conocimiento biológico es un ejercicio muy importante a través de esta técnica (Okeke y Wood, 1980). Un estudio realizado por Okebukola (1990), demostró que la aplicación de la técnica de mapas conceptuales mejoraba significativamente el aprendizaje de varios conceptos de la genética y de ecología en una población de estudiantes con edades entre los 15 y 16 años. Un posible ejemplo se sugiere en la figura A.
• Se debe ilustrar por medio de ejemplos que la explicación lamarckiana n funciona, en lugar de pedirles a los alumnos que acepten las ideas del profesor. Frecuentemente los maestros confían en que el impartir un tema los alumnos tomaran estas ideas, aun cuando se encuentren en conflicto con sus propias explicaciones. En muchos casos el alumno “negocia” y le ofrece a su maestro la explicación que éste demanda, sin que esto implique, necesariamente la aceptación de sus ideas.
Para evitar este problema los maestros deben ofrecer, además de una explicación, ejemplos significativos y comprensibles para los estudiantes.
IV: CONCLUSIÓN
En el contexto de la reforma a los programas de biología, resulta fundamental el análisis de las ideas que los niños manejan antes de tener contacto formal con explicaciones científicas. A partir de este análisis se deberán diseñar instrumentos didácticos que permitan incidir en estas ideas y aprovecharlas para un aprendizaje efectivo y duradero.
Resulta claro que las dificultades que tradicionalmente ha presentado la transmisión de conceptos científicos en la escuela, solo podrán superarse en la medida que seamos capaces de transformar las estrategias educativas tradicionales, que han privilegiado el enciclopedismo y la información, en otras nuevas que tomen en cuenta la estructura del conocimiento de los estudiantes y les
proporcionen elementos con significados sociales y personales.
La evolución es sin duda el concepto más global y unificador en biología y así lo han reconocido los nuevos programas elaborados por la SEP.
Es necesario, en consecuencia, que los maestros cuenten con elementos que le permitan una transmisión más eficaz de este tema.
CUESTIONARIO
1.- ¿CUÁL ES LA IMPORTANCIA DE ENSEÑAR EVOLUCIÓN EN LA ESCUELA SACUNDARIA?
2.- ¿CUÁLES SON LAS CONCEPCONES ERRÓNEAS MÁS FRECUENTES EN TORNO A LA EVOLUCIÓN BIOLÓGICA?
3.- ¿CUÁLES SON LAS PRINCIPALES RAZONES QUE LIMITAN EL APRENDIZAJE DE LA EVOLUCIÓN?
4.- ¿QUÉ TIPO DE ESTRETEGIAS DIDÁCTICAS SE REQUIEREN PARA FACILITAR EL APRENDIZAJE DE LA TEORÍA EVOLUTIVA?

114
DARWIN Y WALLACE
(FRAGMENTO)
FEDRO CARLOS GUILLÉN
INTRODUCCIÓN
La ciencia es una actividad realizada por seres humanos por lo tanto en su historia encontramos una gran cantidad d hechos tan cotidianos como los miedos, angustias y malos entendidos de nuestras propias vidas. En este texto, donde se narra la preocupación de Darwin al conocer que alguien más compartía sus ideas sobre la evolución, encontramos un ejemplo de ello.
GUILLÉN, FEDRO C. (1994), “LAS CARTAS MARCADAS: DARWIN Y WALLACE”, EN LA JORNADA SEMANAL, NÚM. 256, PP.26-30.
El 24 de noviembre de 1859, Darwin publico su teoría evolutiva en el libro que todos conocemos como El origen de las especies. La edición consto de 1250 ejemplares que se agotaron el mismo día pese a que Darwin, en una carta que envío a Charles Lyell en septiembre de ese año, decía ¿Con ingenuidad?: “Murray ha impreso 1250 copias, lo cual me parece una edición demasiado voluminosa, pero espero que no sufra perdidas”. De tal evento, que sentó las bases de la biología moderna se pueden desprender dos hechos: a) ¿cómo demonios se agoto la edición en un solo día?, y b) Darwin tenia la modestia de una señorita victoriana. Un tercer hecho, que me dejo muy impresionado lo explico mi maestra de evolución, la popular Berthita, una mujer que considerando su edad, probablemente fue testigo de los hechos. Al iniciar su explicación nos dijo suspirando como Margarita Gauthier: “hay muchachos, y pensar que todo empezó con una carta...”
La carta a la que se refería la maestra Berthita fue recibida por Charles Darwin el viernes 18 de junio de 1858, el remitente era Alfred Russel Wallace un joven naturalista que trabajaba en el archipiélago Malayo. Después de una aguda crisis de fiebre, Wallace se había levantado de la cama para escribir un pequeño ensayo al que llamó On the tendency of varieties to depart indefinitely from the iriginal type (Sobre la tendencia de las variedades a diferenciarse indefinidamente del tipo original): al terminar su trabajo Wallace escribió una carta a Darwin, en la que pedía sum opinión y la consideración de que hiciera
llegar, si lo creía adecuado, su trabajo a Charles Lyell, eminente geólogo amigo de Darwin. La carta enviada el 12 de marzo tardo (eran otros tiempos) 99 días en llegar a Inglaterra y su impacto fue brutal; Charles Darwin escribió el mismo día que la recibió una nota para Lyell que revelaba su consternación: “Sus palabras han resultado ciertas, con el agravante que se me han adelantado”.
¿En que se le habían adelantado? Pues en la primicia de la teoría evolutiva, Wallace exponía un resumen de las ideas que le había tomado más de 20 años elaborar a Darwin, quien por cautela no las había hecho publicas. Nos encontrábamos con el escenario de un drama perfecto; por un lado, un hombre de 49 años (Darwin) acumulando las evidencias de una vida para proponer la teoría más revolucionaria desde que Newton transformo el pensamiento científico. Por otro, un joven (Wallace) que había tenido un chispazo genial y lo daba a conocer por medio de la dichosa carta.
La prioridad de las ideas efectivamente correspondía a Darwin quien llevaba muchos años pensando sobre el asunto. El 14 de mayo de 1856 había iniciado un libro en el que explicaría su teoría, que fue interrumpido en junio de 1858 cuando recibió la carta de marras. Sin embargo, ese no era el punto; Wallace sin conocimiento de la teoría Darwiniana si había transmitido sus ideas y en consecuencia podría: a) asumir la paternidad de la teoría, o b) confiar en una respuesta que con tres gotas de mala intención podría ser interpretada como un plagio desvergonzado: “¿Qué cree? Lo que usted me expone ya l había pensado”.
Desde luego si yo hubiera sido Wallace, ante la pobre evidencia le hubiera metido una demanda a Darwin que todavía estaría pagando sus herederos pero no es el caso; ni soy inglés, ni tengo 25 años, y mucho menos soy un caballero.
Por alguna razón, que espero que usted comparta querido lector, analizar como se desarrollaron los hechos me parece fascinante y para ello confiaremos en las evidencias históricas.
Veamos:
El 1º de mayo d 1857, Darwin escribió a Wallace: “Es realmente imposible explicar mis teorías(en la extensión de una carta), sobre las causas y medios de variación en estado de naturaleza”. La demostración de que se

115
equivocaba la recibió por parte de Wallace dos años después.
Es decir, era perfectamente posible explicar la teoría de una carta.
El 18 de junio de 1858, día que recibe la comunicación de Wallace, en una amarga carta a Lyell, Darwin tira la toalla en el primer round:
(Nunca he visto una coincidencia más sorprendente. ¿Si Wallace tuviera la copia de mi esquema hecha en 1842 no podría haberlo resumido mejor! Sus mismos términos son ahora los títulos de mis capítulos. Por favor devuélvame el manuscrito; el no ha demostrado su deseo de que yo lo publique, pero naturalmente voy a escribir ofreciéndolo a alguna revista. De este modo mi originalidad, cualquiera que sea, va a quedar destruida, pero mi libro si es que tiene algún valor, no sufrirá deterioro, ya que todo el trabajo consiste en la aplicación de la teoría. Espero que de el visto bueno al esquema de Wallace para poder comunicarle su opinión).
Esto quería decir que Darwin renunciaba a la paternidad, cumplía las indicaciones de Wallace y esperaba los comentarios de Lyell. Sin embargo, para el buen Charles el golpe era demasiado fuerte. Su correspondencia de los siguientes días nos demuestra a un hombre atormentado por su enorme deseo de no perder el derecho sobre sus ideas en esquizofrénico conflicto con su honestidad victoriana. Para curarse en salud, Darwin hizo algo que le era muy común: pidió consejo. En solo ocho días su renuncia se convirtió en la claridad de que la teoría era originalmente suya. El 25 de junio de 1858, le escribió nuevamente a Lyell:
“No hay nada en el esquema de Wallace que no este mucho más completo en el mío, que copie en 1844... envié un breve boceto, del que conservo una copia de mis teorías, a Asa Gray, de modo que podría con toda exactitud decir y probar que no he tomado nada de Wallace. Me gustaría muchisimo publicar ahora un resumen de mis teorías generales pero no logro convencerme de que puedo hacerlo honradamente. Wallace no dice nada de publicarlo, le adjunto su carta. Pero como yo no había pensado sacar a la luz resumen alguno, ¿Puedo hacerlo honradamente aunque Wallace me haya enviado un esquema de su doctrina? Preferiría quemar mi libro entero antes de que él u otro cualquiera pensara que ha obrado indignamente.
¿No cree que el hecho del que él me haya enviado el esquema me ata las manos?... si
pudiera honradamente publicarlo haría constar lo que me induce a publicar ahora el esbozo.
Me gustaría enviar a Wallace una copia de mi carta a Asa Gray para demostrarle que no le he robado su teoría. A propósito, ¿tendría inconveniente en enviar ésta con su respuesta a Hooker y que él a su vez la envié a mí? Porque así tendré la opinión de mis dos mejores y más comprensivos amigos. He escrito esta carta lleno de tristeza, y lo hago ahora para poder olvidarme de este asunto por algún tiempo; estoy agotado de tanto meditar... no lo volveré a molestar ni a Hooker ni a usted acerca de este asunto.
En buen español, Darwin recurrió a la famosísima frase “te digo Chana para que me entiendas Juana”. No quería parecer un abusivo, pero tampoco ceder sus derechos. Por supuesto no le escribió a Wallace (que hubiera sido correcto), sino a dos amigos cuya incondicionalidad hacia él era manifiesta. Seguramente pasó muy mala noche ya que al día siguiente, 26 de junio de 1858, y pese a la advertencia de no volver a molestar con la historia de la carta, molesto a Lyell enviándole una nota:
Querido Lyell: perdóneme que añada una posdata que refuerce en lo posible los argumentos contra mí. Wallace podría decir: “no pensó usted en publicar un estrato de sus teorías hasta que recibió mi comunicación. ¿Es justo que se aproveche de que yo, libremente y si que usted me lo pidiera, le informara de mis ideas, y que impida de ese modo que yo adelante?”. Si publicara inducido por el hecho de saber privadamente que Wallace está en la misma línea, seria un abuso. Se me hace duro verme así obligado a perder mi prioridad de muchos años, pero no estoy del todo seguro que eso altere la justicia del caso. La primera impresión es la que vale, generalmente, y lo primero que yo pense fue que no seria honrado publicar ahora.
¡Bingo! Darwin planteaba todas las objeciones que cualquier persona razonable encontraría en el hecho de que publicara sus ideas. Sin embargo, en lugar de aceptar estas objeciones, siguió escribiendo cartas. El 29 de junio, en respuesta a una misiva de Joseph Hooker, señalo:
Acabo de leer su carta y veo que quiere los artículos en seguida, le envío el de Wallace y el resumen de mi cata a Asa Gray, que expone muy imperfectamente sólo los medios de cambio, y no alude a las razones que apoyan la convicción de que las especies cambian efectivamente. Me atrevería a decir que es demasiado tarde. Ya apenas me

116
preocupa, le envío mi esquema de 1848 solo para que pueda ver de su propio puño y letra que lo leyó. Ya no puedo soportar verlo. No le dedique demasiado tiempo. Es una bajeza por mi parte preocuparme de la prioridad.
Por supuesto, es una carta muy rara, Darwin recuerda al personaje de las películas que grita que no esta triste mientras se limpia las lagrimas. Por otro lado encuentra que su carta a Asa Gray, que él veía como la evidencia de su anticipación, no es tan completa como parecía. Los hechos indican que realmente no había nada que hacer.
Y sin embargo, se hizo.
El 1º de julio de 1858, apenas dos días después, Charles Lyell y Joseph Hooker se presentaron ante la Linnean Society para leer una memoria del trabajo Darwin-Wallace en el que se incluían párrafos del resumen de 1844 y parte de una carta de 1857 dirigida a Asa Gray. El aparente acuerdo era (en versión de Lyell y Hooker) el que sigue:
Darwin estimaba que el valor de las teorías que se exponían en el trabajo de Wallace era tan grande que le propuso a Lyell que obtuviera su permiso para publicar dicho trabajo. Así se convino con la condición de que Darwin no ocultara del publico, como quería, la memoria que él había escrito sobre el mismo tema, que uno de nosotros había leído en 1844, y cuyo contenido habíamos conocido ambos en secreto durante muchos años. Cuando expusimos esto a Mr. Darwin. Él nos dio permiso para utilizar su memoria como consideráramos conveniente.
Después de leer lo anterior cualquiera podría pensar que:
a) Darwin le pidió a Lyell que publicara la obra de Wallace.
b) Darwin quería ocultar del publico su manuscrito de 1844.
c) Lyell tuvo que convencer a Darwin para que su trabajo fuera presentado.
Todo lo anterior, me parece no tiene nada que ver con la realidad. Darwin hubiera dado un brazo por encontrar una salida honorable que le permitiera mantener la prioridad de sus ideas. La lectura y posterior publicación del trabajo “conjunto” fue la solución.
Darwin concluye en una carta que dirige a Hooker, el 13 de julio de 1858, su alivio e evidente:
Su carta a Wallace me parece perfecta, clarisima y sumamente cortas. No creo que
pudiera mejorla y la he enviado hoy con otra mía. Siempre pense que era muy posible que alguien se me adelantara, pero imaginaba que tenia un alma lo suficientemente grande para no preocuparme por ello; ahora me encuentro equivocado y castigado; sin embargo, me había resignado totalmente, y ya tenia media carta escrita a Wallace cediéndole toda prioridad: no habría cambiado, ciertamente, si no hubiera sido por su extraordinaria amabilidad, la de Lyell y la suya. Le aseguro que soy consciente de ella y que no la olvidare. Estoy más satisfecho por lo que ocurrió en la Linnean Society. Había supuesto que su carta y la mía a Asa Gray no serian más que un apéndice al trabajo de Wallace...
El 18 de julio le escribe a Lyell:
Nunca le he agradecido lo bastante los extraordinarios esfuerzos y amabilidad que ha demostrado conmigo en el asunto de Wallace. Hooker me dijo que lo hicieron en la Linnean Society y estoy contentisimo; no creo que Wallace pueda considerar mi conducta desleal, si permito que usted y Hooker hagan lo que consideran justo...
Darwin se maneja como un hombre a punto de ser lo correcto que es convencido a la ultima hora. El argumento final que utiliza es lamentable; al deslindarse de su responsabilidad como el destinatario de la carta de Wallace, Darwin muestra nuevamente esta actitud de “ya que insisten” que mantuvo durante toda el proceso.
Wallace jamás fue consultado sobre esta decisión, y sin embargo, se comporto como un perfecto caballero y siempre reconoció la prioridad de Darwin. El 3 de diciembre de 1887 (ya fallecido Darwin) escribió una carta A. Newton que refleja su visión del asunto:
...En aquel tiempo y no tenia n la más remota idea de que él había llegado ya una teoría definida, y aún menos de que esta era la misma que a mí se me había ocurrido de repente en Ternate, en 1858...
No es que hubiera pensado en morirme (wallace había pasado por un ataque de fiebre) pero si pensaba en desarrollarla (la teoría) todo lo posible, cuando volví a casa sin suponer en absoluto que Darwin se me había adelantado tanto. Puedo decir con toda verdad ahora, como dije hace muchos años, que me alegro de que fuera así; porque yo no siento el amor por el trabajo, por la experimentación y el detalle que eran tan preeminentes en Darwin y sin el cual nada de lo yo pudiera haber escrito habría convencido al mundo...

117
Paradójicamente la respuesta de la Linnean Society ante el comunicado fue gélida, no hubo cuestionamiento ni polémicas series. Pero eso no preocupó a Darwin. A finales de julio inicio la redacción de El origen de las especies. En el primer párrafo hizo referencia al comunicado de Wallace. El 24 de noviembre de 1859, el libro se puso a la venta y los 1250 ejemplares volaron el mismo día, asunto que me parece un misterio. ¿Había acaso 1250 personas ávidas de leer la teoría sin poder esperar al día siguiente? Aparentemente sí. El libro completó seis ediciones y se constituyó sin duda alguna en la piedra fundamental sobre la que descansa el conocimiento biológico moderno.
Charles Darwin murió en 1882 a los 74 años. Fue enterrado en la abadía de Westminster, muy cerca de la tumba de Isaac Newton.
Me parece necesario concluir que si la participación de Darwin en el caso de Wallace dejó mucho que desear, ello en nada demerita su espléndida contribución al pensamiento biológico. Charles Darwin demostró que una mente sistemática y genial es capaz de vencer el desafío más notable –pese a rendirse eventualmente ante las debilidades tan propias de la condición humana.
NOTAS
1.- Las cartas que se citan en este texto se encuentran en Charles Darwin: autobiografía y cartas escogidas, 2, selección de Francis Darwin, Alianza Editorial, Madrid, 1984.
2.- las cursivas que aparecen en las cartas son mías.
CUESTIONARIO
1.- ¿qué hubiera hecho si fuera Darwin? ¿Cree que resolvió bien el problema?
2.- ¿Cree la condición de científico libera a las personas de los problemas y conflictos de cualquier ser humano?
3.- ¿Por qué cree que existe el mito de los científicos como personas alejadas de la realidad, que viven aisladas en sus laboratorios? ¿Cómo podemos modificar esta concepción en nuestros alumnos?

118
LAS MUSAS DE DARWIN (FRAGMENTO)
JOSÉ SARUKHÁN
INTRODUCCIÓN
En este fragmento de su libro Las musas de Darwin, dedicado a relatar la historia del nacimiento de la teoría de la evolución de Charles Darwin, el doctor José Sarukhán, distinguido ecólogo mexicano, presenta una explicación difícil de superar, por su brevedad, concisión y exactitud, de los conceptos fundamentales de dicha teoría.
SARUKHÁN, JOSÉ (1989), LAS MUSAS DE DARWIN, FCE (LA CIENCIA DESDE MÉXICO), PP.254-260 Y 283-288.
LOS PILARES DE LA SELECCIÓN NATURAL
Darwin se basó en cuatro elementos centrales que son, en forma muy cercana, el resultado de la influencia de las musas a las que me he referido en este libro. Como ya también lo mencioné, esos elementos no tuvieron una secuencia especial, ni creo que uno de ellos tenga notablemente más peso que los otros. El orden en el que me referiré a ellos es el que me parece que puede haber ocurrido cronológicamente en el desarrollo de las ideas de Darwin.
1.- La variabilidad individual y su potencialidad.
Todos los miembros de una especie difieren entre sí. Aunque no las podamos detectar fácilmente en muchas especies, las diferencias entre los individuos son tan marcadas como las que podemos reconocer de inmediato entre los integrantes de la especie humana. Esas diferencias individuales, como también ocurre en el hombre, no solamente se refieren a características morfológicas, sino también a aspectos fisiológicos y de comportamiento. Sin embargo, esa variabilidad morfológica, fisiológica y de comportamiento puede tener otro origen. Una fuente de variabilidad es intrínseca al individuo, es decir, tiene origen genético y por lo tanto puede transmitirse a los descendientes. Otra fuente es la influencia de los factores del ambiente, tales como la temperatura, las características del alimento, la educación en el caso del hombre, etc., esta variabilidad de origen ambiental no es heredable.
Aunque estas dos fuentes de determinación de atributos de los individuos están bien establecidas, determinar en que proporción
son causantes de las características de los individuos ha sido uno de los problemas más añejos y difíciles de resolver en la biología. La heredabilidad de un atributo requiere que se conozca la variabilidad total del mismo en toda la especie para determinar, por medio de laborioso experimentos de cruzamiento y por observación, qué tanto de dicha variabilidad es heredada y qué tanto está causada por el ambiente. Lo anterior es menos difícil cuando se trata de atributos o características sencillas, como las que Mendel observó en los chícharos, (con los que definió sus leyes de la herencia) o como las características de los grupos sanguíneos humanos. El problema se torna en extremo difícil cuando se intenta establecer en que proporción la herencia o el ambiente determinan características mucho más complejas como la capacidad reproductora de los individuos de una especie o la inteligencia en el hombre. Es necesario aclarar que además de la variabilidad individual, se pueden distinguir dos categorías más de variación dentro de una especie: la variación geográfica y el polimorfismo. La primera implica cambios mucho más notables que sutiles y diferencias entre los individuos de una población y ocurre por lo general entre individuos de la misma especie, pertenecientes a regiones geográficas diferentes. Un ejemplo bien conocido de dicha variabilidad son las diferencias entre razas humanas típicas de diferentes regiones del planeta, que aún podemos reconocer a pesar de la considerable mezcla racial que existe en la actualidad. A diferencia de la variabilidad entre los individuos de una población, la cual ocurre preponderantemente al azar, la variabilidad geográfica tiene por lo general valores de tipo adaptativo, ya sea el clima u otros factores ambientales.
Usando el nuevo ejemplo de las razas humanas, la pigmentación de la piel en las razas africanas es una protección contra el exceso de radiación solar de las latitudes ecuatoriales, mientras que en latitudes más al norte, la menor incidencia de radiación solar favorece la posición de pieles mucho menos pigmentadas, las cuales dejan pasar más radiación ultravioleta, que es en buena parte causante de la fijación de la vitamina D. Desde luego, esta variabilidad geográfica ocurre en todas las especies, animales o plantas, que tienen una distribución geográfica lo suficientemente amplia como para que en ellas se presenten variaciones ambientales.
El polimorfismo es un fenómeno que ocurre en una misma área y, como su nombre lo

119
denota, se refiere a la existencia de varias formas de la misma especie en el mismo lugar. Esta variación puede expresarse en características morfológicas externas muy notorias como la coloración o el tamaño del cuerpo, o bien en características detectables sólo por medios indirectos. Entre el primer grupo podemos citar las grandes diferencias que existen entre los machos y las hembras de muchos animales (aves o insectos, por ejemplo) o bien entre las hembras de muchas especies de mariposas, como es el caso de las hembras de la mariposa monarca, Danaus plexippus: este marcada polimorfismo de las hembras de los lepidópteros ha sido seleccionado como un mecanismo de protección contra la depredación por aves, ya que al copiar varios modelos de mariposas de sabor desagradable, pueden engañar más fácilmente a sus depredadores y por lo tanto tienen mayores probabilidades de sobrevivir. Un ejemplo de polimorfismo oculto muy bien conocido por nosotros es el caso de los tipos sanguíneos presentes en nuestra especie, los cuales solamente pueden reconocerse por medio de una química en la sangre. Cada uno de nosotros pertenece a uno de los cuatro tipos básicos existentes (A, B, AB, y O) y no se presentan casos intermedios
Los anteriores ejemplos de las muchas variantes que ocurren en la naturaleza. No obstante, la variabilidad que primeramente llamó la atención de Darwin fue la que expresa como resultado de la acción del hombre en el proceso de domesticación de animales y plantas. Pero probablemente lo que le impresionó más fue el enorme potencial que dicha variabilidad tenia para expresarse, como resultado de un proceso selectivo del hombre. Darwin tuvo amplia oportunidad para observar y estudiar tanto la variación resultante del proceso de domesticación como la encontró en la naturaleza durante su viaje alrededor del mundo, en especial en las islas Galápagos.
2. -La transmisión a la progenie de las características variables.
La variabilidad observada entre los individuos puede tener causas exógenas (el ambiente) o endógenas (las características genéticas). Darwin no tenia conocimiento de la forma en que la variabilidad heredada era transmitida a los descendientes, aunque le era evidente que si las características favorables de los padres seleccionados por el ambiente no eran transmitidas a sus descendientes, no habría forma de que ocurriesen cambios en los
atributos de la población y por lo tanto no ocurriría una transformación, o evolución, en las especies. La selección bajo la influencia de la domesticación lo había convencido (correctamente) de ello; solo que ignoraba el mecanismo por el que operaba la herencia. El problema le interesaba de manera especial e incluso... llegó a proponer, en forma errónea, mecanismos por los cuales ahora llamamos la información genética podría ser transmitido de padres a hijos. Los mecanismos de la herencia no fueron conocidos en forma cabal hasta el redescubrimiento de la obra de Gregor Mendel a principios del actual siglo cruciales no solo para explicar muchos aspectos que Darwin no pudo entender por falta de información, sino también para propiciar un avance en el pensamiento darwiniano.
3. - las especies dejan más de la que es posible que sobreviva.
Como resultado de la capacidad potencial de crecimiento exponencial de las especies, éstas generan un número muy superior de individuos de los que normalmente podrían establecerse. Por ejemplo, los abetos de un solo bosque requieren que una sola plantula logre establecerse para reponer cada uno de los individuos presentes; sin embargo, cada árbol de abeto puede producir cada año decenas de miles de semillas, de las cuales es probable que no llegue a establecerse, como plantula, una sola. ¿Por qué este aparente desperdicio de energía y recursos? En realidad no se trata de un desperdicio, sino de un mecanismo de seguridad por parte de los organismos ante lo imprevisible y variable de los factores ambientales causantes de los factores ambientales causantes de la mortalidad de la progenie. Las causas de selección natural han favorecido los comportamientos de los individuos que dejan progenies muy elevadas, con objeto de que se incrementen las probabilidades de que algunos vástagos pueden sobrevivir a todas las causas de la mortalidad. Aquellos genotipos que no tienen ese comportamiento no llegan a dejar suficiente progenie para que al menos algún vástago sobreviva, por lo que tienden a desaparecer de la población. Cuando visitamos un bosque, apreciamos un número determinado de árboles de cada especie, de las plantas herbáceas, de aves, de insectos, etc.
Ninguno de estos organismos se presenta en una sobreabundancia tal que desplace a los demás componentes de la comunidad. Las poblaciones de cada especie se encuentran en

120
estado de equilibrio, es decir, su densidad no oscila sensiblemente de un año al siguiente. Este equilibrio poblacional es alcanzado mediante la mortalidad de un elevadisimo número de individuos, en especial en formas de edad muy temprana, como semillas, plántulas, huevecillos, estados larvarios tempranos, etc. Los factores causantes de esa regulación del tamaño varían para cada especie y por lo general actúan de forma diferente.
4. - la lucha por la existencia.
Si en una comunidad se presenta un número de individuos que excede a los recursos disponibles para sobrevivir, forzosamente ocurrirá la mortalidad, que por lo general es proporcional al exceso de individuos. Recordemos que los recursos necesarios para la sobrevivencia de un individuo son diversos y pueden constituir tanto componentes físicos como biológicos del ambiente. Estos recursos son variados en extremo: agua en el suelo para las plantas, nutrientes, luz, presas para los depredadores espacio físico para llevar a cabo el cortejo, parejas para aparearse, sitios para anidar, etc. Cuando los recursos no se encuentran en suficiente disponibilidad, un individuo tiene que competir por ellos con otros individuos, ya sea de la misma o de diferentes especies. La lucha por la existencia, por lo general a través de la competición por recursos insuficientes, tiene como resultado dos elementos cruciales que determinan la mayor o menor adecuación de un individuos a un ambiente determinado: la sobrevivencia y la capacidad de dejar más descendencia. Aparte de la competición por recursos, los esfuerzos para escapar del efecto de los depredadores es otra forma de lucha por la existencia.
No es fácil apreciar la extensión de la lucha por la existencia si no se percibe que las especies tienen capacidad potencial de crecimiento exponencial. Este punto no le fue claro a Darwin hasta que leyó a Malthus a fines de septiembre de 1838, y percato de la capacidad de crecimiento numérico de las especies y la disparidad de los recursos disponibles para ellos. El hecho de que este elemento también haya inspirado a Wallace, para que paralela e independiente de Darwin concibiese la selección natural como el mecanismo causante de la evolución subraya la enorme importancia de la idea central de la doctrina malthusiana: el potencial de crecimiento exponencial de las especies.
En consecuencia...
Los cuatro elementos centrales de la teoría de la evolución se concatenan fácilmente para hacer de la selección natural el mecanismo más importante de ese proceso de cambio en las especies. Todas las especies dejan más progenie que la necesaria para reproducirse, pero ninguna alcanza a “inundar” la Tierra debido a que los recursos no son suficientes para todos los individuos y a que existen competidores y depredadores, por lo que solamente algunos individuos alcanzan la madurez y se reproducen. Los factores de mortalidad que regulan el tamaño de las poblaciones no actúan en forma indiscriminada. Debido a que todos los individuos son diferentes, su susceptibilidad a los diferentes componentes de la lucha por la existencia también lo es, por lo que algunos individuos tienen mayor probabilidad de sobrevivir y reproducirse que otros. En otras palabras, ante la variabilidad individual., El efecto de la lucha por la existencia sobre las probabilidades de sobrevivencia y reproducción de los organismos no es al azar, sino selectivo. Los individuos mejor adaptados al ambiente en el que viven serán los que tengan mayor probabilidad de sobrevivir y dejar más progenie. Si las diferencias que distinguen a los individuos que mejor adaptados son heredables entonces es posible que las características que favorecen las probabilidades de sobrevivencia y reproducción de un individuo sean heredadas a su progenie, lo cual poseerá las mismas ventajas adaptativas que sus padres. Los individuos que tienen bajas probabilidades de sobrevivencia o de reproducción, dejaran poca o nula progenie. En consecuencia, en el transcurso de númerosas generaciones, si las condiciones ambientales se mantienen constantes, la proporción de individuos bien adaptados tendera a aumentar, así como la probabilidad de que la progenie entre ellos provenga en forma creciente de la cruza entre padres cada vez mejor adaptados. El mecanismo al que se deben estos cambios en las características de una población es la selección natural y su resultado es la evolución orgánica.
La lectura continuada de las frases de cursivas del párrafo anterior representa una síntesis de los elementos fundamentales de la evolución por medio de la selección natural, tal como lo propuso Darwin.
Sin embargo...
El ambiente físico y biótico en el que viven los organismos es esencialmente variable, no solo de un lugar a otro sino también en el tiempo. Las características que hacen a un individuo

121
adaptado frente a determinadas condiciones ambientales pueden resultar inadecuadas entre condiciones diferentes. Las escalas temporal y espacial de esta variabilidad ambiental son enormes. Regiones del territorio de nuestro país, como la mayor parte de la Sierra Madre Oriental, estuvieron cubiertas por el mar en periodos tan “recientes” como hace unos 120 millones de años o menos; la porción superior de la península de Yucatán al norte de Mérida estaba sumergida hace no más de unos cuantos millones de años. Durante la ultima glaciación, hace unos 40000 años, la temperatura media anual del valle de México era de 4 a 5.5ºC más baja que la actual, de manera que se hubiera podido esquiar regularmente todos los inviernos en las montañas aledañas al Distrito Federal. La contaminación atmosférica de la ciudad de México ha modificado tan profundamente las condiciones de los bosques alrededor del Ajusco, que algunas especies de plantas se han reducido en número y otra incrementado. Las heladas de un invierno más intenso que otro matan a ciertas plantas de la misma especie en un jardín y no a otras. De igual manera, la escala de variación espacial es enorme, desde la evidente y conocida comprobación de que el clima de Tapachula es muy diferente al de Mexicali, hasta la de los distintos crecimientos de dos plantas situadas en diferentes partes de un pequeño jardín. Llevando esta idea a un extremo, no hay en la naturaleza dos “trozos” de ambientes que sean idénticos. Como consecuencia de esta variabilidad ambiental, ninguna especie presenta una distribución homogénea en este planeta; todas tienen una distribución que podemos calificar de “agregada”, es decir, de mayor concentración en ciertas arreas y escala o nulas en otras. Incluso la especie humana, quizá la más ampliamente distribuida de todas gracias a su, ingenio tecnológico, sigue ese patrón agregado de distribución, con altas concentraciones en ciertas arreas y escasa o nula población en otras, como en el Ártico, las grandes zonas desérticas, etc.
UNA NUEVA SINTESIS
Contra lo que uno hubiese pensado, el descubrimiento del trabajo de Mendel en los primeros años de este siglo despertó una polémica entre genetistas mendelianos y biólogos darwinianos sobre la forma en que ocurría la evolución y el papel de la selección natural en la variabilidad genética de las especies. Los primeros genetistas
mendelianos sostenían que la aparición de la nuevas especies obedecía exclusivamente a la ocurrencia de mutaciones cuyos efectos eran de gran envergadura, a las que llamaron macromutaciones, esta explicación se oponía, desde luego, al principio darwiniano de evolución por medio de la acumulación de cambios casi imperceptibles en las especies. Esta polémica termino cundo se extendió el campo de la genética experimental, en buena parte iniciada por los estudios de Thomas Hunt Morgan con Drosophila, y cuando algunos genetistas con excelentes bases metodológicas y matemáticas encontraron que las mutaciones que producían cambios importantes eran más bien la excepción que la regla. Algunos de ellos, como Ronald A Fhiser, John B.S. Haaldane y Sewall Wright, que estudiaban la genética de las poblaciones, llegaron a la conclusión de que las mutaciones pequeñas eran mucho más frecuentes que las macromutaciones y que tenían un efecto más importante que el que les había asignado hasta entonces. Sus estudios establecieron también que había la posibilidad muy grande de que tales mutaciones pequeñas se fijasen rápidamente en la población si resultaban favorables. También llegaron a la conclusion de que las mutaciones ligeras no eran de forma alguna la única fuente de variación genética que las especies presentaban. La recombinación de las características genéticas de los organismos, como resultado de la reproducción sexual, constituí una fuente por lo menos igualmente importante de variabilidad biológica.
Estos genetistas llegaron a demostrar que este era el proceso más importante para determinar la tasa de evolución, y que el papel de las mutaciones consistía en complementar el proceso y la variabilidad poblacional adquirida por la reproducción sexual.
Lo anterior hacia evidente que las poblaciones presentaban continuamente una gama casi interminable y constante de variabilidad biológica, sobre la cual actuaban los agentes selectivos del medio, y que el resultado de la inagotable variación natural, adicionada por los cambios más abruptos de las mutaciones puntuales.
Además la idea simplista de que cada factor genético controlaba una característica morfológica o funcional de los organismos, derivada de la observación de caracteres determinados por un solo factor. Como los que habían servido de base a los estudios de Mendel y algunos de sus seguidores a principios del siglo XX, fue rechazada por los

122
avanzados estudios cuantitativos de los genetistas. La genética cuantitativa y los estudios poblacionales de mostraron que las características morfológicas guardaban una relación extremadamente compleja con los factores genéticos. A pesar de la existencia de los factores individuales o genes, la recombinación de ellos y su acción conjunta hacen en extremo difícil establecer relaciones directas entre un determinante genético y ciertos atributos fenotípicos. Por esa razón no hay forma de aseverar que un determinado gen es el principal causante de ciertos atributos de adaptación de un organismo. El genotipo, como un todo, es el que determina lo anterior. La importancia que tiene par un individuo la posición de un gene determinado depende tanto de la forma en que este se combina con el resto del genotipo con el que le toco, por suerte, asociarse, como del efecto conjunto que esta combinación tiene sobre la adecuación de dicho individuo, es decir, sobre sus probabilidades de sobrevivencia y de reproducción. Lo mismo puede aplicarse en el caso de una mutación; su éxito depende de la forma en que se combine con el resto del genotipo que esta asociado y favorezca la adecuación de un individuo. Si la mutación en sí es favorable, pero su presencia en el genotipo produce modificaciones que van en detrimento de las ventajas de adecuación del resto de los genes, muy probablemente será seleccionada en forma desfavorable.
Aun en ausencia de mutaciones, la variabilidad natural de la recombinación genética resultante de la reproducción sexual es suficientemente grande para asegurar que, si las condiciones ambientales así lo determinan, haya un proceso de transformación en la especie de que se trata. El reconocimiento de los anterior por los genetistas modernos y los evolucionistas neodarwinianos cambio la concepción evolucionista moderna, en la que la población desempeña un papel de crucial importancia. Si bien la selección actúa en la adecuación de un individuo, la constante variabilidad genética, y por lo tanto fenotípica, de los individuos se genera solamente en el seno de la población. El interés de los biólogos evolucionistas se ha centrado consecuentemente en el nivel organizativo de la poblaciones, particularmente en la población concebida como el conjunto de individuos con los que otro puede estar genéticamente relacionado a lo largo de su vida. Otros genetistas y naturalistas hicieron contribuciones medulares al desarrollo de lo que en las décadas de los treinta y cuarenta llego a conocerse como la “síntesis moderna del Darwinismo”.
Mencionare a los tres más importantes. Theodusius Dobzhansky, un genetista de poblaciones que trabajo con la mosca de la fruta y que publico en 1937 su valioso libro La genética y el origen de las especies. Julián Huxley (nietos de Thomas), que publico lo que ha sido probablemente el mejor resumen de las ideas neodarwinismo clásico en su libro de 1942: Evolución: la nueva síntesis. Finalmente, Ernst Mayr, un eminente zoólogo y evolucionista que aun asiste a su laboratorio en el Museo de Zoología Comparada de la Universidad de Harvard y que hizo valiosas contribuciones con su obra La sistemática y el origen de las especies, publicada en 1942, cabe también mencionar a George G Simpson, que desde el campo de la paleontología, tendió un puente hacia la genética de poblaciones, demostrando que aquella se conformaba el marco estructural teórico de ésta, con lo que se completó una visión global y unificadora del pensamiento evolucionista.
Hacia la década de los sesenta el neodarwinismo parecía haber adquirido una firme posición como pensamiento unificador de la biología moderna. Pero como siempre ocurre en el desarrollo de la ciencia, nuevos hallazgos cuestionaron, al menos en parte, la coherencia de las ideas del neodarwinismo, las criticas provinieron fundamentalmente de dos lados: de los estudios de genética evolutiva y de la forma en que pueda ocurrir la macroevolucion. Las primeras surgen de observaciones que apuntan al hecho de que una buena parte de la evolución del ADN no tiene relación aparente con aspectos adaptativos del organismo y es, en consecuencia, independientemente de los procesos de la selección natural. Los productos de los genes, las proteínas estructurales, presentan niveles muy altos de diversidad y variabilidad, incluso dentro de una misma especie. Cada vez se producen más datos que sugieren que una gran parte de esta variabilidad ocurre con total independencia de los procesos de selección natural y probablemente sean el resultado de mutaciones aleatorias, combinadas con efectos de deriva genética.
No obstante lo anterior, quizá los procesos de evolución en las proteínas estructurales tengan muy poco que ver con la evolución de los organismos. Se ha encontrado, por ejemplo, que especies que difieren notablemente en aspectos morfológicos, funcionales y de comportamiento, como el hombre y el gorila, solo difieren en 14% en la secuencia de su ADN. Probablemente una parte importante de los cambios adaptativos

123
que una especie experimenta se deben a un número relativamente reducido de mutaciones cruciales que regulan el tipo de desarrollo de un organismo...
Desde el campo de la paleontología, las criticas más fuertes al neodarwinismo han venido de los proponentes de la teoría del equilibrio puntuado, Niles Eldredge y Stephen J Gould. Esta teoría mantiene que las discontinuidades del registro fósil, más que representar imperfecciones en la preservación de los organismos, reflejan el proceso mismo de la evolución, es decir, un proceso de tipo saltatorio, en que los cambios evolutivos ocurren espaciada y abruptamente en lugar de irse acumulando lentamente, como sostienen las ideas darwinianas ortodoxas. En el contexto de esta teoría, las especies permanecen sin cambios evolutivos importantes por periodos largos en los que pueden extinguirse por diversas causas, hasta que ocurre un nuevo episodio de extenso cambio evolutivo y de producción de nuevas especies. Esta teoría tiene varios problemas en su formulación, pues para tal proceso tenga lugar, es necesario suponer una serie de condiciones que es difícil que ocurran. Entre ellas están las siguientes: que la estratigrafía sea continua, que las secuencias fósiles locales no hayan estado sujetas a cambios biogeográficos rápidos e importantes y, más relevante aún, que no hayan ocurrido cambios adaptativos continuos en los tejidos suaves de los organismos. Aparte del hecho de que el material paleontológico no es lo mejor para trabajar en aspectos evolutivos y de procesos de especiación, y de que el concepto de especie para los paleontológicos ciertamente difiere del de los taxonomos, de los genetistas y de los ecólogos de las poblaciones, la teoría está aún en una etapa inicial y en el proceso de ir adquiriendo mayor información que la robustezca; debido a lo anterior, no entrare en más detalles en esta teoría. Base añadir que las ideas saltatorias de la evolución no están necesariamente reñidas con las ideas neodarwinistas y que probablemente contribuirán a enriquecerlas en el futuro próximo.
CUESTIONARIO
1. - ¿CUÁL ES LA IMPORTANCIA DE LA VARIABILIDAD EN EL MECANISMO DE LA SELECCIÓN NATURAL? ¿QUÉ CARACTERISTICAS DEBE TOMAR ESTA VARIABILIDAD PARA ESTAR EXPUESTA A SELECCIÓN NATURAL?
2. -DISCUTA LA RELACION ENTRE POLIMORFISMO Y ESPECIACION.
3. -¿QUÉ SIGNIFICA EN TERMINOS BIOLOGICOS “LUCHA POR LA EXISTENCIA”?
4.- DESCRIBA EN POCAS PALABRAS EN QUE CONSISTE EL PROCESO DE SELECCIÓN NATURAL.

124
INTRODUCCION AL ORIGEN DE LAS ESPECIES
RICHARD E. LEAKEY
INTRODUCCIÓN
En este texto encontramos una excelente reseña de la vida y los estudios de Darwin sobre la evolución. Asimismo se presentan los descubrimientos posteriores que influyeron en la postulación de la teoría sintética de la evolución y de las polémicas actuales en torno a los procesos evolutivos y la selección natural.
Darwin y su Teoría.
La primera edición completa de El origen de las especies se agoto el mismo día que apareció, el 24 de noviembre de 1859. La segunda edición salió a la venta en enero de 1860y, en total, el libro se edito seis veces en la vida de Darwin. Desde su aparición, se le han hecho númerosas reimpresiones y se han traducido a más de treinta idiomas. Aunque primordialmente, y ante todo, es un clásico científico, el origen de las especies fue escrito para el publico culto de la época de Darwin, y por eso tiene un valor literario único entre las obras de imaginación científica. Fue un libro importante en tiempos de su autor y lo que sigue siendo, ya que la teoría de la evolución es la piedra angular de la biología moderna, y la obra de Darwin es el cimiento en que descansa dicha teoría.
Sin embargo, no fue Darwin el primero en proponer que las especies de las plantas y animales pueden cambiar con el tiempo.
A fines del siglo XVIII el abuelo de Charles, Erasmus Darwin, escribió un relato sobre la evolución y, poco después, en 1809, el naturalista francés Jean Baptiste de Lamarck publico su Philosophie zoologique, que contenía sus propias especulaciones sobre la mutabilidad de las especies biológicas. De hecho, el propio Darwin menciona no menos de veinte predecesores que habían escrito sobre aspectos de la evolución. Pero es a partir de Darwin que se origina la teoría moderna de la evolución. ¿Por qué? Principalmente por dos razones.
La primera de ellas Darwin examino paciente y sistemáticamente todo tipo de evidencia relacionada con su tema. De joven paso cinco productivos años como naturalista a bordo del H.M.S. Beagle (1831-1836). Durante ese largo viaje alrededor del mundo se convirtió
en un excelente naturalista, observando, coleccionando y meditando constantemente sobre los númerosos fenómenos geológicos y biológicos con que se enfrentaba. Ya en 1837 empezó a dudar de la permanencia e inmutabilidad de las especies y, aunque entre 1837 y 1859 estuvo muy ocupado con diferentes actividades científicas, la cuestión del origen de las especies –que en aquel momento los científicos llamaban “el misterio de los misterios”- siempre lo persiguió. Durante esos años leyó mucho, penso profundamente y experimento con sumo cuidado. Como resultado, El origen de las especies es un trabajo de amplitud y profundidad extraordinarias.
La segunda razón es que Darwin fue proporcionar un mecanismo pausible para explicar como las especies pueden cambiar: la selección natural. En Darwin apareció tal idea por vez primera en 1838, después de leer el Ensayo sobre el principio de la población de Thomas Malthus, religioso y economista político de principios del siglo XIX. Inicialmente Malthus se preocupo por la población humana, pero señalo que es un principio de la naturaleza que los organismos vivientes produzcan más individuos de los que normalmente se espera sobrevivan hasta alcanzar la madurez que les permita reproducirse. Un roble da origen a cientos de bellotas anualmente; un ave procrea varias docenas de pajarillos en su vida, y un salmón desova por millares al año; cada de uno de estos descendientes puede llegar a convertirse en adulto. A pesar de esta capacidad de reproducción masiva, la población adulta tiende a permanecer en un número estable de generación en generación.
El trabajo de Malthus ayudo a Darwin a percibir un punto importante: podría haber selección entre la descendencia que sobrevive y la que perece. Puesto que los miembros individuales de una especie difieren ligeramente entre sí, aquellas que cuentan con ciertas características que les proporcionan ventajas, por ejemplo para alimentarse o para escapar de los depredadores, tendrán mayores oportunidades de sobrevivir. Aceptando la variación, cuyos resultados pueden observarse por doquier, la selección natural, de acuerdo con Darwin, puede influir en la evolución biológica (o lo que él llamó “descendencia con modificación”).
De igual manera que el hombre puede efectuar cambios sorprendentes en animales domésticos seleccionando artificialmente algunas características que considere

125
deseables, así la naturaleza “selecciona”, entre los miembros de una especie, aquellos que mejor pueden afrontar los riesgos de la vida. Como las condiciones del medio cambian, la selección natural favorece determinadas características en una población que varia al azar. Tal como afirmo el filosofo evolucionista del siglo XIX Herbert Spencer, la naturaleza garantiza “la supervivencia de los más aptos”.
Darwin por lo tanto, no fue el primero en exponer una teoría de la evolución, pero fue suyo el primer relato convincente y maduro. Resulta que aun el principio de la selección natural no fue solamente de él, ya que el naturalista Alfred Russel Wallace (1823-1915) lo detectó, independientemente, en 1858, antes de que Darwin publicara los resultados de su minuciosa investigación. Wallace no conocía a Darwin, pero supo de su reputación como naturalista que tenia opiniones nada ortodoxas, y entre ambos se mantuvo correspondencia, en términos generales, sobre el tema de la permanencia de las especies. Wallace envío a Darwin su breve ensayo On the Tendency of Varieties to Depart indefinitely from the original Type, en donde se explicaban los principios de la selección natural, sin saber que Darwin ya los había descubierto, ni hasta que punto su ensayo se adelantaba a la obra principal de Darwin. En la carta que acompañaba su ensayo, Wallace pedía a Darwin que leyera su trabajo y, de considerarlo conveniente, lo hiciera llegar a Charles Lyell. Al principio Darwin estaba desconcertado, pero Charles Lyell y Joseph Hooker, que simpatizaba con sus opiniones y lo habían urgido a que lo publicara, arreglaron todo para que una memoria conjunta Darwin-Wallace sobre la selección natural se leyera ante la Sociedad Linneana en julio de 1858, y posteriormente se publicara en el Journal of the linnean a Society ese mismo año. Tanto Darwin como Wallace se comportaron generosamente, y a pesar de que el segundo era un prestigiado naturalista, siempre otorgo a Darwin el crédito principal. Al igual que Darwin, Wallace llego a la comprensión de la existencia de la selección natural después de leer la obra de Malthus. En vista de los trabajos llevados a cabo por Wallace, Darwin desistió de su esfuerzo por escribir un extenso libro sobre la selección natural, que había empezado en 1856 (nunca lo termino), y encauzo sus energías hacia un trabajo más corto sobre la evolución.
Este trabajo fue El origen de las especies. Como hemos visto, con este libro Darwin alcanzo un doble objetivo: anuncio el
testimonio de la evolución y describió el mecanismo por el cual se pueden formar nuevas especies. Durante su vida, Darwin tuvo más éxito con la primera de dichas aportaciones, ya que el origen de las especies convenció a muchos colegas liberales de la existencia de la evolución biológica. Tan persuasiva fue la serie de evidencias recopilada por Darwin, que Thomas Huxley, al leer por vez primera su obra exclamó: “¡Que tonto fui al no pensarlo antes!” Pero aun Huxley, quien se llamó a sí mismo “guardián de Darwin” y se convirtió en el más enérgico defensor de su obra a finales del siglo XIX, no creyó que la selección natural hubiera quedado demostrada como el principal mecanismo del cambio evolutivo. Dado que la selección natural no había sido probada experimentalmente, Huxley y otros naturalistas se reservaron su entusiasta aprobación, y el propio Darwin inicio la búsqueda de mecanismos adicionales. Darwin escribió otro libro sobre uno de estos mecanismos suplementarios –la selección sexual-, el cual comento brevemente en la sexta edición de El origen de las especies. En dicha edición se basa el presente volumen. Pero es claro que aun la critica excesiva sobre lo apropiado de la selección natural como medio responsable de la evolución, no cambio la idea de Darwin de que aquella es, con mucho, el mecanismo más importante. No es hasta el siglo XX que somos capaces de apreciar cabalmente el genio de la tenacidad de Darwin, ya que la genética moderna, la observación de campo y la investigación de laboratorio nos han demostrado el poder explicativo de las teorías de Darwin. Ha quedado en claro que la síntesis del trabajo de Darwin sobre la selección Natural y la obra de Gregol Mendel sobre la genética ofrecen un cuadro coherente e inteligible sobre el cambio evolutivo, especialmente después de 1920 con los escritos de genetistas de poblaciones tales como sir Ronald Fisher, J.B.S. Haldane y Sewall Wright.
A esto se le ha llamado la síntesis neodarwiniana: darwiniana porque acepta la selección natural, y “neo” porque emplea las teorías de la herencia que Darwin desconocía, a pesar de que Mendel publico su obra clásica en 1866.
Volvamos ahora a los detalles de esta síntesis neodarwiniana. Es significativo que muchos de estos neodarwinianos de principios de siglo estuvieron interesados en poblaciones totales de plantas y animales, ya que fue Darwin, sobre todo, quien propicio el cambio de pensamiento en la biología moderna, de la

126
noción de tipología a la de población. Antes que Darwin la mayoría de los biólogos concebían las especies como grupos fijos y eternos ordenados por Dios. Puesto que Dios había creado directamente las especies individuales de plantas y animales, cada especie tenia sus características esenciales definidas –y se aceptaba tácitamente que eran las mismas que poseía el individuo original la pareja reproductora de cada tipo de especie. Aunque algunos naturalistas reconocían que era posible cierta variación dentro de la especie, los científicos predarwinians insistían en que había limites naturales para tal variación.
Darwin demostró como la “descendencia con modificación” puede dar origen a la aparición de nuevas especies. Al igual que los biólogos modernos, constató que, en un punto fijo en el tiempo, las especies tienen una existencia real: la noción biológica no es tan solo una abstracción humana.
Para los fines de su argumento, Darwin evito dar una definición exacta pero, en El origen de las especies, esta implica una ideas de especies semejantes a la concebida por científicos contemporáneos, quienes la definen como aquella población que simultáneamente es comunidad reproductora, unidad ecológica y unidad genética. Esto significa que los individuos que constituyen determinada especie pueden cruzarse entre sí, ocupan el mismo nicho ecológico, generalmente compiten por el mismo alimento y sufren las mismas clases de depredadores; también comparten el mismo conjunto de genes que pasa de generación en generación. Darwin había aceptado la mayoría de los métodos que emplea el biólogo moderno para diferenciar las especies y explicar el proceso de formación de las mismas.
No debe pensarse que la biología evolutiva no haya alcanzado grandes logros desde los tiempos de Darwin. Los ha tenido, y el propósito de la presente edición es poner al alcance del lector moderno la obra de Darwin, así como explicar de que manera la investigación moderna ha ampliado nuestro conocimiento acerca de los problemas que él enfrento... El origen de la especies es en la actualidad una introducción tan buena, como cualquier otra, para el tema de la evolución. El genio de Darwin trasciende el siglo que nos separa de él y lo convierte, al mismo tiempo, en un científico histórico de gran originalidad y en contemporáneo nuestro.
Los problemas de Darwin
Una de las virtudes de El origen de las especies es la forma en que Darwin manejó sin temor las difíciles confrontaciones de su teoría sobre la descendencia con modificación por selección natural. Como la biología evolucionista no proporciona por si misma la clase de pruebas en que se apoyan otras disciplinas, tales como la química y la fisiología, es todavía, hasta cierto punto, una ciencia deductiva, y al leer El origen de las especies nos sorprendemos de la frecuencia con la que Darwin trabajo integrando conjunto de hechos y observaciones, explicables a través de su teoría, pero difíciles de comprender por los supuestos convencionales de su época. La teoría de Darwin es bella y grandiosa porque explica mucho con poco; y no sorprende que el status de las evolución por selección natural sea comparado a menudo con la teoría de las gravitación universal de Newton, que atañe tanto a la caída de una manzana como a los movimientos de las mareas y a la órbita de un planeta alrededor del sol.
No obstante, Darwin tuvo problemas, muchos de los cuales reconoció y abordo en la primera edición de El origen de las especies. Después de 1859, los críticos suscitaron otras interrogantes que fueron consideradas por Darwin en las ediciones subsecuentes. En la mayoría de los casos, Darwin logro proporcionar explicaciones completamente adecuadas a las diversas anomalías y dificultades... convendría mencionar aquí tres o cuatro de los mayores obstáculos que tuvo que enfrentar Darwin. En esta sección nos dedicaremos sobre todo al Darwin histórico, pero mencionare brevemente algunos avances recientes, reservando para la sección final una discusión más extensa sobre los aspectos modernos de la biología evolutiva.
Podemos empezar con un problema que, en principio, no interesó particularmente a Darwin: el tiempo en las primeras décadas del siglo XIX la literalidad de la cronología bíblica sobre la historia de la Tierra había sido seriamente objetada. Debido a los trabajos de geólogos y paleontólogos tan prestigiados como Georges Cuvier en Francia y William Smith y Charles Lyell en la Gran Bretaña, tanto los científicos como el hombre culto percataron de la inmensidad del tiempo geológico. Supieron que la tierra estuvo inicialmente habitada por especies de plantas y animales que ya no existen: en particular, la imaginación victoriana, al igual que la nuestra, simpatizó hasta el romanticismo con los grandes dinosaurios y otras bestias

127
desaparecidas, cuyos restos podían encontrarse en la Gran Bretaña, para celebrar el año nuevo de 1853, Richard Owen (quien más tarde se convertiría en uno de los más tenaces críticos de Darwin) cenó esplendorosamente con un grupo de personas dentro de la reproducción de un dinosaurio. Ya en 1831, cuando se embarco en el Beagle, Darwin sabia que los fenómenos geológicos que atestiguaría seguramente no iban a encajar en la escala del tiempo de 6000 años calculada según una estricta interpretación de la Biblia. La obra de Charles Lyell, principles of Geology, tuvo especial importancia para Darwin. El primer volumen se publico a finales de 1830, antes de que abandonara Inglaterra, y el segundo lo alcanzo en América del Sur. Lyell se dio a la tarea de explicar la teoría de la Tierra refiriéndose tan solo a causas vigentes. En particular estas causas comprendían la elevación de la Tierra por acciones volcánicas o sísmicas y la erosión por efectos del agua y el viento. Ya que el fenómeno para abordar consideraba no solamente las grandes cadenas montañosas como los Alpes y loa Andes, sino también la existencia de fósiles marinos en estratos que aparecen en la parte superior de las montañas, era evidente, pues, que el programa de Lyell requería de lapsos muy amplios para tener éxito.
La elegante explicación de la obra de Lyell impresiono a Darwin, ya que parecía garantizar que el tiempo no representaba problema alguno. Sin embargo, el testimonio de los fósiles era otra cosa. Uno de los descubrimientos que se apoyaba la revolución del tiempo de principios de siglo XIX era la aceptación de que ciertas clases de plantas y animales fosilizados estaban asociados con determinados estratos geológicos. Por lo tanto, los fósiles podrían servir como ayuda en la ubicación relativa de los estratos en el tiempo. A partir de 1820 y con el progreso de las ciencias gemelas, la geología y la paleontología, las observaciones estratigráficas de diversas partes del mundo empezaron a ser integradas para proporcionar una imagen coherente de la historia de la vida sobre la tierra.
A pesar de que la secuencia de los fósiles parecía tener una orientación, pues se encontraban plantas y animales simples antes que las formas de vida más complicadas, y los fósiles de estratos más recientes representaban mayor semejanza con los seres vivientes, los geólogos anteriores a Darwin no ofrecieron una interpretación evolucionista a sus descubrimientos. Había tres principales
razones científicas para esto. En primer lugar, los fósiles más antiguos que se conocían eran de animales hasta cierto punto complicados – la mayoría de los invertebrados marinos, tales como moluscos y crustáceos. Los geólogos de 1830 estaban convencidos de que habían encontrado el origen de la vida en el estrato llamado cámbrico, y ninguna hipótesis, salvo la de la creación especial, se juzgaba adecuada para explicar la súbita aparición de los fósiles cámbricos. Parecía que las formaciones anteriores al periodo cámbrico estaba totalmente desprovistas de fósiles. En segundo lugar, los diferentes estratos tenían generalmente sus fósiles propios y característicos de flora y fauna, y la transición entre los diferentes estratos de abrupta. Esto represento para los geólogos la probabilidad de sucesiva creaciones y extinciones masivas. Por lo tanto, la manera más sencilla de conciliar el Génesis con la geología era pensar que la historia bíblica se refería solamente a la ultima creación, la de las plantas y animales actuales, incluyendo por supuesto al hombre mismo. Se argumentaba que no había razón para que la historia de los periodos anteriores se incluyera en el génesis, puesto que no tenían relación con las necesidades espirituales del hombre. Aun en 1820 se presentaron serios problemas ante esta concesión, pues el descubrimiento de vestigios humanos en cavernas inglesas podía ser interpretado (y lo fue posteriormente) como prueba de la coexistencia del hombre con otras especies de mamíferos desaparecidos, tales como los mamuts. Sin embargo, hasta el final de la década de 1850, universalmente se suponía que el hombre era de origen reciente. Cuando la evidencia que atribuía una gran antigüedad al hombre se hizo irrefutable
Al inicio de la década de 1860, se beneficio el caso de Darwin, aunque este nunca lo explotó en El origen de las especies, pensando que un tratado demasiado extenso sobre el hombre predisponía a los científicos en contra de los aspectos más generales de su teoría de la descendencia con modificación. Darwin mismo sostuvo que el hombre es resultado de la evolución, como lo atestiguan sus notas de finales de la década de 1830. Considero el caso de la evolución humana en dos libros publicados después de 1859: The Deseen of Man and Selection in Relation to Sex de 1871, y The Expression of the Emotions in Man and Animals de 1872.
Las consideraciones religiosas, tuvieron sin lugar a duda, un papel muy importante en la negativa de los geólogos a dar una interpretación evolucionista a sus

128
descubrimientos paleontológicos. Pero además de la repentina aparición geológica de signos de vida en el periodo cámbrico y las abruptas transicione4s entre periodos contiguos, había otra razón íntimamente ligada con esto, para que la creación individual, más que la evolución gradual, fuera la consideración científica predominante antes de la aparición de El origen de las especies. Tal razón significaba la dramática aparición, a través del testimonio de los fósiles, de algunos grupos superiores, en particular de diversas clases de vertebrados (peces, anfibios, reptiles, aves, mamíferos). Parecía que había pocos fósiles en transición, y el razonamiento fisiológico concebía que no existía una transición gradual entre las branquias y los pulmones, o entre un miembro delantero de un vertebrado y un ala. Algunos evolucionistas anteriores a Darwin, tales como Geoffroy St. Hilaire de Francia y Robert Chambers de la Gran Bretaña, desarrollaron hipótesis basadas en la suposición que podían ocurrir cambios bruscos durante el desarrollo embriológico, pero la mayor parte de los científicos pensaba que con esto solo se sustituía un milagro por otro y continuaban creyendo en una creación especial. Este problema era especialmente agudo para Darwin, ya que el siempre insistió en que la evolución era gradual y no esporádica, y también se enfrento a los testimonios de fósiles que parecían indicar los cambios bruscos.
El origen de los grupos superiores es aun tema de discusión entre los paleontólogos, aunque los nuevos descubrimientos fósiles después de Darwin han limitado muchas diferencias entre los recuentos que ofrecen los fósiles. Ya en la época de Darwin se encontraron ciertas formas en estado de transición, como el Archaeopteryx, que es un producto intermedio entre los reptiles y las aves. Otros descubrimientos han disminuido las brechas entre los peces y los vertebrados de Tierra y entre los reptiles y los mamíferos. Muchos paleontólogos piensan actualmente que la evolución siempre ha procedido en la misma escala gradual, y que la ausencia de formas transitorias puede explicarse con los argumentos que empleo Darwin, especialmente el de que hay inmensas brechas en el testimonio de los fósiles y que la transición generalmente se presenta en una localidad restringida, y la migración de la nueva forma ocurre cuando esta ha alcanzado cierto grado de avance. Pero algunos paleontólogos, sobre todo Stephen Jay Goul, de harvard, creen, como resultado de la evidencia de los fósiles, que en diferentes etapas de la historia de la vida de la evolución
ha progresado con desusada rapidez –como en brotes -, y que las ramas más tupidas del árbol de la evolución se han dado en esas ocasiones. Pudo haber ocurrido porque un grupo en evolución tal vez alcanzó una etapa en su organización en la que tenía una ventaja sobre otros grupos y, una vez que traspuso ese umbral, fue capaz de difundirse rápidamente y ocupar múltiples nichos. Un “brote” puede resultar de un cambio considerable en el clima, o de la extinción de otro grupo de especies que dejaron muchos nichos vacantes.
En relación con la evolución de los grupos superiores aparece el problema más generalizado de los órganos complejos. Es difícil concebir su desarrollo gradual, ya que solamente el órgano funcional completo es útil para el individuo. ¿De que manera una línea evolutiva puede dar lugar a plumas para volar, si las plumas rudimentarias no tienen uso? El propio Darwin estaba especialmente preocupado por los problemas relacionados con el ojo, ya que con frecuencia los creacionistas citaban ese órgano como una prueba de que Dios había creado cada especie individualmente. En 1860 Darwin confeso al naturalista norteamericano Asa Gray: “aún ahora, el ojo me da escalofrío: pero cuando pienso en las sutiles gradaciones conocidas hasta ahora, mi razón me dice que debo superar el escalofrío”. El enfoque de Darwin ante el problema general fue semejante a los biólogos modernos: una combinación de anatomía comparada, embriología, fisiología, paleontología y razón.
A pesar de estas cuestiones, Darwin estaba en lo cierto al decir que el testimonio de los fósiles no presenta problemas insalvables para la teoría de la evolución... como veremos en la siguiente sección, en el curso de las dos décadas pasadas la micropaleontología ha ampliado enormemente la historia de la vida sobre la Tierra, de una manera que hubiera satisfecho a Darwin. Es decir, que la paleontología moderna, con sus grandes descubrimientos y nuevas técnicas, como la fechar por medio de la radiactividad, simplemente ha confirmado sus teorías. Pero en relación con otro de sus problemas –el de la herencia -, la ciencia moderna ha podido avanzar más allá de Darwin. Darwin no conoció el mecanismo de la herencia, pero sabía que las variaciones ocurren y que al menos algunas de ellas se heredan. Estaba particularmente interesado en las variaciones relativamente amplias que pueden observarse en las plantas y animales domésticos, no solamente por la analogía que encontró entre

129
la selección natural y artificial, sino porque también las plantas y animales domésticos parecían ofrecerle una oportunidad mejor para comprender el fenómeno general de la herencia.
Darwin mismo llevó a cabo experimentos para criar algunas especies domesticas, especialmente con palomas. (La paloma aparece frecuentemente en El origen de las especies. El editor de Darwin envío el manuscrito para que diera su opinión. Él critico manifestó que sentía mucho que Darwin no hubiera escrito simplemente un libro sobre la cría de las palomas, y agregaba: “Todo mundo esta interesado en las palomas. El libro seria comentado en todas las revistas del reino y pronto encontraría en los mostradores de toda librería”). Con base en sus experimentos con especies domesticas, Darwin escribió un tratado sobre domesticación, que se publico en dos volúmenes en el año de 1868, el que desarrollo una teoría sobre la herencia, a la que domino “pangénesis”.
En ésta teoría Darwin propuso que cada órgano del cuerpo emite unas partículas microscópicas a las que llamo “gémulas”, y que estas gémulas se reúnen en los órganos reproductores en donde pueden influir en la forma, tamaño y funciones de los órganos correspondientes de la descendencia. Darwin deseaba explicar su creencia diciendo que algunas características o hábitos adquiridos durante la vida de los padres podrían afectar los atributos correspondientes de sus crías. A este fenómeno se le dio el nombre de “herencia por uso y desuso”; pero también se le conoce de un modo más familiar como “la herencia de caracteres adquiridos” o “lamarckismo”, en honor de Jean Bapstiste de Lamarck. Sin embargo la teoría de Lamarck era algo diferente, ya que presuponía además un deseo de cambio, que causaba que una modificación se presentara en el propio organismo y después se transmitiera a su descendencia. En la época de Darwin, en general no se acepto tal aspecto de la teoría de Lamarck, aunque casi todos los científicos creían que los caracteres adquiridos por uso y desuso podrían ser heredados.
Darwin desarrollo su teoría de la pangénesis para poder describir como se presenta la herencia por uso y desuso. Sin embargo se percato de que su teoría era meramente especulativa y no le puso mucho entusiasmo. Cuando fue duramente criticado por su primo Francis Galton (eminente psicólogo, estadígrafo y estudioso de la herencia), Darwin abandono calladamente la pangénesis,
aunque continuo aceptando la herencia de los caracteres adquiridos. Sin embargo, Darwin insistía correctamente en que la selección natural solo requería que la variación hereditaria ocurriera. La selección natural no implicaba ningún mecanismo particular de la herencia y ésta es la razón por las que las teorías de Darwin sobre la evolución se han integrado tan fácilmente a la genética moderna. Como Darwin reconoció que no comprendía los mecanismos hereditarios, trato de concentrarse en lo que hoy conocemos como el nivel de variación fenotípica... muchas de las observaciones que hizo sobre la herencia, variación e hibridismo todavía pueden ser aceptadas, aunque en otros aspectos podemos ver que Darwin confundía los procesos genéticos con los fisiológicos. El biólogo alemán August Weismann (1834-1914) fue quien, a partir de 1870, hizo todo lo posible por convencer al mundo científico de que la sustancia hereditaria, o “plasma germinal” como la llamo, pasaba de la generación en generación sin ser influida por cambios corporales adquiridos a través del ejercicio o la falta de él, o por lesiones o enfermedades, la idea de Weismann sobre la “continuidad del plasma germinal” estaba basada en observaciones de las variaciones fenotípicas, semejantes a las que efectúo Darwin, y en el progreso del conocimiento de la estructura y funcionamiento de las células y de sus componentes, como el núcleo. La mayoría de los científicos han aceptado que los descubrimientos de Weismann y de otros descartaron el lamarckismo, pero aun en el siglo XX ha habido voces disidentes, las más famosas han sido las del genetista austríaco Paul Kammerer (1888-1926) y la del biólogo ruso Trofim Denisovish Lysenko (1898-1976).
El trabajo de Kammerer ha sido descrito por el iconoclasta Arthur Koestler en The Case of the Midwife Toad. Kammerer estudio los caracteres adquiridos de muchos animales, pero su experimento más famoso fue con el llamado sapo partero, Alytes Obstetricans. La mayoría de los sapos regresan al agua para cruzarse y a los machos se les desarrolla una callosidad en las extremidades anteriores que les permite sujetar a la hembra durante el apareamiento en el agua. Pero el sapo partero se reproduce en tierra y los machos no tienen esa callosidad. Kammerer sostenía que cuando los sapos parteros se vieron forzados a aparearse en el agua, los machos desarrollaron las "“callosidades nupciales” pigmentadas que heredaron sus crías. La mayoría de los científicos de su tiempo desecharon los resultados de los experimentos

130
de Kammerer, sobre todo cuando se descubrió que las callosidades prenupciales pigmentadas de la especie que este presento no eran sino tinta china cuidadosamente inyectada bajo la piel. Poco después de esta revelación Kammerer se suicido. Koestler mantiene que el suicidio de Kammerer fue resultado directo del desprecio de la comunidad científica hacia él, pero la revuelta social que estallo en Austria después de la primera guerra mundial también pudo haber contribuido a esta tragedia. Es probable que Kammerer no haya efectuado el fraude; más aun, es posible que haya sido víctima de un complot. Esto nunca lo sabremos. De cualquier forma, Koestler insiste en que la mayor parte del trabajo experimental de Kammerer todavía tiene actualidad y que solo el dogmatismo científico rechaza la interpretación lamarckiana del fenómeno descrito por Kammerer.
El caso de Lysenko es todavía más patético, ya que trato de ganarse la simpatía de Stalin, y así influyo en la política agrícola soviética durante dos décadas posteriores a 1930. Lysenko no era genetista y pronto los científicos occidentales desacreditaron sus trabajos lamarckianos por extravagantes. Pero en una Rusia crónicamente deprimida por falta de alimentos, la promesa de Lysenko de obtener mayor productividad en la crianza de animales de granja y en el cultivo de cereales, como trigo y centeno, era seductora. La herencia de caracteres adquiridos tiene la ventaja que puede permitir que la evolución proceda mucho más rápido que el azaroso proceso de la variación por selección natural. Las críticas mendelinas a Lysenko fueron acalladas en Rusia y aquellos que se rehusaron a aceptar sus ideas se vieron destruidos de sus puestos y enviados a prisión. Irónicamente entre aquellos que murieron en prisión se encontraba el único hombre que podía haber logrado el incremento de la productividad agrícola deseada por Stalin. Ese hombre era Nikolai Ivanovich Vavilov, quien en su trabajo sobre los orígenes de las plantas expuso las bases para gran parte del cultivo moderno. Al final, las predicciones de Lysenko no dieron resultados y, con la muerte del régimen estilista, Lysenko cayó en desgracia.
Esto y otros acontecimientos del siglo XX confirman nuestra perenne fascinación por el lamarckismo parece ser que en la psique humana existe algo profundamente arraigado que quiere que los esfuerzos y logros de nuestra generación se perpetúen en los genes de las generaciones futuras. Como veremos, el llamado dogma central de la genética
molecular afirma que los acontecimientos somáticos corporales, n pueden traducirse en información genética individual. Darwin estaba en un error al aceptar la herencia por uso y desuso, pero acertaba al reconocer que la teoría de la selección natural no dependía de esta cuestión.
Darwin también estaba equivocado en su exposición de lo que se llama “herencia combinada”, y varios críticos señalaron que la herencia combinada presentaba problemas al concepto de cambio evolutivo. Darwin sostuvo que generalmente cada progenitor contribuye de igual forma a la composición física de su descendencia; así que cualquier atributo heredado, como la altura o la coloración, en general, podría ser igual al promedio de los padres. Pero, como señalo en 1867 uno de los críticos de Darwin, Fleeming Jenkin, esta opinión tiene serias consecuencias para la perpetuación de la innovación evolutiva. Si ocurre la herencia combinada, cualquier innovación hereditaria que se presente en un individuo se introducirá en las generaciones siguientes; la primera generación tendrá la innovación a la mitad, la siguiente a una cuarta parte y así sucesivamente, hasta que el efecto de la innovación ya no se exprese. D Darwin creía que la herencia por uso y desuso podía resolver parcialmente este enigma, ya que aumentaba la proporciona de la variación heredada. Salvo por esto, Darwin únicamente insistía en que la variación ocurre –“debido a causas que ignoramos”- y para que aparezca una nueva variedad o especie, la misma variación deberá ocurrir repetidamente y la población deberá ser aislada para evitar que la variación fracase.
Sin embargo, a Darwin le atraía la herencia combinada, ya que concordaba con su hipótesis fundamental de Natura non fact saltum (la naturaleza no da saltos). Esta hipótesis también fue la base para su creencia de que el cambio evolutivo siempre ocurre gradual y lentamente. Sus teorías fueron claramente beneficiadas por la revolución en el concepto del tiempo geológico mencionada al inicio de esta sección. Así, el intento de Darwin por incursionar en un análisis cuantitativo del tiempo geológico le produjo nuevas inquietudes. En la primera edición de El origen de las especies se incluía un calculo basado en porcentajes de erosión estimados, en donde se sugería que el Weald, ancha franja boscosa que se localiza entre North y South Downs, en Inglaterra, y que representa los restos erosionados de una cadena de colinas, debió de necesitar cerca de 300 millones de años para desgastarse hasta su

131
nivel actual. Si fuera cierto, pensó la Tierra tendría que ser mucho más antigua. “Que incomprensiblemente vastos han sido los periodos pasados"” escribió en la primera edición de El origen de las especies. Sin embargo, cuando apareció la primera edición de su obra, algunos físicos como William Thomson (más tarde lord Kelvin), estaban aplicando la termodinámica para estudiar la edad de la Tierra. Su cálculo basado en porcentajes de pérdida de calor de la superficie terrestre, era de casi 200 millones de años; es decir, menos de lo que Darwin había asignado para la formación de Weald. Mientras Darwin discretamente suprimía este calculo de las ediciones posteriores de El origen de las especies, el concepto del tiempo lo seguí preocupando. Varios críticos que no aceptaban la evolución utilizaron el trabajo de Thomson, el físico de más prestigio en la Gran Bretaña, como un arma contra Darwin, quien a su vez tuvo que admitir que los 200 millones de años que, según Thomson, habían transcurrido desde la consolidación de la corteza terrestre, apenas eran suficientes para el desarrollo de las distintas formas de vida.
A pesar de este problema, Darwin se sostuvo y no abandono su teoría ni cayo en la especulación de que cambios bruscos e irregulares pudieron milagrosamente haber acelerado la evolución. El problema no se resolvió hasta después de la muerte de Darwin, cuando el descubrimiento de la radiactividad señalo otra fuente de energía dentro del núcleo de la Tierra, y así se demostró que las cifras de Thomson eran demasiado conservadoras.
Las partículas radioactivas en descomposición proporcionan ahora a los geólogos un método para fechar las rocas y determinar con bastante precisión la edad de los estratos y los fósiles, cosa que Darwin no pudo hacer. La culminación al dilema de Darwin ante el tiempo se debió a que un de sus hijos, el distinguido astrónomo sir George Darwin, fue de los primeros en sugerir que la radioactividad debe tomarse en cuanta al revisar los cálculos sobre la edad de la Tierra. Las estimaciones actuales señalan que la edad de la Tierra es de unos 4600 millones de años. Desafortunadamente, las mediciones del Weald que Darwin realizó eran erróneas; Ahora se sabe que su antigüedad varia entre 20 y 30 millones de años.
Estos problemas – del testimonio de los registros fósiles; del origen de adaptaciones complejas y grupos biológicos superiores; de la herencia y la variación; y del tiempo necesario para que ocurra el proceso
evolutivo- son solamente algunos a los que tuvo que enfrentarse Darwin. Muchos otros problemas pueden entresacarse de El origen de las especies. En ediciones posteriores de su gran obra, Darwin se esforzó por incorporar nuevos conocimientos y responder a los críticos. Aunque el origen de las especies cambio considerablemente entre la primera y la sexta edición, el objetivo de Darwin se mantuvo constante: demostrar el poder de la selección natural en la formación gradual de nuevas especies. La ciencia moderna ha demostrado con creces la validez de la confianza que Darwin tuvo en sus propios descubrimientos.
DARWIN Y NOSOTROS
Darwin presento diversas clases de información relacionada con el tema de la evolución; la herencia y la variación, los fósiles, las formaciones geológicas, la distribución geográfica, la embriología, la taxonomía y la homologa. Hay trabajos recientes sobre la evolución que muestran la vigencia de su investigación. De la época de Darwin a la fecha, ha habido avances individuales en cada una de tales disciplinas, en ocasiones, han cambiado tanto que son irreconocibles si se les compara con lo que fueron el siglo pasado. Pero Darwin fue el primero que mostró donde buscar las pruebas de la evolución. En esta sección examinare algunas de las formas en que la investigación moderna ha conjuntado las partes de ciertos rompecabezas preparados por Darwin, y como Ha dado respuesta a preguntas que él mismo no pudo ni siquiera formular.
El primer campo en que nuestra comprensión de la evolución trasciende a la de Darwin es el de la genética. Todos entendemos los principios aunque no los detalles de la herencia, y sin duda la genética molecular es el logro individual más grande de las ciencias naturales de nuestro siglo. La genética moderna se deriva del trabajo del monje austríaco Gregor Mendel (1822-1884) y Darwin podría haberse beneficiado de él, pero la importancia de tales observaciones no fueron apreciadas hasta después de la muerte de ambos. Es un malentendido común decir que los trabajos de Mendel sobre el hibridismo en las plantas, de 1866, fueron desconocidos hasta 1900 de hecho, hay por lo menos ocho referencias al trabajo de Mendel en la literatura científica de 1866 a 1900. Sin embargo, su trabajo no causo impacto alguno entre la comunidad científica, porque la teoría de Mendel parecía estar en desacuerdo con las

132
observaciones generales sobre la herencia. Aquellos que leyeron el documento de Mendel pensaron que sus observaciones sobre la herencia de partes eran excepciones a la regla general. Fue hasta este siglo cuando se reconoció la importancia de la teoría de Mendel.
Los descubrimientos de Mendel sobre los patrones de la herencia en las plantas de jardín, como los chícharos, lo llevaron a proponer la idea de la herencia de partes. Al estudiar algunos rasgos como el color de la flor, la altura de la planta y la forma y textura de la semilla, Mendel notó que las contribuciones paternas se expresan con desigualdad: la combinación no tiene lugar.
Para explicar los resultados, Mendel propuso que estos rasgos o características eran heredados como “elementos” (a los que ahora llamamos genes). Se recibe un elemento de cada progenitor. Algunos elementos son dominantes y otros son recesivos; si la cría tiene dos elementos o un elemento dominante y uno recesivo, se manifestara el rasgo o característica dominante; si solamente cuenta con dos elementos recesivos, el rasgo recesivo será el que aparezca.
Mendel eligió deliberadamente caracteres simples, representativos de extremos opuestos, para sus experimentos; esto es, caracteres con formas claramente perceptibles y no intermedias. A los caracteres que se heredan en esta forma se les llama mendelianos. Sin embargo, la mayoría de los caracteres están controlados por un número de genes que actúa conjuntamente y modifica las actividades del otro grupo. Por lo tanto, las proporciones o relaciones muy simples difícilmente se observaran al estudiar la herencia, a pesar de que cada gen por separado es heredado a la manera mendeliana. Por este motivo, se pensó inicialmente que la herencia de partes era la excepción y no la regla.
El propio Mendel desarrolló los conceptos de dominante y recesivo, y su trabajo abarca una clara distinción entre genotipo y fenotipo. Lo que nosotros observamos directamente es el fenotipo (la forma de la semilla o el color de los ojos). Mientras que al conocimiento del genotipo, que produce lo que observamos, solamente puede llegarse por medios más sutiles. Para el año 1900 cuando ya se apreciaba en todo su valor el trabajo de Mendel, los microscopistas tenían una buena idea de lo que sucedía en dos clases de divisiones celulares, la mitosis y la meiosis. En la mitosis se puede observar que algunos
filamentos de núcleo de la célula, llamados cromosomas, se dividen y cada célula hija recibe una dotación completa de los mismos. La mitosis es la forma común de división celular. Pero las células sexuales atraviesan por un proceso de división más complicado, llamado meiosis. La meiosis conduce a la formación de gametos, esto es, óvulos y espermatozoides. Incluye dos divisiones del núcleo, pero solo una duplicación de cromosomas. Del proceso anterior resultan cuatro núcleos hijos, cada uno de los cuales tiene solamente la mitad de los cromosomas de la célula madre. Durante la reproducción, cada progenitor contribuye con un gameto, de manera que cuando el óvulo y el espermatozoide se unen, el producto es un óvulo fecundado con una dotación completa de cromosomas.
La observación del comportamiento de los cromosomas durante la meiosis llevo a pensar que pueden seguir los patrones de herencia estudiados por Mendel. En 1909 el biólogo danés Wilhelm Johannsen llamo gen a la unidad mendeliana de la herencia, y los trabajos experimentales efectuados por el genetista norteamericano Thomas Hunt morgan y otros, determinaron, por la misma época, que los genes se encuentran en los cromosomas. Morgan llevó a cabo la mayor parte de sus experimentos con la mosca de la fruta, o del mediterráneo, Drosophila melanogaster, y todavía es este insecto el que continua aportando gran parte de la nueva información genética.
Cada especie de planta o animal tiene un número característico de cromosomas en sus células; la Drosophila melanogaster tiene ocho, los seres humanos tienen cuarenta y seis. Sin embargo, seria más correcto decir que las células humanas tienen veintitrés pares, ya que cada cromosoma tiene su pareja, en el lado opuesto, durante la división celular. Por lo tanto, el núcleo de cada célula somática contiene dos series complementarias de genes, mientras que el gameto posee solamente una serie, esto es, un miembro por cada par de cromosomas. La importancia de esto, en relación con la teoría de Mendel, es obvia. Cada miembro de un par de cromosomas homólogos lleva un filamento de genes cifrado con los mismos caracteres. Así, como cada cromosoma tiene su pareja, cada gen también tiene lo que podríamos llamar su “gen hermano”. Estos dos genes no son necesariamente idénticos: controlan el mismo carácter, pero pueden producir efectos completamente diferentes en él. De cada par de genes, uno proviene de cada uno de los

133
progenitores. Una cría puede heredar un gen idéntico de cada padre; entonces se dice que es homocigótico para ese gen. Si cada progenitor contribuye con un gen correspondiente diferente, la cría es heterocigótica. Estos términos solamente se refieren a los genotipos, pues la dominancia encubre la expresión fenotípica de la heterocigocis, y ésta puede manifestarse en la aparición de rasgos recesivos de la cría.
Estoas conceptos son importantes para comprender el sexo, ya que el sexo de un individuo es determinado genéticamente en la fecundación.
Hombres y mujeres tienen veintidós pares de cromosomas en común, pero los cromosomas del par vigésimo tercero –llamados cromosomas sexuales- son diferentes para cada sexo. Las mujeres tiene dos cromosomas X, mientras que los hombre tienen un cromosoma X y uno Y. Todas las células de los óvulos humanos (con veintitrés cromosomas) contienen un cromosoma X, mientras que las células de los espermatozoides contienen un cromosoma X o uno Y. El sexo del producto es determinado, entonces, por la clase de espermatozoides (X o Y) que fertiliza al óvulo.
Los cromosomas Y son mucho más pequeños que los cromosomas X, y tienen menos genes. Como la mujer tiene dos cromosomas X, rara vez sufre de trastornos genéticos recesivos que portan esos cromosomas X. Ella puede portar un gen recesivo nocivo en uno de sus cromosomas X sin que sufra consecuencias personales. Sin embargo, la mitad de sus hijos varones (en promedio) sufrirán la expresión del gen, ya que el cromosoma Y del hijo no tendrá un gen complementario al gen recesivo dañino del cromosoma X. Los rasgos o características heredados en esta forma, se conocen como vinculados al sexo. La hemofilia es la enfermedad más conocida que esta vinculada al sexo. En tiempos de Darwin ya se conocía, pero su patrón de herencia no pudo explicarse por la teoría de la herencia combinada. Las variaciones genéticas son debidas a la reorganización accidental de los cromosomas, que ocurre junto con el intercambio de partículas de pares de cromosomas correspondientes durante la meiosis. A este proceso se le llama intercruzamiento. El resultado final de tales procesos es que la cría recibe una combinación de los genes que poseían sus progenitores. Sin embargo, ocasionalmente puede ocurrir una mutación y un gen completamente nuevo se llega a producir. En ultima instancia, las mutaciones son el incentivo para la evolución, pues es a través
de ellas que aparecen genes completamente nuevos y, por lo tanto, nuevas variaciones pueden ocurrir. Las mutaciones genéticas son acontecimientos fortuitos, aunque puedan incrementarse con la exposición a la radiación y a ciertos productos químicos.
La mayoría de las mutaciones son nocivas ya que disminuyen las aptitudes entre las crías y, por lo tanto, son eliminados mediante la selección natural. Sin embargo, en ocasiones se produce una mutación benefica y los individuos que la llevan adquieren mayores aptitudes (mas adelante veremos algunos ejemplos). Las nuevas especies se forman por acumulación gradual de genotipos transformados, debido a una combinación de la variación genética y de la selección natural: a trabes de “el azar y la necesidad”, como lo expuso el microbiólogo francés Jacques Monod. Este es un planteamiento contemporáneo de la parte medular de la teoría de Darwin. Podemos ser aun más precisos, ya que durante los últimos veinticinco años la genética mendeliana se ha transformado en genética molecular. Ahora podemos comprender la base molecular de algunos de los procesos biológicos descritos anteriormente.
En 1953 James Watson y fransis Crick anunciaron que habían dilucidado la estructura química del material genético: el ácido desoxirribonucleico (ADN). El ADN es una molécula larga que consiste en una doble cadena de unidades alternas de azúcar y fosfato. Adherida a cada unidad de azúcar está una de estas cuatro bases: adenina, timina guanina o citosina (A,T,G,C). La molécula larga de doble filamento está trenzada en forma de hélice y, esta doble estructura helicoidal es de tal forma que puede “zafarse”, y cada mitad del filamento de ADN puede sintetizar al medio filamento complementario. El proceso se presenta cuando los cromosomas se duplican durante la división de la célula. La secuencia de las bases es la que traduce claramente el mensaje genético de la molécula de ADN. Las “letras” individuales que forman un mensaje están compuestas por tres bases consecutivas. Así, las secuencias de los elementos ATTCGACGC se leería: ATT,CGA,CGC.
Se ha determinado que tres secuencias particulares en forma de tercias, significan “alto”. Un gen principia y termina con la señal de alto. Todas las demás secuencias de tercias se codifican para los aminoácidos o sustancias químicas que forman la estructura de las proteínas. Hay veinte o más tipos diferentes de aminoácidos, y la proteína se forma de una

134
cadena de ellos. Los aminoácidos presentes y su secuencia dentro de la cadena, es lo que otorga ala proteína sus propiedades bioquímicas. Así, el código de la genética se traduce en una proteína con funciones específicas en la célula de un organismo vivo. Lo sorprendente es que el código genético parece ser universal, ya que, hasta donde se sabe, el código de tercias de bases es el mismo para los mismos aminoácidos en todas las formas de vida sobre la Tierra. Este hecho es, en sí, una rotunda evidencia de la unidad de la vida de los senderos comunes de la evolución.
La otra asombrosa implicación de este diseño es que la función de los genes es simplemente codificar la integración de las proteinas. Propiamente dicho, no hay genes para el color de los ojos. O la forma de las semillas; esto es consecuencia fenotipica de proteína codificada por un gen particular. Casi siempre las proteínas mismas actúan como enzimas: sustancias que controlan las reacciones químicas en el organismo. Aunque la proporcionen que los genes se traducen en proteínas puede ser controlada por determinado número de factores, la información genética fluye en una sola dirección: del ADN hacia delante. Esta aseveración es llamada el dogma central de la genética molecular. Se ha elaborado a partir de un enorme circulo de información experimental, y parece poco probable que sea seriamente refutada. Esto significa que las características adquiridas no son heredadas, ya que no hay modo de que la información adecuada alcance a las moléculas del ADN y cambie la estructura genética. Sin lugar a duda, se puede decir que el lamarkismo –tal como se definió anteriormente- ha muerto y no resurgirá.
Espero que estos detalles de uno de los temas más fascinantes emanados de la biología moderna, permitan al lector comprender mejor el fenómeno de las mutaciones. Las mutaciones ocurren, generalmente, por la perdida o adición de una base, o por la sustitución de una base por otra, durante la reproducción de una molécula del ADN. Además, un segmento completo del ADN puede perderse o invertirse. Es muy sencillo ver por qué las mutaciones son casi siempre nocivas, ya que las interacciones de las enzimas en el organismo están tan finamente equilibradas que cualquier cambio tiene pocas probabilidades de ser benéfico. Sin embargo, ocasionalmente las mutaciones, con nuevas enzimas, pueden ser de utilidad al organismo. Un ejemplo reciente, que tiene particular
importancia para el hombre, es el desarrollo de las bacterias resistentes a la penicilina.
La penicilina se desarrollo inicialmente durante la segunda guerra mundial, y tuvo gran éxito en el tratamiento de amplia variedad de infecciones bacterianas. Todavía es muy útil, pero en el curso de los últimos veinte años, diversas clases de bacterias han comenzado a producir una enzima que invalida la acción de la penicilina y, en consecuencia, se han hecho relativamente resistentes a este antibiótico en particular. Como la bacteria se puede reproducir cada veinte o treinta minutos, no toma mucho tiempo para que una forma mutante reemplace a la forma original, si el medio se mantiene constante. Así pues, en los hospitales, donde es probable que el medio bacteriano contenga antibióticos, es común encontrar formas resistentes. En otros medios, donde es poco probable que las bacteria entren en contacto con los antibióticos, las formas resistentes son menos frecuentes. No es solo la mutación, sino la relación entre la mutación, con sus consecuencias genotípicas, y el ambiente, lo que determina que las aptitudes de un organismo mutante sean disminuidas, incrementadas o invariables.
Este ejemplo de la selección natural en las bacterias nos da una buena idea de las acciones moleculares. La genética molecular también permite a los científicos estimar grados de cambio evolutivo. La hemoglobina – la proteína que conduce oxigeno en la sangre- ha sido ampliamente estudiada desde este punto de vista, ya que se conoce la estructura de la molécula de la hemoglobina de muchas especies de vertebrados. Ésta varia de una especie a otra, y la comparación de diferentes secuencias de aminoácidos nos proporciona una estimación de la cercanía genética de dos especies. Como se podría esperar, la hemoglobina humana es mucho más parecida a la del chimpancé que a la de un pez o un reptil. Comparando cuidadosamente las estructuras de la hemoglobina de diferentes especies, se ha llegado a estimar que esta ha cambiado su secuencia a una escala aproximada de una sustitución por colocación de aminoácidos cada 1000 millones de años, y que esa escala es bastante constante. De manera que podemos contar el número de cambios que han ocurrido en diferentes especies y obtener una estimación de las afinidades entre ellas, lo que puede servir para analizar las genealogías derivadas de los estudios de anatomía comparada y de otros testimonios la importancia potencial de una

135
pequeña mutación ubicada en una sola parte del filamento del ADN (llamado mutación puntual) está ilustrada en la hemoglobina –S humana, que es la hemoglobina de la anemia de células falciformes. Ésta es una enfermedad incapacitante y frecuentemente mortal. La hemoglobina –S está codificada por un gen recesivo que representado por el símbolo Hbs. Como el gen es el recesivo, la anemia de células falciformes sólo se presenta en individuos homocigóticos del gen (Hbs Hbs). Se le conoce como anemia de células falciformes porque los glóbulos rojos de la sangre se colapsan cuando disminuye la concentración de oxigeno y su imagen es falciforme bajo el microscopio. El individuo heterocigoto (HbaHbs) no sufre esta anemia, pero da formas falciformes en sus células sanguíneas si la concentración de oxigeno es muy baja, la diferencia entre la hemoglobina normal y la falciforme es solamente un aminoácido, resultado de una mutación puntual en el gen que codifica la hemoglobina. Sin embargo, las consecuencias para el homocigoto (Hbs Hbs) pueden ser mortales.
Los pequeños cambios genéticos pueden tener efectos mortales; pero surge la interrogante:¿por qué este rasgo o característica es tan común en algunas partes de África? Hay pueblos en África occidental de heterocigotos (Hba Hbs) para la hemoglobina anormal. Si la selección natural operara con la precisión que afirmaba Darwin, podríamos esperar que un gen tan nocivo ya hubiera sido eliminado.
Pero la clave se encuentra en la malaria, una grave enfermedad parasitaria transmitida por mosquitos que se reproducen en todas la área donde es común el gen de hemoglobina-s, por ejemplo en África occidental, donde la malaria es casi general. Éste parásito entra al torrente sanguíneo cuando el mosquito pica, se reproduce con rapidez en individuos normales y causa los síntomas clásicos de la malaria, que pueden ser fatales. Pero en individuos heterocigóticos por la característica de la célula falciforme, el parásito es incapaz de reproducirse exitosamente y los síntomas son mucho menos severos,. Ha menudo se ha dicho que el heterocigoto es resistente a la malaria, porque sus glóbulos rojos se colapsan cuando los invade el parásito, pero no es así de sencillo. La verdadera explicación puede ser que la forma mutante de hemoglobina no puede ser digerida por el parasito y, por lo tanto, los heterocigóticos (Hba Hbs) no desarrollan la malaria.
Una vez más vemos como la aptitud esta relacionada con el medio, pues en regiones
donde se presenta la malaria, ser heterocigoto representa una ventaja sobre el homocigoto (Hbs Hbs) con células falciformes y el homocigoto (Hba Hba) con células normales. En regiones donde no es frecuente la malaria, el gen Hbs no seria seleccionado ya quien no representaría ventaja alguna para su portador, y si seria nocivo cuando se presentara en condición homocigótica. Esto ha sucedido entre la población negra de los Estados Unidos. La mayor parte de los esclavos que llegaron a dicho país provenían de la costa occidental de África, y posiblemente tuvieron una alta incidencia de hemoglobina –S. Como Norteamérica no es una región propensa a la malaria, la frecuencia del gen Hbs ha disminuido, aunque todavía cerca del cinco porciento de los norteamericanos de origen africano muestra rasgos falciformes heredados de sus ancestros.
El aumento en la resistencia a la malaria de individuos que poseen un gen de hemoglobina –S ha dado como resultado dos grupos distintos (Hba Hba y Hba Hbs) dentro de cierta población humana. Se dice que estos grupos diferentes, en una misma especie, constituyen el polimorfismo. Las agrupaciones para clasificación común de la sangre humana (A,Ab,B,O), representan otro ejemplo de polimorfismo en el hombre. Un caso de polimorfismo aun más claro aparece en la mariposa nocturna, Biston Betularia, que se encuentra en las islas británicas. En las regiones industriales de Inglaterra ha habido un incremento en la frecuencia de una forma mutante, negra o melanica, de esta mariposa. Los mutantes negros, que eran muy raros y se observaron por vez primera en 1849, para finales del siglo XIX se habían vuelto de lo más común en el área altamente industrializada de los alrededores de Manchester y Liverpool. La especie ordinaria es blanca-plateada, moteada con negro; anteriormente había estado muy bien protegida contra las aves depredadoras entre los líquenes adheridos a los troncos de los árboles donde permanecía oculta durante el día.
Con la contaminación industrial del aire, los líquenes murieron y el tronco de los árboles ennegreció con el hollín, lo que ha ocasionado que la variedad melanica esté ahora mejor protegida que la especie clara. Una mayor depredación en la especie clara que en la melanica dio como resultado un aumento gradual del mutante y una disminución del original. Esta respuesta a la contaminación se ha observado en númerosas clases de

136
mariposas, en otros insectos y en algunas arañas. El melanismo industrial, como se le conoce, ha sido también en ciertas partes de Norteamérica y en la Europa continental, sobre todo en el valle de Ruhr en Alemania. Con la implantación de leyes contra la contaminación en la Gran Bretaña, la variedad melanica ya no es tan frecuente y las especies claras han vuelto a aparecer en distintas partes.
El auge de la variedad melanica de la mariposa nocturna es uno de los mejores ejemplos que tenemos de la evolución por variación combinada con selección natural, en un medio cambiante. Irónicamente, este ejemplo de la selección natural en acción –justo lo que Darwin necesitaba- tenia lugar mientras que El origen de las especies se editaba varias veces, pero nadie se percató de ello en aquel momento.
La formación de un poliformismo en determinadas especies esta muy lejos de la especiaciòn (formación de las especies), aunque Darwin siempre insistió en que una variedad permanente debería considerarse como una especie en formación. La genética moderna nos ha dado una mejor idea de la especiaciòn de la que Darwin tenia, aunque él señaló con gran percepción la importancia crucial del aislamiento en el proceso. El aislamiento geográfico, como una fuerza de la evolución, estaba arraigado en Darwin desde las experiencias que obtuvo en las islas Galápagos, las que visito durante su travesía a bordo del Beagle. A pesar de que las islas se encuentran solamente a unos 800 kilómetros de la costa occidental de América del sur, tienen flora y fauna únicas. Es mas, algunas especies varían marcadamente de una isla a otra. El gobernador de las Galápagos podía identificar las islas de origen de cualquier tortuga gigante que allí se encontrara, simplemente por la forma y marcas de su concha.
Las muchas especies de pinzones tenían variaciones sorprendentes, sobre todo el tamaño y forma de su pico, característica obviamente relacionada con la forma en que cada especie obtenía su alimento. Como las islas Galápagos son de origen volcánico y geológicamente reciente, su aislamiento del continente americano (por aguas profundas y corrientes rápidas) y de las islas entre sí demostraron palpablemente a Darwin cómo se podía desarrollar una nueva comunidad. El aislamiento geográfico no solo previene la dilución de los genes mutantes que se presentan entre grupos aislados; también puede exponer el grupo a nuevas condiciones
ambientales donde sean favorecidas nuevas variaciones. Muchos biólogos de la actualidad piensan que, casi siempre, las nuevas especies se desarrollan a partir de pequeñas poblaciones que, por alguna razón, se han aislado del grupo progenitor. Ernst Mayr ha sido un instrumento muy importante en el establecimiento de la teoría de la especiaciòn que se conoce como especiaciòn alopátrica (allopatric speciation), que significa aproximadamente “ en otro lugar”. Los grupos pequeños y aislados encuentran nuevas presiones ecológicas y las mutaciones favorables pueden esparcirse rápidamente. Mayr cree que, bajo condiciones propicias, la formación de especies puede ser un proceso relativamente rápido. Existe mucha evidencia de campo que apoya la importancia de la especiaciòn alopátrica, y la velocidad del proceso puede ser útil para explicar por qué son tan raros los fósiles de transición o intermedios.
Existen otros mecanismos por los que puede ocurrir la reproducción aislada, algunas veces sin necesidad de la separación física de los grupos. Esos mecanismos pueden comprender la variación de las épocas (las especies cercanas pueden cruzarse en distintas épocas del año), las complejidades de la conducta (los elaborados rituales durante el cortejo son uno de los medios importantes por los que se reconocen las especies), los factores mecánicos (los órganos reproductores de las especies cercanas están estructurados para evitar la copulaciòn), las preferencias ecológicas (las especies relacionadas entre sí prefieren diferentes ambientes, tal como los insectos que comen hojas de diversas especies de plantas y raramente entran en contacto aunque habiten una misma región), o las incompatibilidades fisiológicas (la cría de cruzas intraespecíficas es débil o el embrión no se desarrolla más allá de cierta etapa; alternativamente, los híbridos intraespecificos son saludables pero estériles o de poca fertilidad, de manera que resultan incapaces de reproducirse).
Darwin reconoció tales mecanismos pero no creyó que podrían ser favorecidos por la selección natural. Atribuyó todos los casos de aislamiento reproductivo alas diferencias que se `presentaron incidentalmente entre las especies cuando aparecieron las divergencias entre ellas. En parte estaba en lo cierto, la esterilidad de los híbridos como la mula, es causada por una divergencia en la estructura y el número de las cromosomas de sus progenitores, el caballo y el asno. La mula es, en sí, muy fuerte pero causa de que los

137
cromosomas no hacen pares de manera ordenada, la meiosis no procede regularmente. Los gametos producidos tienen o muchos o muy pocos cromosomas y, por lo tanto, la mula es estéril.
Ahora se ha visto que, en tal situación, la selección natural puede favorecer un mecanismo aislante que actuó anteriormente en el proceso reproductor para prevenir que se formara el híbrido estéril. Supongamos que dos especies, que pudieran cruzarse pero que produjeran híbridos estériles o de poca fertilidad coexistieran naturalmente: cada vez que el macho y la hembra de diferente especie se cruzaran, su esfuerzo reproductor seria inútil en términos evolucionistas, ya que el híbrido, siendo estéril, no propagaría sus genes. Si apareciera un mutante que fuera discriminante y que solamente se cruzara con los dos de su propia especie, tendería a transmitir sus genes, incluyendo el gen “discriminador”, a las siguientes generaciones y todas sus crías serian fértiles. La selección natural favorecería al mutante discriminador y, con el tiempo, la mutación se extendería a la población. La especiaciòn es un fenómeno genético y, como hemos visto, uno de los medios en que nuestro conocimiento de la genética –mendeliana y molecular- ha ampliado la apreciación que tenemos la visión de la vida elaborada inicialmente por Darwin. Pero ¿dónde comenzó la vida? Para Darwin, casi no existían fósiles anteriores al cambriano. (Él pensó que había presentes númerosos fósiles de una pequeña criatura, a la que llamó Eozoon, en ciertos estratos anteriores al periodo cambriano en Canadá, pero posteriormente se demostró que tan solo eran formaciones cristalinas.) Sin embargo, él afirmo su creencia de que la vida se origino en una sola fuente o en unas pocas formas simples. Tanto la lógica de su teoría como sus profundas convicciones, presuponían la existencia de la vida antes del periodo cámbrico, con su fauna abundante y variada, la que incluye representantes de casi todos los invertebrados de nuestros días.
Hasta hace poco, el aparentemente súbito descubrimiento de fósiles en el cámbrico era motivo de preocupación para los paleontólogos. sIn embargo, recientemente se han llevado estudios micro paleontológicos de gran precisión que revelan númeroso fósiles unicelulares en rocas muy anteriores al precámbrico, y en el cámbrico superior se han encontrado fósiles de algunos organismos multicelulares simples, tales como gusanos y medusas. (Todavía es inexplicable porque los organismos unicelulares que existieron como
tales hace 2000 o 3000 millones de años, dieron lugar a númerosas criaturas diversas del cámbrico en un lapso de solamente 200 millones de años; tal fenómeno quedo encerrado en la mayor explosión de actividad en los últimos 50 millones de años de dicho periodo. La llamada “explosión cámbrica” es un problema al que volveremos después.) Esta investigación ha ampliado el tiempo de vida sobre la Tierra de 570 millones de años (la edad de los primeros depósitos cámbricos) a cerca de 3350 millones, ya ha arrojado nueva luz sobre los primeros seres vivientes.
Las células más primitivas no tenían núcleo y los científicos las llaman procariontes. Existen dos grupos de procariontes, la bacteria y el alga azul-verde. Todas las plantas y animales superiores, hongos, protozoarios y otras algas diferentes a la azul-verde, son eucariontes. Dentro de cada célula hay un núcleo circundando por una membrana que contiene los cromosomas y otras diversas estructuras de las que carecen los procariontes.
Mientras que estos últimos pueden intercambiar pequeñas porciones de material genético de otras células entre sí, los eucariontes tienen un proceso eficiente y regular para el intercambio genético: la reproducción sexual. La evolución de los eucariontes a partir de los procariontes fue uno de los acontecimientos más significativos en la historia de la tierra.
La vida se origino sin duda en un ambiente carente de oxigeno, y algunos de los primeros organismos, como el alga azul-verde, tuvieron un papel muy importante en la producción de nuestra atmósfera moderna, ya que despiden oxigeno gaseoso a través del proceso de la fotosíntesis. Llevó cerca de mil millones de años para que estos organismos microscópicos transformaran la atmósfera en un ambiente rico en oxigeno, como el que ahora respiramos. Los cambios atmosféricos efectuados por tales organismos primitivos ilustran una generalización biológica muy importante: las plantas y los animales no solo existen en el medio, también lo transforma. La Tierra no es únicamente el escenario donde se representa la evolución, también es actor en el drama.
Gran parte de nuestro conocimiento sobre los primeros sucesos de la evolución debe ser obtenido mediante una cuidadosa deducción. Una serie de experimentos realizados por Stanley L Miller y Harold C Urey, simulando lo que pudo haber sido la condición del planeta primitivo, han demostrado que muchas de la unidades orgánicas básicas de la vida pueden

138
producirse a partir de una mezcla de agua, metano, amoniaco e hidrógeno. Las descargas eléctricas (semejantes a las producidas por los rayos) aplicadas a la mezcla, aislada herméticamente de la atmósfera, provén númerosas sustancias orgánicas más complicadas, tales como los aldehídos, los ácidos carboxílicos y los componentes básicos de la proteína; los aminoácidos. Por supuesto que hay una gran distancia entre Este “caldo orgánico” y la célula viva y, hasta ahora, nadie ha podido crear vida en el laboratorio. Pero tanto los experimentos de Miller y Urey como la investigación básica de la bioquímica, desarrollada durante las dos ultimas décadas, han explicados algunos de los probables pasos en el inicio de la evolución bioquímica.
Es muy importante tomar en cuenta que la síntesis de estos compuestos orgánicos solamente fue posible en una atmósfera carente de oxigeno.
Además, las radiaciones ultravioleta, así como los relámpagos o rayos, fueron una fuente importante de energía para la síntesis que se llevo a cabo.
Dado que el oxigeno (O2) se acumulo en la atmósfera, una capa de ozono (O3) se desarrollo por enzima de él, y actualmente el ozono filtra la mayor parte de la radiación ultravioleta que emana del sol. Por estas razones, entre otras, un organismo viviente no podría producirse de novo hoy día en la Tierra.
Pero en la época de Darwin se creía que la “generación espontánea” era posible desde el momento que pequeñas criaturas se observaban bajo el microscopio en soluciones en las que se les había añadido alguna sustancia vegetal. De hecho, Lamarck y otros de los primeros evolucionistas, como Robert Chambers, se apoyaron en la generación espontánea para sustentar el origen de los organismos primarios. Ellos consideraron el progreso evolutivo como inevitable: una fuerza inexorable que empujaba a cada criatura hacia niveles cada vez más altos de perfección, y por esa razón sus teorías no acataban la existencia continuada de las “formas primitivas” en el preciso momento en que fue publicado El origen de las especies, Louis Pasteur llevaba a cabo sus experimentos que demostraban que la generación espontánea no ocurría, pero sus resultados no representaron un problema para Darwin, quien no veía razón alguna para que un organismo primario persistiera sin cambios, estando bien adaptado.
Volviendo al problema de la “explosión cámbrica”: se han elaborado varias teorías para explicar el fenómeno. Hay quienes sostienen que la “explosión” es ilusoria, y que la primera diseminación de los invertebrados tuvo lugar durante el precámbrico, pero en una localidad restringida, o en donde la fosilización era imposible. A esto siguió una rápida migración, debida a un cierto cambio en las condiciones. Otro grupo de teorías invoca un cambio en el ambiente que activo la rápida diseminación de formas. La más plausible de estas teorías sostiene que la concentración de oxigeno solo logro un nivel suficientemente alto para sostener animales multicelulares a fines del precámbrico. Un tercer grupo de teorías intenta simplemente explicar la "“explosión"”en términos biológicos o ecológicos, sin hacer mención de cambio climatológico alguno. Una de ellas sostiene que esta diseminación de formas fue activada por la evolución de células eucariontes, las cuales, con los cromosomas contenidos en su núcleo, llevan a cabo una eficiente forma de reproducción sexual desconocida entre los procariontes. La presencia de este refinamiento en la reproducción sexual permitió un gran aumento en la tasa de variación genética, lo que se tradujo en una explosión de la diseminación adaptativa.
Pero, ¿cuándo se desarrollaron por primera vez los eucariontes? Algunos paleontólogos sostienen que fue mil millones de años antes de que se iniciara el periodo cámbrico; en cuyo caso, la teoría antes mencionada no es muy convincente. Pero existe desacuerdo sobre la interpretación de los microfósiles antiguos, y otros paleontólogos piensan que los eucariontes aparecieron mucho después, en el precámbrico.
Puesto que los científicos están tratando de comprender acontecimientos que tuvieron lugar hace miles de millones de años, parece que nunca conoceremos la historia completa del origen de la vida. Pero la teoría de la evolución es, sin duda, la más aceptada hoy día por las ciencias naturales.
Las observaciones e investigaciones del mañana podrán producir lo inesperado. ¿Quién pudo haber predicho la existencia de un celacanto viviente? Esta especie de pez se creía extinta desde hace 100 millones de años. Por otro lado la magnitud y variedad de las fenómenos explicables por la teoría de la evolución, es asombrosa. Darwin inicio un programa de investigación multidisciplinario que, durante el siglo pasado, dio respuesta a muchos interrogantes que se suscitaron con El origen de la especies. Pero la biología

139
evolutiva contemporánea aun sigue sin dar respuesta a muchos puntos de controversia; y ahora mencionaremos algunos de ellos.
INVESTIGACION Y CONTROVERSIA: BIOLOGÍA EVOLUTIVA ACTUAL
Al bosquejar un siglo de biología darwiniana, puede haber dado la impresión de que la mayoría de los asuntos de la evolución han quedado minuciosamente resueltos. No es cierto, pues cada rama de la ciencia cambia constantemente, y la controversia proporciona vigor y emoción al que hacer científico. Sin embargo, hemos vistos varios aspectos que parecen haber determinado sin duda alguna: la realidad de la evolución; el ADN como sustancia genética; y el dogma central de la genética molecular. Cada una de estas áreas del saber tiene, por supuesto, preguntas sin respuesta, pero cada una contiene su parte medular de testimonios en los que podemos confiar.
En la actualidad se llevan a cabo investigaciones sobre la importancia de la evolución en cada una de las disciplinas que Darwin exploro para probar su teoría. Anatomía, embriología, fisiología, geología, paleontología, genética, taxonomia y ecología: todas tienen algo que aportar a la teoría de la evolución. Tomemos unos cuanto ejemplos. La teoría del desplazamiento continental sostiene que las actuales masas terrestres estuvieron alguna vez conectadas entre sí. Existe gran evidencia que lo sugiere y de que todavía los continentes se separan lentamente. Esta teoría tiene consecuencias importantes para explicar las observaciones que hicieron los geólogos del siglo XIX sobre la similitud de los fósiles encontrados en estratos antiguos del Viejo y el Nuevo Mundo, sobre todo en el hemisferio sur. Los creacionistas lo explicarían fácilmente como resultado de la creación del mundo entero, pero actualmente es posible comprender tal fenómeno en términos de desplazamientos continentales. Asimismo, los nuevos conocimientos en las relaciones taxonómicas de las especies pueden ser recopilados con el empleo de computadoras que manejan grandes cantidades de información detallada para compararla. La ciencia de la ecología, que Darwin fue el primero en comprender en toda su complejidad, ha adquirido vigencia nuevamente, ya que el hombre cambia su medio cada vez mas. Ahora la genética molecular es una de las áreas de la investigación biológica más excitantes, y hay mucho que aprender sobre la regulación de
los genes, la expresión de los mismos y los detalles de la estructura y función genética de los organismos más complejos. Es tan fundamental la teoría de la evolución para las ciencias de la vida, que no hay investigación biológica, en cierto nivel, que no sea relevante para el tema. Mientras mayor sea el conocimiento que tengamos de los organismos, mejor comprenderemos su evolución.
Mas específicamente aún, existen cuatro o cinco temas de controversia en la biología evolutiva moderna, que deben mencionarse. Ellos son: el problema de las mutaciones neutrales, la universalidad de la selección natural como mecanismo evolutivo, la evolución del sexo, el origen del altruismo y la “sociobiologia”, o la posibilidad de una síntesis que incluya la biología evolutiva y las ciencias sociales. Brevemente consideraremos cada uno de ellos.
Ya hemos visto que la mutación lleva consigo cierto cambio en la molécula del ADN, el cual, a su vez, afecta las proteínas cuya síntesis regula. Las mutaciones producen proteínas alteradas y, en términos de la selección natural darwiniana, las benéficas son favorecidas y las nocivas son desechadas. Pero ¿qué sucede con una mutación sin consecuencias selectivas? ¿Qué pasa si una nueva molécula de proteína, con un aminoácido sustituido, lleva acabo su función con la misma eficiencia? Bajo circunstancias normales, en una población amplia y que se produce libremente, una mutación de este tipo no aumentara su frecuencia. Sin embargo, algunos biólogos sostienen que un buen porcentaje de mutaciones son neutrales, en cuyo caso esto representa una importante fuente de cambio evolutivo no darwiniano (es decir, una fuente de cambio no afectado por la selección natural), aun cuando las mutaciones neutrales no conduzcan, por si mismas, a nuevas adaptaciones.
Las diferencias entre proteínas, debidas a tales mutaciones, pueden ser detectadas con una gama de técnicas bioquímicas. Durante los últimos diez años, el emplear estas técnicas los genetistas han descubierto muchas más variaciones en las proteínas de la población de lo que se hubiera podido pensar. La existencia de más de una forma de proteína en una población es conocida como polimorfismo proteínico,. Para que Este polimorfismo se mantenga por selección natural, una parte de la población debe morir como sucede en el caso de la anemia de células falciformes, en el ejemplo de la malaria. Si todos los polimorfismos proteínicos

140
que se han encontrado se hubiera mantenido por selección natural, entonces los cálculos nos dicen que el número de individuos que tendrían que morir superaría a menudo la capacidad reproductora del organismo.
Por lo tanto, los evolucionistas han tenido que volver a la conclusión de que estas variantes en las proteínas son neutrales. De ser así, no requieren cambiar de frecuencia de una generación a otra, y todas las proteínas deben mostrar el mismo monto de polimorfismo. Pero, de hecho, algunas proteínas son mucho más variables que otras. ¿Por qué? La cuestión de las mutaciones neutrales continua siendo uno de los problemas centrales de la teoría moderna de la evolución.
Las mutaciones neutrales representan un medio para que otros factores, apartes de la selección natural, puedan afectar el agrupamiento de genes; otro lo es la deriva génica, pero esta solo opera en pequeñas poblaciones. En ausencia de la selección natural, el gen no debería cambiar de frecuencia entre una generación y la siguiente. En poblaciones grandes, deberá ser así pero en poblaciones pequeñas las frecuencias de los genes pueden cambiar debido a la casualidad. Si lanzamos al aire cien veces una moneda, es poco probable que obtengamos cien cruces y ninguna cara, pero si la lanzamos dos veces, es muy posible que recibamos dos cruces y ninguna cara. Este efecto casual, cuando la frecuencia es pequeña, es lo que produce la deriva génica.
En poblaciones pequeñas, tanto humana como de otros organismos, los efectos de deriva génica pueden observarse a menudo. La frecuencia de los caracteres en tales poblaciones pueden diferir considerablemente de la frecuencia con que aparecen los mismos caracteres en grandes poblaciones, de las cuales se han aislado. Por ejemplo: en grandes núcleos de población humana son raros la mayoría de los trastornos genéticos; pero en la comunidades religiosas cerradas, como es el caso de la comunidad Amish Pensilvania, en los Estados Unidos, un mismo trastorno genético, como el síndrome de Troyer (una forma de distrofia muscular), es bastante común. En estos casos, casi siempre las comunidades han sido fundadas por un reducido número de individuos, algunos de los cuales pudieron ser portadores de raros genes recesivos, en condición heterocigótica. En poblaciones pequeñas derivadas por completo de sus fundadores, tales genes se vuelven comunes por las probabilidades del desplazamiento genético, y los individuos homocigotos por los genes recesivos y que
por tanto exhiben la característica recesiva, son más frecuentes que en el mundo exterior, los trastornos genéticos son solamente las manifestaciones más obvias de los cambios de las frecuencias genéticas en el agrupamiento de genes en una población pequeña.
El debate sobre las mutaciones neutrales y la deriva génica forma parte del argumento entre quienes piensan que todas las características que un organismo muestra tienen un significado adaptativo, y los creen que algunas estructuras son simplemente fortuitas o que resultan de un desarrollo limitado, del cual no siempre estamos conscientes. Un ejemplo de esto último es el mentón humano. Esta conformación que no tiene ninguno de los simios superiores, era una incógnita para los biólogos, hasta que se descubrió que no tenia ningún significado adaptativo. El mentón es el resultado de diferentes grados de crecimiento que operan en las márgenes superior e inferior de la mandíbula inferior. Esos grados de crecimiento han disminuido en relación con nuestros ancestros, pero el crecimiento del borde superior se ha reducido mucho más y ha dado como resultado una protuberancia en el margen inferior que es lo que llamamos mentón. Este es un ejemplo de un fenómeno general que se conoce como alometria.
Como ejemplo de argumento sobre el significado adaptativo, está el que se refiere a los variados y complejos patrones que se observan en las conchas de muchos moluscos, que para algunos evolucionistas no son adaptativos. Sin embargo, la importancia funcional de cualquier carácter puede no ser obvia, a menos que la conducta y las relaciones ecológicas del organismo se comprendan cabalmente. Se pensó por mucho tiempo que las franjas de patrones polimórficos y la coloración de ciertas especies de caracoles de tierra. Cepea nemoralis, no tenían significado adaptativo alguno. Estos caracoles existen en color rosa, amarillo y café, con cinco, tres, una o ninguna franja negroide.
Después de observar cuidadosamente el tordo, uno de los depredadores de caracol, Philip Sheppard a Arhur Cain pudieron demostrar que esta ave tiende a capturar los caracoles más llamativos, y que en algunos lugares una clase está mejor disimulada que otras. Por ejemplo, en pastizales, los caracoles de concha café eran capturados con mayor frecuencia que los de concha amarilla, cuyo color los ocultaba; pero entre las hojas secas, en cambio, los de concha café estaban más protegidos. En los pastizales, los

141
caracoles de cinco franjas tenían ventaja, pues se confundían con los tallos de las plantas. Antes de determinar si una característica es o no adaptativa, debe estudiarse cuidadosamente la biología del organismo como un todo.
Por otro lado, los críticos de la tesis de que “todo es adaptativo”, tienen un punto a su favor cuando dicen que si uno se lo proporcione es muy fácil encontrar explicaciones adaptativas. Si cualquier característica se acepta automáticamente como adaptativa, no es difícil, en la mayoría de los casos, construir explicaciones plausibles. Darwin mismo reconoció esta trampa cuando escribió:
Si solo hubiera existido pájaros carpinteros verdes y no se conocieran muchas especies negras y de varios colores, me atrevería a decir que hubiéramos creído que el color verde era una hermosa adaptación para ocultar de sus enemigos estas aves que viven en los árboles y, en consecuencia, que era éste un carácter de importancia que había sido adquirido mediante selección natural, cuando en realidad el color probablemente es debido, en su mayor parte, a la selección sexual.
Las polémicas entre “seleccioncitas” y “neutralistas” no deben representar un retroceso a los desacuerdos del siglo XIX, cuando muchos científicos que aceptaron la evolución cuestionaron la selección natural como su principal mecanismo.
A partir del trabajo de los genetistas de poblaciones, efectuado entre los años de 1930 y 1940, el funcionamiento de la selección natural sobre las variaciones y mutaciones ha sido ampliamente reconocido como la fuerza impulsora del cambio evolutivo, y la investigación de númerosos evolucionistas contemporáneos como Ernst Mayr y Theoodosius Dobzhansky tiende a confirmarlo. Sin embargo, todavía hay biólogos que se sienten incómodos ante la idea de que la selección natural sea un concepto tan elástico.
Tanto la adaptación como la selección natural, aunque son intuitivamente fáciles de captar, con frecuencias son difíciles de estudiar rigurosamente, ya que su investigación implica no solamente relaciones ecológicas muy complejas, sino también la aplicación de matemáticas avanzadas a la genética de poblaciones. Los críticos de la selección natural pueden esta en lo cierto al cuestionar su universalidad, pero la importancia de otros mecanismos, tales como las mutaciones neutrales y los desplazamientos genéticos,
todavía es desconocida. MIentras tanto, es claro que la teoría moderna de la evolución emplea la selección natural, al igual que lo hizo Darwin, aunque la genética de poblaciones y la genética molecular han transformados el modo de comprenderla.
¿Evolucionaron el sexo y la reproducción sexual también a trabes de la selección natural? Sin lugar a dudas la respuesta es afirmativa. Pero como probablemente la reproducción sexual se desarrolló hace 1000 o 2000 millones de años. Muchos detalles son desconocidos. En general, la reproducción sexual es la característica que separa a los eucariontes de los procariontes, aunque ocasionalmente ciertos procariontes intercambian información genética entre sí, y algunos eucariontes se reproducen asexualmente. En términos evolutivos, la reproducción sexual es de suma importancia ya que númerosas especies le dedican mucha energía. La ventaja de la reproducción sexual es obvia, ya que propicia una gran variabilidad. Un organismo asexual simplemente produce copias idénticas a si mismo, por lo que su potencial para el cambio es limitado, y éste solamente se puede presentar por la vía de la mutación genética. En este marco, el logro de dos mutaciones favorables por un solo individuo requiere dos acontecimientos independientes, mientras que las especies que se reproducen sexualmente, las probabilidades de que los genes se encuentren por medio de intercambio genético son mucho mayores.
Por otro lado, un individuo femenino de una especie, que optara por la reproducción asexual, de inmediato aumentaría aproximadamente al doble si adecuación genética. Es decir, todo su esfuerzo reproductor lo dedicaría a reproducir sus propios genes, en lugar de que la mitad de ese esfuerzo se empleara en los genes de la pareja. Así que, dicho de otra forma, un individuo transmitiría el doble de copias de sus genes a la siguiente generación. Por lo tanto, se podría esperar que los individuos femeninos de una especie bien adaptada, se inclinaran con frecuencia por la reproducción asexual o partenogénesis (y que las poblaciones se convirtieran en básicamente femeninas). El hecho de que esto no suceda, confirma la naturaleza siempre cambiante del medio, que requiere que las especies se adapten constantemente a circunstancias ligeramente diferentes; esto conoce como seguimiento ambiental; dicho de una manera coloquial, como la hipótesis de la Reina Roja (en honor del personaje de Alicia en el país de

142
las maravillas, quien tenia que mantenerse corriendo para permanecer en su sitio).
El individuo que mantenga su cría aislada del agrupamiento genético, reproduciéndose por partenogénesis, la destinará a la extinción, ya que sin la variabilidad que proporciona la reproducción sexual no podría seguir al mismo ritmo de su entorno.
Muchos animales invertebrados se reproducen, a veces, por partenogénesis, pero este proceso generalmente se alterna con la reproducción sexual. Los pulgones, por ejemplo, se reproducen asexualmente durante el verano, cuando existe gran provisión de alimentos. Entonces nacen pulgones hembras sin alas, que rápidamente dan origen a nuevas hembras sin alas, lo que se traduce en una explosión de la población. Los machos solamente nacen en el otoño, cuando tiene lugar la reproducción sexual; se ponen los huevecillos, los que se conservan durante el invierno, y emergen en la primavera siguiente. Hay algunas plantas en la reproducción sexual ha perdido importancia, pero sucede en unas cuantas, como las zarzas, especie de Rubus, y el diente de león, especie de Taraxacum, que son altamente eficientes y cuentan con medios para expandirse vegetativamente.
Darwin argumentaba que la selección natural actúa solo en el individuo (aunque hizo una excepción de los insectos sociales). Los biólogos de hoy tienen una opinión similar, pero en el ínterin surgió una teoría bastante confusa, que sugería que la selección natural podía presentarse en las especies; sin embargo, fue aceptada por algunos y se volvió muy popular. Con esta teoría se pretendía explicar lo que podrían ser actos altruistas de un animal hacia otro, o hacia un grupo. En la versión popular de la teoría, se decía que esos actos eran “por el bien de la especie”.
Tales explicaciones se descartan con solo pensar un poco: un ave hembra no protege ni alimenta a sus cría “por el bien de la especie”, sino por la propagación de sus propios genes. (Se debe tener en cuenta que este no es un motivo, sino que tal ave no tuviera genéticamente determinado el “instinto materno”, sus genes se extinguirían rápidamente, pues sus crías no sobrevivirían). De igual manera, las aves no limitan el número de huevecillos que ponen para prevenir la explosión de la población, “por el bien de la especie”. Si ponen solo cuatro huevecillos es porque serian incapaces de recolectar comida suficiente para cinco aves
recién nacidas, y todas o casi todas morirían de hambre; por lo tanto, no llevarían adelante la transmisión de los genes de sus progenitores.
Otros casos de altruismo no pueden ser pasados por alto. En las hordas de mandriles, un miembro no puede arriesgar su vida en defensa del grupo.
En ciertas especies de aves, no todos los miembros se aparean ni crían a sus pequeños; las aves que no lo hacen, pueden ayudar a su hermana a conseguir un compañero, o pueden ayudar a criar a los menores. Hay una teoría que parece explicar todas estas formas de comportamiento en términos darwinianos: la selección familiar. Esta teoría, en su forma más simple, afirma que con un hermano se comparten los genes (la mitad) como un hijo; de modo que, ayudando al hermano a sobrevivir y a reproducirse, se ayuda a perpetuar los propios genes (dicho de otra manera, se aumenta la adecuación genética), tal como si se protegiera al propio hijo. El grado de parentesco de cualquier pareja de individuos es calculado según el porcentaje de genes que tiene en común, y se han hecho mediciones para ver hasta donde un individuo debe mostrar “altruismo” hacia otro. La teoría es atractiva, pero hay trabajos que indican que el parentesco no puede ser equiparado con el avance en la aptitud genética alcanzada por un hecho altruista tan sencillamente como se ha sugerido, y que se requiere de un enfoque matemático mucho más complejo que tome en cuenta, por ejemplo, la edad y el potencial reproductor del parentesco para su determinación. Para poner un ejemplo sencillo, se puede alcanzar muy poca capacidad genética al rescatar a un hermano mayor que por sí solo tiene posibilidades de sobrevivir.
Uno de los casos de altruismo más difíciles de explicar es el del ave que da un grito de alarma cuando se acerca un gavilán. Es te grito permite que la banda escape al mismo tiempo, confundiendo al gavilán y reduciendo el número de pájaros capturados. Pero el pájaro que dio el grito se hace notorio y, por lo tanto, tiene mayores posibilidades de que lo atrapen. Solamente si un gran número de aves de la bandada esta emparentando con él, puede explicarse esta conducta como selección de parentesco.
Como Darwin observo, los insectos sociables son un caso excepcional. La obrera estéril se puede decir que realmente trabaja por el bien de la colonia. La selección natural funciona en la colonia, vista ésta como un todo; por lo

143
tanto, la colonia se concibe de mejor manera como un hiperindividuo, en el cual cada miembro actúa en forma análoga a las células del cuerpo; puesto que las obreras son irreversiblemente estériles, sus actividades no pueden considerarse como “altruistas”, ya que el auto sacrificio no es posible si no hay algo que sacrificar. La verdadera incógnita en relación con estos insectos sociables la encontramos en la etapa primaria de su evolución.
El abuso del termino “altruismo”, en este contexto, y la atención desmedida que se le ha dado al debate, son indicativos de lo que está realmente en juego. ¿Es el altruismo entre los seres humanos simplemente una expresión más de la evolución de funciones? Dicho de otra forma, ¿trascienden los valores humanos el mundo de la selección natural, o es posible entender nuestro comportamiento a trabes de una juiciosa combinación de biología evolutiva, la etologia (el estudio de la conducta animal) y las ciencias sociales? él biólogo de Harvard, Eduard O. Wilson, es el principal patrocinador de la tesis de que la biología y las ciencias sociales se pueden amalgamar en lo que él llama “sociobiologia”.
Wilson afirma que como algunos de los rasgos de la conducta humana son universales, la transmisión cultural no puede ofrecer una explicación cabal. Él sostiene que algunas conductas, como el rechazo al incesto, pueden tener una base genética. Las ideas de Wilson han provocados violentas criticas y ciertamente existe el peligro de extrapolar la conducta animal a la humana, o viceversa, a la manera poco critica que caracteriza mucha de la “sociobiologia”. Uno se debe preguntar si se puede hablar de conducta de los animales en términos humanos, los que han sido ampliamente usados por los sociobiólogos, tales como “fidelidad” y “coqueteo”, sin prejuiciar inmediatamente nuestras conclusiones a favor de la analogía implícita. Ciertamente que hay muchas fallas, pero también hay mucho que tomar en cuenta. Un planteamiento más riguroso, y una discusión menos apasionada por parte de quienes están en desacuerdo son necesarios.
Por lo pronto, esto es cierto: todos los aspectos de la moderna biología evolutiva se pueden considerar como parte del programa de investigación iniciado con El origen de las especies, sin duda el trabajo sobre biología más importante jamás escrito.
CUESTIONARIO
1.- ¿POR QUÉ, A PESAR DE QUE DARWIN NO FUE EL PRIMERO EN HABLAR DE EVOLUCION, SU TRABAJO MARCA EL ORIGEN DE LA CONCEPCION MODERNA DE EVOLUCION?
2.- ¿CUÁLES FUERON ALGUNOS DE LOS PUNTOS QUE DARWIN NO LOGRO EXPLICAR ADECUADAMENTE EN El ORIGEN DE LAS ESPECIES? ¿CÓMO SE RESOLVIERON POSTERIORMENTE?
3.- ¿QUÉ RELACION EXISTE ENTRE LOS TRABAJOS DE DARWIN Y MENDEL?
4.- DISCUTA EL SIGNIFICADO DE LAS MUTACIONES PUNTUALES DENTRO DEL PROCESO DE SELECCIÓN NATURAL Y EVOLUCION, Y LA VALIDEZ DE LA SELECCIÓN NATURAL COMO UNICO MECANISMO EVOLUTIVO.

144
CAMBIO CONCEPTUAL Y METODOLÓGICO EN LA
ENSEÑANZA Y EL APRENDIZAJE DE LA EVOLUCION DE LOS SERES
VIVOS UN EJEMPLO CONCRETO
GENÉ, A.
DEPARTAMENT DE DIDACTICA DE LA MATEMÁTICA I DE LES CIENCIES EXPERIMENTALS.
UNIVERSITAT AUTÓNOMA DE BARCELONA. CAMP DE MART, 33, 25004 – LLEIDA.
SUMMARY
In this work we developed and assessed the efficiency of a didactic lesson based on the science teaching-learning paradigm as a conceptual, methodological and attitudinal change.
For this purpose we chose the topic of the evolution of living beings through natural selection.
INTRODUCCIÓN-PLANTEAMIENTO DEL PROBLEMA E HIPOTESIS DEL TRABAJO
Desde el trabajo de Lucas (1971), en el que por primera vez se constata la similitud entre las explicaciones de los alumnos respecto a la evolución de los seres vivos con las de Lamarck, las investigaciones sobre las preconcepciones referidas ha este tema han ido aumentando. Se ha mostrado la coincidencia de estas ideas en estudiantes de muchos países lejanos entre sí, tanto geográfica como culturalmente. Así, Jungwirt (1975) encuentra ideas lamarckianas en escolares de Israel; Brumbi (1980), en escolares Ingleses; Hallden (1988), en adolescentes suecos; Kinnear (1983) y Martín (1983), en estudiantes de Australia.
En nuestro país, dos recientes trabajos (Alvadalejo y Lucas 1988 y Jiménez Aleixandre 1989) han estudiado las características de los pensamientos de los alumnos sobre este tema. El primero muestra las concepciones sobre la mutación entre los estudiantes de BUP. Jiménez Aleixandre en su interesante tesis doctoral, además de estudiar las concepciones de los alumnos, propone un modelo de enseñanza / aprendizaje para salir al paso de los errores.
Nuestro trabajo no pretende mostrar que hay concepciones incorrectas. Nuestro estudio plantea hasta que punto un determinado modelo didáctico puede favorecer o no la superación de estas preconcepciones.
En nuestra opinión, el hecho que los estudiantes expliquen las modificaciones de los seres vivos de la misma manera que lo hizo Lamack, no puede ser debido a la casualidad. No puede explicarse por el azar. Pensamos con Gil (1983; Gl Carrascosa. 1985) que la coincidencia de las ideas es debida a la coincidencia en la metodología utilizada para aproximarse a la realidad. Una metodología que no va más allá de la evidencia, de la simple observación y de la inducción de conclusiones a partir de la misma.
La utilización de instrumentos de aprendizaje de estas características lleva a pensar que un órgano crece porque se usa mucho, y se atrofia si no se usa. Que estas características pasen de padres a hijos también es fácil de explicar, con el sentido común, por la herencia de los caracteres adquiridos, como lo hizo Lamarck. Veamos algún ejemplo.
Planteamos a alumnos de 14 años la siguiente cuestión. (Grau 1989):
Las aves acuáticas que nadan por la superficie del agua utilizan las patas para impulsarse tienen una membrana interdigital desarrollada, como la de la figura, que hace más eficaz el movimiento de la pata, y por lo tanto, su desplazamiento.
Explica como crees que se ha originado esta membrana.
Respuesta Escolar: yo pienso que las aves acuáticas van desarrollando esta membrana a lo largo de su vida hasta llegar a adultos. Quiero decir que como los patos desde pequeños están en contacto con el agua, el movimiento continuo de sus patas hace que la membrana vaya siendo cada vez más fuerte. Si no se encontrara en contacto con el agua, esta membrana desaparecería ya que no tendría ninguna utilidad. Los patos pequeños heredan de sus padres esta membrana.
Respuesta de Lamarck: Filosofía Zoológica (1809).
...En todo animal que no haya llegado al final de su desarrollo, el uso frecuente y sostenido de un órgano, lo fortifica, lo desarrolla lo agranda y le da una potencia proporcional a la duración de uso. En cambio, la falta constante de uso de un órgano, lo debilita, lo deteriora,

145
disminuye progresivamente sus facultades y acaba por hacerlo desaparecer.
Todo cambio adquirido en un órgano por un habito de uso suficiente, se conserva y pasa a las otras generaciones si el cambio es común al macho y a la hembra...
Siguiendo con nuestro razonamiento, si la causa de exista coincidencia entre la ideas de Lamarck y nuestro alumnos está en la forma “natural” de acercarse a la realidad, será preciso modificar esta metodología para que superen las concepciones incorrectas. Pero ¿en qué sentido debe modificarse la forma de acercarse a la realidad?
Un primer requisito será hacer dudar de las evidencias, hacer dudar del sentido común. Para ello se deben plantear cuestiones o situaciones que no puedan explicarse por el pensamiento del alumno, por su propio paradigma.
El conflicto que surge con el propio pensamiento es el punto de partida para ir construyendo nuevos conocimientos pero, eso sí, con una metodología diferente que parta de este problema-conflicto. La solución a este problema ha de estar dirigida por hipótesis de trabajo cuya contrastacion vaya permitiendo destruir los errores y construir un nuevo marco conceptual, una nueva teoría.
Nuestra hipótesis de trabajo pues, se concreta en que la utilización de una unidad didáctica con estas orientaciones facilitara la superación de las concepciones incorrectas de los alumnos y la construcción de la teoría de la evolución de los seres vivos por selección natural.
Vamos a continuación a mostrar como ha sido contrastada esta hipótesis.
DISEÑO EXPERIMENTAL
El eje central del diseño experimental ha sido la confección de una unidad didáctica fundamentada en el modelo que se propone.
El hilo conductor de esta unidad –de la que se muestra algunos fragmentos en el anexo- ha sido la consideración de aspectos clave, que a nivel histórico, fueron útiles para la construcción de la teoría de la evolución por selección natural.
La unidad plantea los siguientes problemas:
1. ¿Cómo cambian los seres vivos?
2. ¿Puede variar el número de individuos de una población?
3. ¿Qué pasa cuando las condiciones ambientales cambian?
4. ¿Qué es una mutación?
5. ¿Cómo actúa el ser humano sobre las especies para sacar beneficio?
6. ¿Por qué son tan diferentes los individuos salvajes y domésticos de una misma especie?
7. ¿Cómo se forman las nuevas especies?
8. ¿Qué les paso a los dinosaurios?
9. ¿Dónde esta localizado el material genético que hace que un ser vivo modifique sus características?
10. ¿Puede explicarse la evolución humana por la teoría de la evolución por selección natural?
11. ¿Ha influido la teoría de la evolución de los seres vivos en la sociedad humana?
La unidad didáctica se trabajó con alumnos de 8º de EGB y futuros profesores de primaria.
Las actividades de la unidad pensadas para conectar con los conocimientos previos y, por tanto, para iniciar el aprendizaje, fueron utilizadas para conocer estadísticamente los preconceptos. Estos primeros resultados han sido utilizados como punto de referencia, al servir para ver hasta que punto se había conseguido el aprendizaje significativo de la evolución de los seres vivos por selección natural.
Paralelamente se ha contado con dos grupos control que han trabajado el tema durante el mismo curso con las metodologías de trabajo de aula habituales. Una metodología (grupo c,1) responde al modelo de transmisión de conocimientos elaborados. El grupo c,2 trabajó con un modelo activo o de descubrimiento autónomo. Los resultados correspondientes a los maestros de primaria en formación se han comparado con un grupo de futuros maestros de ciencias que habían estudiado este tema en BUP y COU.
Pasamos a continuación a concretar el diseño, presentando las cuestiones utilizadas, tanto en el pretest como en el postest.
Cuestión 1. Las aves acuáticas que nadan por la superficie del agua y utilizan las patas para impulsarse tienen una membrana interdigital desarrollada, como la de la figura, que hace más eficaz el movimiento de la pata y por lo tanto su desplazamiento.
Explica como crees que se ha originado esta membrana.

146
Esta pregunta (Grau 1989), basada en un problema planteado por Lamarck en su libro de Filosofía Zoológica, ha sido pasada como pretest a alumnos de 8º de EGB y futuros maestros en número aproximado de un centenar.
Cuestión 2. Cuando se utilizaron por primera vez los insecticidas aerosoles contra las moscas y los mosquitos, resultaron muy eficaces, es decir, casi todos los insectos morían cuando se ponían en contacto con el insecticida.
Hoy 30 años después, solamente una pequeña proporción de estos insectos muere cuando utilizamos los aerosoles con la misma composición.
Explica por qué crees que esto ocurre.
Esta cuestión ha estado utilizada previamente por Jiménez Aleixandre (1989), y ha servido como postest para ver hasta que punto se había conseguido el aprendizaje significativo de la evolución de los seres vivos por selección natural. Ha sido pasada a los grupos control, y a los alumnos de 8º de EGB y futuros maestros.
A continuación vamos a presentar los resultados obtenidos.
RESULTADOS-ANALISIS Y CONCLUSIONES
Cuestión 1: pretest. Las aves acuáticas que nadan por la superficie del agua y utilizan las patas para impulsarse tienen una membrana interdigital desarrollada, como la de la figura, que hace más eficaz el movimiento de la pata y por lo tanto su desplazamiento.
Explica como crees que se ha originado esta membrana.
ALUMNOS QUE DAN UNA RESPUESTA LAMARCKISTA
%
100
90
80
70
60
50
40
30
20
10
0
(1) (2)
(1) Alumnos de 8º de EGB antes de empezar a trabajar el tema con el modelo de enseñanza / aprendizaje que se propone. N=30; %=92´3: sd=4´8.
(2) Futuros maestros de primaria antes de empezar a trabajar el tema con el modelo de enseñanza / aprendizaje que propone. N=54; %=85’1; sd=48.
Cuestión 2. Cuando se utilizaron por primera vez los insecticidas aerosoles contra las moscas y los mosquitos, resultaron muy eficaces, es decir, casi todos los insectos morían cuando se ponían en contacto con el insecticida.
Hoy 30 años después, solamente una pequeña proporción de estos insectos muere cuando utilizamos los aerosoles con la misma composición.
Explica por qué crees que esto ocurre.

147
ALUMNOS QUE DAN LA RESPUESTA CORRECTA
%
100
90
80
70
60
50
40
30
20
10
0
(1) (2) (3) (4) (5)
(1)Alumnos de 8º de EGB que han estudiado el tema. N=30: %=85’7;sd=6’6.
(2)Maestros que han estudiado el tema. N=54; %=100; sd=0.
(3)Grupo de 8º de EGB 1,c metodología de transmisión. N=64; %=4’6; sd=2’6.
(4)Grupo de 8º de EGB 2.c. metodología de descubrimiento autónomo. N=30; %=3’8; sd=3’4.
(5)Maestros de ciencias en formación. Han estudiado el tema BUP y COU. N=51; %=26’3; sd=6’1.
La t de Student nos da, comparando los grupos de 8º de EGB entre sí y los grupos de futuros maestros, diferencias significativas muy elevadas, con una & superior en todos los casos al 0’01
A la luz de estos primeros resultados, podemos pensar que el modelo de enseñanza aprendizaje de las ciencias naturales como cambio conceptual y metodológico que se concreta en el programa-guía que presentamos en el anexo, parece ser útil para la superación de las errores conceptuales en el tema de la evolución de los seres vivos. De hecho, los alumnos de 8º de EGB y los futuros maestros aprendieron de forma significativa la evolución de los seres vivos por selección natural.
Hemos de indicar que los alumnos de 8º de EGB, que utilizaron el modelo que proponemos se les planteó, después de tres meses de termino el tema, dos problemas para ver si en sus soluciones aplicaban adecuadamente la teoría de la evolución por selección natural. En ambos problemas las respuestas correctas superaron el 80%.
Estos resultados parecen ser esperanzadores respecto a la eficiencia del material didáctico utilizado, si bien deberemos profundizar en las causas que provocan que un 20% de alumnos hayan dado todavía explicaciones lamarckianas. Será preciso, pensamos, dar respuesta a cuestiones como:
-¿De qué manera se puede afrontar en el aula y en el centro las dificultades que tienen algunos alumnos y alumnas para realizar un cambio conceptual y metodológico?
-¿Hasta qué punto instrumentos básicos de aprendizaje como: la lectura comprensiva de textos, la realización de resúmenes, la capacidad para realizar debates, de exponer con claridad y concreción las ideas... puede facilitar este cambio?
-¿Se puede facilitar desde la enseñanza primaria el cambio metodológico?
-¿Cómo relacionar la enseñanza de las ciencias en la escuela primaria con la secundaria para favorecer este cambio?
-Etcétera.
Nota: Este trabajo ha sido realizado con la ayuda del institut d'Estudis Ilerdencs de Lleida.
ANEXO
En este anexo presentamos a modo de ejemplo algunas actividades del programa-guía diseñado para trabajar el tema de la evolución de los seres vivos por selección natural.
Hemos escogido tres apartados de la unidad:
-El apartado 2 ¿Puede variar el número de individuos de una población?, tiene como objetivo ayudar a los alumnos de definir y comprender el problema que nos ocupa.
-El apartado 3 ¿Qué pasa cuando las condiciones ambientales cambian? Forma parte del bloque destinado a la construcción de nuevos conocimientos.
-El apartado 10 ¿Puede explicarse la evolución humana por la teoría de la evolución por selección natural? ¿Cómo se originaron las

148
mujeres y los hombres? Reúne actividades de aplicación de los conocimientos adquiridos y de reflexión sobre el papel de la cultura en la evolución humana.
Las actividades se presentan acompañadas de unos comentarios para facilitar la aplicación en el aula.
2. ¿PUEDE VARIAR EL NÚMERO DE INDIVIDUOS DE UNA POBLACIÓN?
Actividad 1.
Los elefantes se reproducen muy lentamente, a pesar de ello en 700 años una pareja de elefantes podría haber tenido 19.000.000 de descendientes.
Una elefanta puede tener la primera cría a los doce años, y tener otra cada dos años. Si las crías sobreviven todas y tienen cachorros y se reproducen cada dos años... podríamos explicar fácilmente estos resultados.
Compara las dos imágenes que muestran la misma zona de una sabana africana natural inalterada por donde no han pasado nunca los humanos. La primera imagen es de hace 100 años. La segunda es de nuestros días.
Emitir y fundamentar hipótesis que permitan explicar esta situación.
(Nota: Se muestran aquí dos dibujos en los que aparece la misma zona de la sabana con prácticamente el mismo número de elefantes a pesar de que hay entre las dos escenas una diferencia de 100 años.)
Comentario: Es necesario insistir en que los seres humanos no han intervenido en ningún momento en la vida de esta sabana, ya que así se fuerza a los alumnos a dar explicaciones naturales al problemas que se plantea. En general, se refieren en sus hipótesis a las condiciones adversas: falta de alimento, escasez de agua, presencia de muchos depredadores, epidemias...
Indican también que, cuando esto ocurre, los individuos que primero mueren sin los más débiles.
El problema que sugieren sus hipótesis es por que un determinado individuo es más débil que otro de su misma especie y, por tanto, tiene mayores probabilidades de ser eliminado por los factores naturales. También preguntan que es lo que determina que un individuo sea más fuerte que otro y, en consecuencia, con mayores posibilidades de resistir a las condiciones adversas.
Antes de abordar este problema, clave para la comprensión de la teoría de la evolución por selección natural, pensamos que es necesario que los alumnos comprendan que esto ocurre en todas las especies y que además el factor humano puede jugar también su papel. Para ello proponemos la realización de la siguiente actividad:
Actividad 2:
Pensad ejemplos concretos de especies de animales y plantas en las que:
1) el número de individuos vaya disminuyendo
2) el número de individuos aumente
3) el número de individuos sea prácticamente constante.
Indicar las causas que, en nuestra opinión, están contribuyendo a su disminución, aumento o equilibrio.
Contrastar vuestras opiniones con la bibliografía.
Comentario: esta actividad favorece el trabajo autónomo de los pequeños grupos. Es preciso, sin embargo, discutir la estructura del trabajo y exigir concreción en las conclusiones.
Una manera de hacerlo es organizar una mesa redonda en la que cada grupo pueda hablar durante un tiempo limitado.
Después de las exposiciones se puede abrir el debate para proponer mejoras, opiniones contraria, críticas...
Después de esta actividad, vamos a recuperar el problema que se suele plantear en la actividad 1, o bien lo presentamos nosotros.
Actividad 3
¿Qué determina que un individuo sea más débil o más fuerte y que, en consecuencia, sea eliminado por la naturaleza o pueda sobrevivir?
Emitir hipótesis que permitan dar respuesta a esta cuestión.
Comentario: Aquí surgen interrogantes sobre el papel de la herencia por si misma. También sobre la relación entre el ambiente y la herencia: ¿Puede el ambiente actuar favoreciendo una determinada característica de un ser vivo? ¿Quién actúa primero, la herencia o el ambiente?...

149
Las hipótesis generales que se plantean en los grupos y a lo largo de la puesta en común para dar respuesta a estas cuestiones nos empiezan a aproximar a la tesis defendida por Darwin. Queda todavía un largo camino.
3 ¿QUÉ PASA CUANDO LAS CONDCIONES AMBIENTALES CAMBIAN?
Actividad 1.
Las mariposas geometras son muy abundantes en la Gran Bretaña. Bastantes especies de pájaros se alimentan de estas mariposas, que cazan cuando se encuentran descansando en las cortezas de los árboles durante el día.
Antes de 1850 casi todas las mariposas geómetras de la ciudad de Manchester eran de un color claro que se confundían muy bien con las cortezas de los árboles en los que descansaban.
Una colección de mariposas geómetras recogida en aquella época, tenia el siguiente aspecto.
Nota: se muestra aquí un dibujo de una colección de mariposa con una de ellas de color oscuro. Todas las demás son grises.
A lo largo del siglo XIX, el ambiente de la ciudad de Manchester cambio radicalmente. Antes de la revolución industrial las cortezas de los árboles donde descansaban las mariposas geómetras eran de color claro porque tenían unos pequeños líquenes ilustrados en ellas. La contaminación producida por las fabricas a finales del siglo, mato todos estos líquenes y oscureció las cortezas.
La colección de mariposas geómetras recogida a principios del siglo XX tenía este aspecto.
¿Cómo explicarías estos cambios en la población de mariposas geómetras?
Comentario: Aquí, la mayoría de las hipótesis se refieren simplemente a que las mariposas negras aumentan porque pasan desapercibidas y claras disminuyen al ser comidas por los pájaros.
Algunos alumnos se refieren también a que algunas mariposas se vuelven negras al adaptarse al nuevo medio, pero abandonan a sus concepciones al realizar la actividad 2, no solamente es el peso de la propia cuestión, sino también por el peso de los argumentos en contra defendidos por sus compañeros y compañeras de clase.
Actividad 2.
En 1956, el gobierno ingles promulga una ley para reducir la contaminación atmosférica. Desde entonces, la proporción de las mariposas geómetras de color claro recogidas en áreas industriales ha vuelto a crecer.
¿Como se ha producido este aumento de las mariposas claras?
Actividad 3.
Contrastad vuestras hipótesis con el siguiente documento:
Las investigaciones realizadas han permitidos confirmar que los pájaros encuentran y comen más mariposas oscuras que las sobre las cortezas de los árboles cubiertas de líquenes, y las mariposas claras sobre cortezas oscuras.
En una zona industrial, las mariposas oscuras tienen mayores probabilidades de sobrevivir y dejar descendencia. Al cabo de cierto tiempo aumentará, por lo tanto, la proporción de las mariposas oscuras de una población.
Con ayuda a explicar el cambio rápido sufrido por la población de las mariposas geómetras en las ciudades industriales en los primeros años de la revolución industrial.
¿Qué está pasando ahora?
En el año 1956, después de la ley para reducir la contaminación atmosférica, la cantidad de mariposas claras de las zonas industriales ha vuelto a crecer. Las mariposas oscuras que ahora descansan en las cortezas claras de los árboles cubiertas de líquenes son mejor vistas por los pájaros. Así, se ha ido reduciendo su número y aumentando el de mariposas claras que pasan más desapercibidas.
Adaptación de un texto de British Museum (1981)/Diputació de Barcelona (1982). - L’origen de les espécies.
Comentario; este bloque de actividades resulta generalmente útil e interesante, ya que la teoría propuesta por Darwin permite explicar muy bien los problemas planteados.
Es sabido que para “derrumbar” una teoría científica. Lamarckista en el caso de los alumnos- no es suficiente demostrar que ésta no sirve para dar respuesta a determinados problemas, es preciso ver que existen otras teorías que sí explican estos problemas.
10. ¿PUEDE EXPLICARSE LA EVOLUCION HUMANA POR LA TEORIA DE LA EVOLUCION POR SELECCIÓN NATURAL? ¿CÓMO SE ORIGINARON LAS MUJERES Y LOS HOMBRES?

150
Comentario: no se puede terminar el tema sin abordar la problemática de la evolución humana. Por este motivo proponemos unas breves actividades que tienen como objetivo hacer ver a los alumnos que es posible aplicar la teoría de la evolución por selección natural a la evolución humana.
Para contrastar la hipótesis de los alumnos, proponemos la lectura de unas paginas del libro de J. Monod, El azar y la necesidad. Editorial Tusquets, pp. 163-167.
Actividad 1
En la figura 1 se muestra la imagen imaginaria y el cráneo de un animal llamado procónsul que vivió sobre la Tierra hace unos 25.000.000 de años. Su cráneo tenia unas características parecidas a las del hombre y la mujer actuales y también al de los monos.
En la figura 2 se puede ver la imagen de un Australopitecus que vivió hace 5.000.000 de años y que considera como el miembro más antiguo descubierto de la familia humana.
¿Cómo se podía explicar de acuerdo con la teoría de la selección natural de Darwin el origen de los seres humanos.
Emitir hipótesis de trabajo.
Actividad 2.
Contrastad vuestras hipótesis con la lectura del texto de J. Monod, El azar y la Necesidad.