bioi. maria s anoeles aüüilae s.148.206.53.84/tesiuami/uam lote 5/uam20374.pdf · de la rana en...
TRANSCRIPT

Bioi. MARIA S ANOELES AüüILAE S.

r.
INFORME FINAL DEL SERVICIO SOCIAL
"DEPEIWINACION DEL " E R O CROYIOSOMICO, ELABORACIGN DEL
CARIOTIPO E IDIGGRAMA Y BANDAS CROMOSOMCAS DE LA
POBLACION DE Rana berlandieri DEL ESTADO DE TLAXCALA".
CRUZ OARCTA ALIC IA (No. matr. 79221895)
MARTINEZ OSORIC FRANCISCA SGCOaRO (No. iaatr. 7%?21833 1

--+==== c.
Agradecemos la valioea aguda de
B i o l . Maria de los Angeles Aguilar Santamarfa
Biol. Alma Guadrilupe Arellano Meneees

I NTRGDUC C ION
Rana be r l and i e r i e S comunmente conocida como rena - leopardo. oscuras ovaladas o alarga.das con bordes de co lorec ión inhv tenue (dependiendo de l a subespecie que se t r a t e ) . E l c o l o r d e l a rana en l a reg i6n vent ra l es blango nacarado y en l a dorsa l es generalmente gr isáceo con t om l i dades verdoses y negras; se caracterizó.n estos orgtnismos pcr presentar pl iegues dorso la te ra l es generalmente descoloridos muy bien de f in i&o i , que pueden estar desplazados a l a hltura d e l u ros t i l o o bien s e r continuos, también presenta una l i n e a tenue en ia mandíbula super ior menos prominente que en l a s o t ras rmas leoprrdo, usuelmente es incomplettr en l a parte Ernterior de l o s o jos . Los machos presentm unos sticos bucales externos muy marcados &sf coino oviductos v e s t i g i a l e s (Rudich, 1980).
Es una rana. robusta con un patrón dorsa l de manchas
Taxonornia:
Reino Animalia Subreino Chordata Subphylum Vertebrata Sup e r c l a s e Tetrapod a Clase Amphibia Orden finura Suborden D ip las i coe la Famil ia Rmidae Género Zspecie - Rsna ber land ie r i , (Rudich .) 1980).
E l nombre p ip i ens agrupa a un complejo de rnzss o especies empnrentsdas cuyo t e r r i t o r i o de distr ibuc ión s e extiende desde e l sur de Canad6 hasta México y Costa Rica (Bejar, 1981).
Los t rabhjos y evidencias acumuladas hast& l a fecha. indican que e l complejo 'R. p ip iens está formado por une s e r i e de espec ies que difieren-unm de o t r r v en sus propiedades ndapteitivas y ca rac t e r í s t i cas v i s i b l e s , pero que varían gra- dualmente en mb i en t es intermedios. Untre iós trabbu,jos encá- minados s. c l a s i f i c a r a l a s r a m s leoptrrdq se encilentrsn l o s reb l i zados por L i t t l e j o h n y (ildharn (1968) en I& regidn centrúl
.

. .
.. . de l o s Estados Unidos; ellos estudiaron el c m t o de l a s rams ya que éste es específico parr cada especie y al obtener el patrón de canto de apareamiento observaron una variación geo- gráfica en la estructura del llamado. cuatro f o r m a s álopátricas (poblaciones o especies que viven en ereas diferentes) que llamaron: norte, sur, este y oeste. Su distribución se puede observar en la figura 1 y l ~ s ca- racterísticas de los llanados de apareamiento se dan en l a tabla 1.
Distinguieron así
t -N-
Fig. 1. Distribución geográfica de l a s ratis lempardo en función del tipo de canto, en la regiCn central de l o s EsQa- dos Unidos. (norte O, oeste , este , sura). (Littlejohn y O l d h a m , 1968).

T ipo de No. de Duración d e l Tasa de Duración d e l llamado individuos llarnado pulsos pulso
í seg) (No/seg) (mm/ses)
N 4 3.75
O 7 0.66
h 7 0.41
S 7 0.64
( 3 30 -4.73 1
(0.48 -0.89)
(0.31 -0.52)
(0.47 -0.83)
13 -9 (12.9-14.6)
5.6 ( 4.6- 6.8)
14.8 (14-3-15 3)
28.2 (26.0-31.3)
12.8 (16- 20) 27.0 (23- 35) 35.4 (33- 50) 19.1 (16- 22)
Tabla 1. Valores promedio de algunas ca rac t e r í s t i cas en e i canto de aparea- miento de - R.pipiens, entre paréntesis se muestran l o s rangos de va l o r es obtenidos ( L i t t l e j o h n y Oldham, 1986).
E l conjunto de rasgos d i s t i n t i v o s como son tempera- tura de adaptación, velocidad de desarro l l o , tempertrtura m b i e n t a l , f recuenc ia en e l llamado de aparearniento y caracterev mo r f o i 6 g~cos como p l iegues dorso la te ra l es , pigmentación, número y d i s t r ibuc ión de Itis manchas dorsales y presencia o ausencia de oviductos vestigi6.les en nachos trduitos han permitido d i s t i n gu i r l a s d i f e r en t es formas de ranas leophrdo part su c l as i f i cac ibn . separaron a l a s R. pipiens de i o s Estados Unidos de l a s ranas mexicanas considerkndolas a éstas como una especie d i s t in ta : - R. ber l end i e r i ( t a b l a 2).
De e s t a manera Sanders y Smith (1971)

P l i e gues Gviductoe L ine& de i i ginent o Especies do rso l a t e ra l e s en machos los l ab ios 6n iu i l l o
- - R. p ip iens continuos presentes completa ausente
it. b e r l snd i e r i despiazhdos present es incompleta present e - -
Tabla 2. Resumen de l o s pr inc ipa les caracteres morfoldgicos tomados en consideración para 18 c i a s i f i c a c i ó n de espec ies (Sanders y Smith, 1971).
Posteriormente Sanders (1973), a l estudiar l a d i s t r i - bución de l a s manchas dorso latera les en 1a.s rsncs leopardo mexicanéiq es tab lec id un pztr6n de ca rac t e r í s t i cas morfcldgicas ( t ab l a 3 ) e l cual co inc id í a ccn un rengo de d i s t r ibuc ión geo- g rá f i co , de t a i manere que i a especie g. )~cixn$&_oi queab forniti.da por t r e e subespecies que ccn: E. b. b-eritindiea, le- cti l izada a l es te d e l t e r r i t o r i o mexicano; g. &. f o r r e r i , l a cual hs.bita l a regibn oeste y p o r Últiuio 16 subespecie (. b. brownorum 16 cual se extiende a l sureste de l t i HepÚblics, ~ e x i c a n a r (fig* 2) .
S ubespecie Dibujo dorsa l P l i egues dorso lhteró ies
R.b.berlandieri manchas la rgas discontinuos poster0 - - - i r r egu la r es con medialmente fusión frecuente
R .b.f o r r e r i manchas la rgas continuos y ovtiladss con f recuenc ia
- - -- H. b. brownorum mancht>s i i neú l es continuos C b r t G S - -
Tabla 3. Caracter ís t icas mcr fo l i g i cas de Itis subespecies ñ.beriandieri (Shnders, 1973) .
I

t 1
w-
Pig. 2. Distribucidn geográfioa de g.berlandieri (area punteada). (Bejar, 1981).

. ,.
..
...
.., ,
....
"... -<
C"
I.
F'.
.._. C.
.*< -
...<
c.
L.
,p-
.... r.
1..
r.
L. - -.
m.
** .
"I.
LI
Otros métodos usados para ac io rar l a pos ic ión taxonómica de l a s ranas leopardo es a trzvés de cruzas interpoblac ionales con e l c u l t i v o pos t e r i o r de l o s renacuz j o s y l a observación de su desarro l lo . Un ejemplo es e l caso de Fros t ( 1 9 8 2 ) que hace un^. evduac idn d e l grado de compa- t i b i l i d a d genktica por h ibr idac ión experimental entre pobla- ciones de - - R.b.berlandieri, 2 .b . f o r re r i y g.magnaoculeris.
d i método usado para evaluar l e s cruzas interpo- b lac iona les c cns i s t i ó en tonm l a r e l ac i ón B/A ctimo e l porcen- t a j e de compatibii idad genét isa entre l a s rcna,s leop,truo consideradas en dicho estudio ( A es e i n b e r c de huevbs fe - cundhdos, B es e l número de huevos fcrmedos) teniendo coino cont ro l l c ~ cruza entre machos y hembras &.b.beriandieri a Ih cuai s e i e asignó un iüO$ de compkitibilidad.
Los resultados mostrkron n i v e l e s contrt s t m t e s de compatibil idad, por ejemplo al cruzar machos R.b.forreri con hembras - - R.b. b e r l k n d ~ . g ~ ~ aproximadamente e l 9546 de l o s embrio- nea fueron organismos v iab les , en contrhste l o s embriones de l a s cruzas rec íprocss (machos R.b.berlandieri --- con hembras - - - R.b. forrer i ) fueron extremadamente anormales con respecto a l o s contro les ( t ab l a 4).
En lzs cruzas entre R.magnaocularis con - R.b.berlan- - --I_
d i e r i l o s embriones h ibr idLs de cada unir de l a s cruzas en 12s dos direcciones presentaron un patrón s im i l a r de dessmroiio con respecto a l o s controles, mientras que l a s cruzas de machos R.magnaocular9.s I con hembras &.&.forreri produjeron embriones anormales (con microce fa l i a y c o l a f l e x i b l e pr inc i - palmente).

. .
NO. de Hibridos va. Macho s Hembros cruzaa controles ($\
Rebebe R.b.b. 3 100 Rebef. R.b.f 6 100 R.b.f Rebebe 4 95 l2.b.b. Robef. 4 O R.b.f R.b.f. X Rebebe 2 43 R.m. Rebebe 1 80 R.b.b. R.m. 1 94 R.m. Robof. 5 O -
Tabla 4. Compatibilidad gendtica observada en las crueas entre diferentes especies y subespecies de ranas leopardo. R.b.b.r iJ.b.berlandie$i; R.b.f.r g.b.forreri; R.m.r R. magn aocularis (Frost, 1982).
Otros datos sobre l a compatibilidad genética 8.
-
obtuvieron comparando las proteínas d e l suero (albúmina) entre l o a individuos silvestres g.&.berlandieri, &b.forreri y un hibrido de estas dos aubespecies; ai análisis eiectro- fo rdt ico revel6 que cada raza presentaba un a l e l o para l a albúmina pero e l híbrido presentaba ambos a le los (Frost, 1982). En consecuencia l e posición de &b.forreri como una sub- especie de R. berlandieri (sanders y Smith, 1971; Sanders, 19731, es iñcorrecta, debiendo considerarse a g.b. forreri como una especie distinta --_I R.forrer i (Proat,i982).
La variacidn de los cromosomas de una especie de anuros respecto a otra permiten en l a actualidad dibujar un &bol geneoiógico que comprende desde los antepasados más lejano8 hasta los más próximos.

. .
.. .. Comparando los cromosomas de las d i s t i n t a s espec ies s e puede conocer para cada cromosoma una v a r i a c i ó n a i pasar de una espec ie a o t r a , v a r i a c i ó n representada p o r modificaciones en l a morfo1oe;la de cada cromosoma, l a c u a l e s parec ida para t o d o s los individuos de una e s p e c i e (Morescalchi , 1973).
L a figura 3 muestra un &bol geneológico basado en c a r a c t e r e s anatómicos donde s e puede a p r e c i a r e l n i v e l que ocupa l a familia Ranidae. En l a figura 4 aparece un esquema que muestra las p o s i b l e s r e l a c i o n e s c a r i o l ó g i c a s de las p r i n c i p a l e s familias de enuros, en 61 s e puede observar l a morfologla de los c a r i o t i p o s .
E l hecho de que l o s c a r i o t i p o s de l o s a n f i b i o s su- p e r i o r e s no presenten cromosomas a c r o c é n t r i c o s i m p l i c a una c i e r t a f u s i ó n de sus cromosomas, f u s i ó n d e s c r i t a p o r Mores- ca lch i (1973) y Goin ( 1 9 7 8 ) , donde s e d n e l l o s l a evoiución de los cromosomas a n f i b i o s s e ha l l evado a cabo p o r f u s i ó n de los microcromosomas y cromosomas a c r o c é n t r i c o s ya que las formas p r i m i t i v a s t i e n e n un número elevado de e l l o s .
P o r es tudios c i t o g e n é t i c o s s e sabe que las e s p e c i e s s u p e r i o r e s de a n f i b i o s t i e n e n un número cromosómico menor a l que presentan las formas i n f e r i o r e s , e s t o i n d i c a que hay una tendenc ia a is reducción d e l c a r i o t i p o que puede l l e v a r s e a cabo p o r d i s t i n t o s mecanismos, e s t a s modificaciones pueden s e r observadas con p r e c i s i ó n gracias a las t é c n i c a s de bandeo. Dado que e l patrón de bandas e s c a r a c t e r f s t i c o p a r a cade porc ión de cromatina, cada segmento puede s e r reconocido i n c l u s o s i no e s t 6 en e l l u g a r que l e corresponde en un de- terminado cromosoma.
Las t é c n i c a s de bandeo cromosómico son ampliamente u t i l i z a d a s para e s t u d i a r los cromosomas de v a r i o s organismos, en p a r t i c u l a r en e l estudio de homologfa cromosómica, lo-- c a l i z a c i b n d e l DNA s a t é l i t e y de las reg iones d e l organizador n u c l e o l a r , Et otras pa labras e s t o s es tudios permiten r e l a - c i o n a r el. n i v e l molecular de l o s cambios evolut ivos con e l n i v e l cromosómico ( B i r s t e i n , 1982) .
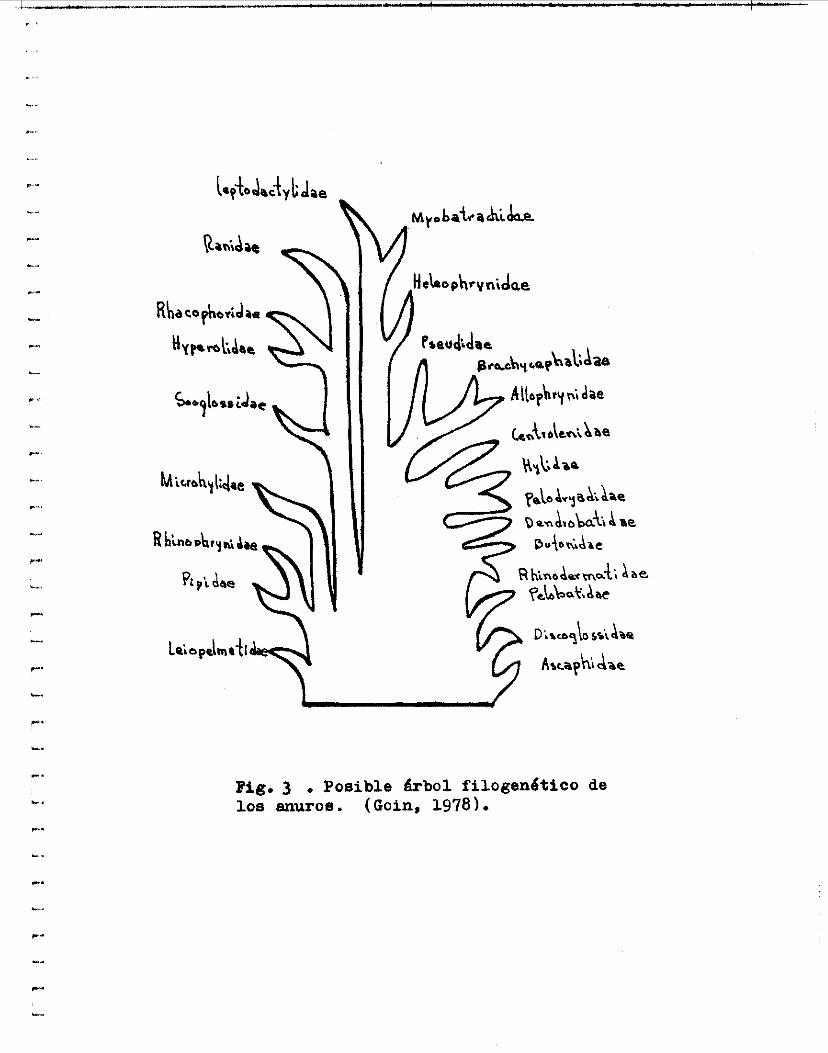
Pig. 3 . Posible &bol fi logenético de loa muroe. (Goin, 1978).


til ob j e t i v o general de este t rbbajo es cont r ibu i r a l conocimiento de l a s re l ec iones f i l o g ené t i c a s en l a pobla- c ión de - H. ber lánd ie r i que habita en e l Estado de Y laxca la a través de l a mor fo log ía cromosómica, y l o s ob je t i vbs especí- f i c o s son l o s siguientes: e laborar e l ca r i o t i po , elaborar e l idiograma y estandcrizer l a t é cn i ca para obtener bmdas G.
Determinar e l n h e r o cromocómico,
MATERIAL Y M!X”l’DO
E l método de es te t r cba j o es t6 d i v id ido en s e i s
a ) Co lecta de mater ia l b i o l ó g i c o b ) Obtención de cromosomns c ) Determinación d e l n h e r o cromosónico d ) GbtenciÓn d e l ca r i o t i po e ) Zlaboracibn d e l i d i o g r a m f) Obtención de bandas cromosdmicas
partes:
a ) Obtención de mater ia l b i o l óg i co :
Se co lectaron 19 rznas en Tecomalucan Edo. de Tlax- ca l a ; 12 de e l l a s fueron usadas en experimentación y l a s demás se encuentran f i j a d a s (en alcohol a l T O A ) con e l f i n de t ene r l as como re ferenc ia . sus cnxacter ís t icas inorfoló- g i c e s aparecen en e l anexo.
b) Obtención de cromosomas:
Las ranas fueron inyectadas intraperitonealmente con una solución de co l ch i c ine (Sigma) a l 0.316 pma s e r sacri- f i c adas después de 10, 6, 4 y 3 horas con e l f i n de obtener cromosomas con d i s t i n t o s grudos de coinpactación. Se aneste- s ia ron con é t e r e t í l i c o y se colocaron en u n a chero le de d i s ecc i ón donde se l e s l imp16 y cor tó l a reg ión vent ra l a i c i a i t w s d e l esternón, removiendo e l per i card io para d e j a r expuesto e l corazón. Con una j e r inga prev imente heparinizada se h i z o una punción cardihcz y se i x t ru j o l a sangre permi- t i endo que cada l b t i d o contribuyera a su llenado.

La sangre así obtenida se sembró en fraacos dmpula (2 a 3 gotas por frasco), conteniendo 5 m l de medio Yc Coy (Microlab) d i luido 3:2 con agua bidesti lada e s t é r i l , 0.02 ml de una solucidn ant ibidt ica "Antibac" (Microlab) y 0.2 ml de fitohemaglutinina (Microlab). incubaron a temperatura ambiente entre 96 y 168 horas o hasta l a aoidi f icaci6n de l medio. Transcurrido este tiempo se aña- dieron a l o s cu l t i vos 2 a 3 gotas de coichicina (Sigma) a i 0.396 de 10 a 4 horas antes de l a cosecha.
Los frascos con l a sangre se
Inmediatamente después se procedió a extraer l a mé- dula ósea haciendo las disecciones de fémur, hhero y t i b i o f í - bula. con medio Mc Coy (Microlab) d i luido 3r2 con agua bidesti lada e s t é r i l . La médula 6sea con e l medio se c o h c t ó en tubos de centri fuga para continuar con l a cosecha.
Se cortaron l a s e p í f i s i s y se extrajo l a médula ósea
Para extraer e i e p i t e l i o intestina& éste se cortó, maceró y se dejó incubando con t r ips ina a 37 C durante 20 minutos, luego se hic ieron pasar las células por una malla y se colectaron en tubos de centrffuga con medio Mc Coy (Micro- lab) y agua bidest i lada e s t é r i l (3:2).
Una vez que se tuvieron las células de médula ósea, sangre y e p i t e l i o in tes t ina l en tubos de centrífuga se proce- d i6 a l a obtencidn de cromosomas se& la técnica descr i ta por Bogart (1970).
El material obtenido se resuspendió vigorosamente y se centrifugd a 1500 rprn durante 10 minutos, se desechó el sobrenadante dejando 0.5 ml sobre el paquete ce lular y se resuspendi6; con agitación constante se añadieron 5 m i de soiuci6n hipot6nica (KC1, 0.046 bl) durante 20 minutos, se centrifugd a 1500 r p m durante 10 minutos desechando e l sobre- nadante y dejando 0.5 m l sobre el paquete celular; de igual manera se agregó solución f i j ado ra (metanoi-ácido acét ico 3:l ) y se centrifugó a 1500 r p m durante 10 minutos; los lavados con metano1 y ácido acético se repit ieron hasta que e l sobre- nadaate apareció claro.
c ) Determinacidn d e l número cromosómico:
Con e l material obtenido se hicieron preparaciones

p o r e i método de secado a l a fiama. Las preparaciones se rev i saron a l microscopio de contraste de fases , conthndo y registrando e l número de croinosomas en cada mitos is (50 por ind iv iduo) para poder determinar e l n h e r o modal.
d ) Elaboración d e l car io t ipo :
Las preparaciones se t iñeron con Giemsa (Sigma) a i 10% durante 20 minutos; se seleccionaron l a s mejores m i t os i s de cada individuo, con número cromosÓmico completo (número modal). A dichas mi tos i s se l e s tomaron microfo- t o g r a f í e s y se recortaron l o s cromosomas ordenhdoios p o r pares, de mayor a menor tamaño.
e ) Elaboración d e l idiograma:
A l o s ca r i o t i pos obtenidos se l e s tomeron l a s medidas promedi.0 y se cálcularon l a s medidas r e l z t i v a s de l o s brazos l a r g o s y cortos a s í como l a longitud t o t d de cada uno de l o s cromosomas; para elaborar e l idiogrsma.
Para designar a los cromosoinzs coino metccéntricos (m,, submetacéntricos (sm) o acrocéntricos ( t ) se u t i l i z ó l a nomenclatura. propuesta p o r L e v a (19641, donde l a re la - c ión u p da l a pos ic ión d e l centrómero; q es e l tamaño r e l ú t i v o d e l brazo lárgo y p es e l tamaño r e l a t i v o d e l brezo corto.
Para obtener l a s medidec r e l h t i v a s se calculeron l o s promedios de l o s brazos cor tos (Zp) y l a r g o s ( X Q ) de crda cromosoma y l a sumsitoria de todos e l l o s diÓ un inhero a l cual se l e asignó e l l0O.b de l a long i tud t o t a l , luego a crda cromosoma l e correspondió un porcentaje de esa longitud.
f) Obtención de bandas cromosómicas:
Las prepareciones s i n t e ñ i r se dejaron incubando O a 37 C durante 10 ininutos en una solución de t r i p s i na (D i f co )
a l 0.005% en NaCl (J.T.Baker) a l 0.9% (aumentsndo e l tieinpo a in t e r va l os de 3 minutos según se observaran l o s cromosomas). Inmediatamente después l a s preparaciones se lavzron en so- luc i ón sa l ina (NaC1 a l 0.9s) dos veces y s e t iñeron con Giemsa (Sigma) a1 56 en b u f f e r de Sorensen.

RESULTADOS
Se colectaron 19 ranas de Tecomalucen, Estado de Tlaxcala; l a mayoría eran individuos que pesaban entre 6 y 15 gramos y que acababan de pasar por su estado de metamorfo- sis por l o que solo se pudo determinar e l sexo a dos de e l las.
A se i s de l a s ranas colectadas (UAMI-10 a UAMI-15) se l e s inyectd colchicina y se sacr i f icaron después de 6 y 10 horas para obtener un mayor número de c4lulas en metafase; las céiuias se obtuvieron a pa r t i r de sangre, e p i t e l i o intes- t i n a l y médula ósea. Se r eg i s t r ó un t o t a l de 311mitosis de las cuales 165 presentaron 26 cromosomas mientras que l a s otras 146 mitosis presentaron entre 20 y 25, por lo que e l n h e r o cromos6mico quedd determinado como 2-26 ( tab la 5).
Se tomaron 1 2 fotograf€as de l a s mejores mitosis y se elaboraron l o s car iot ipos los cuales quedaron formados por 5 pares de cromosomas grandes, 8 parea de cromosomas pe- queños, ausencia de cromosomas sexuales; también se observa una conatriccidn secundaria en los brazos largos del par ni€- mero d i e z ( f o t og ra f í a 1).
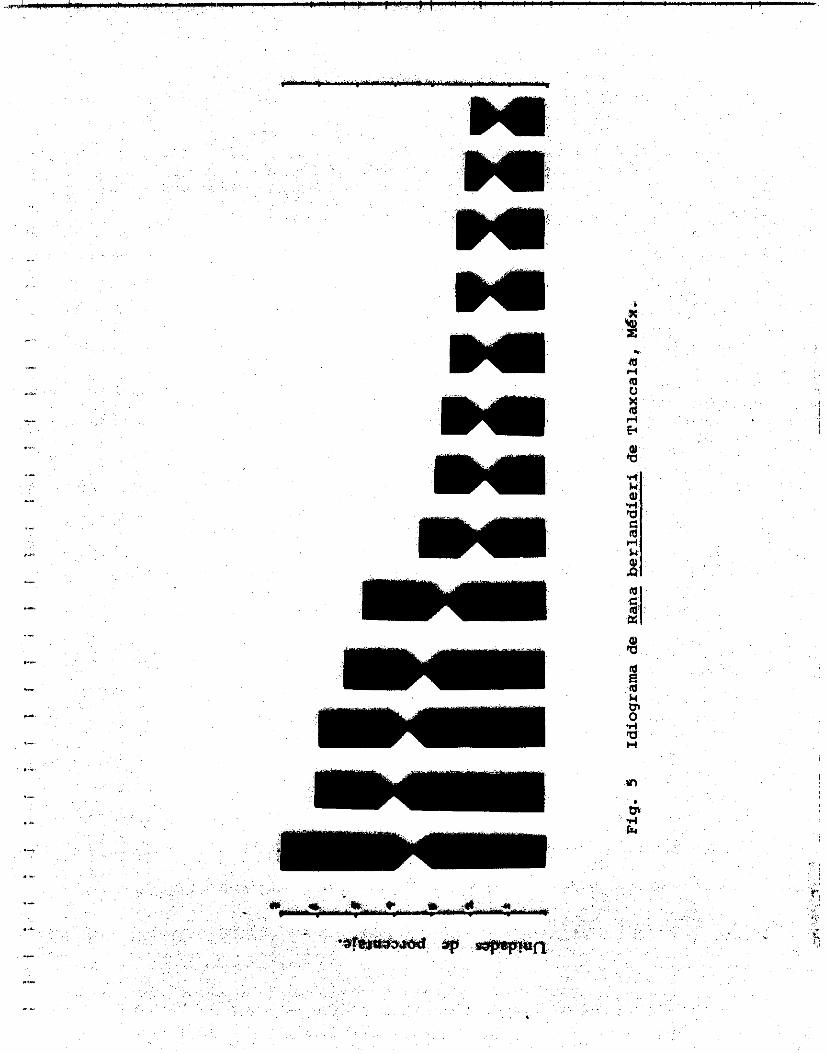
Las tablas 6 y 7 muestran l a s medidas obtenidas para construir e l idiograma ( f i gura 5 ) y calcular las longitudes r e l a t i vas promedio de los 1 3 pares de cromosomas.
Para obtener bandas cromos6micas se redujo e l tiempo de colchicina de 6 a 3 y 4 horas, de esta manera disminuyd e l número de mitosis pero su calidad fue mejor. El bandeo cromosómico se l o p 6 usando une concentracidn de t r ips ina a l 0.025% en solucidn sal ina (NaC1 0.9s) durante 10 minutos.
La f o t og ra f í a número 2 muestra una mitcsis con bendas 0 y el arreglo de los cromosombs hom6logos de acuerdo con l a correspondencia de l a s bandas.
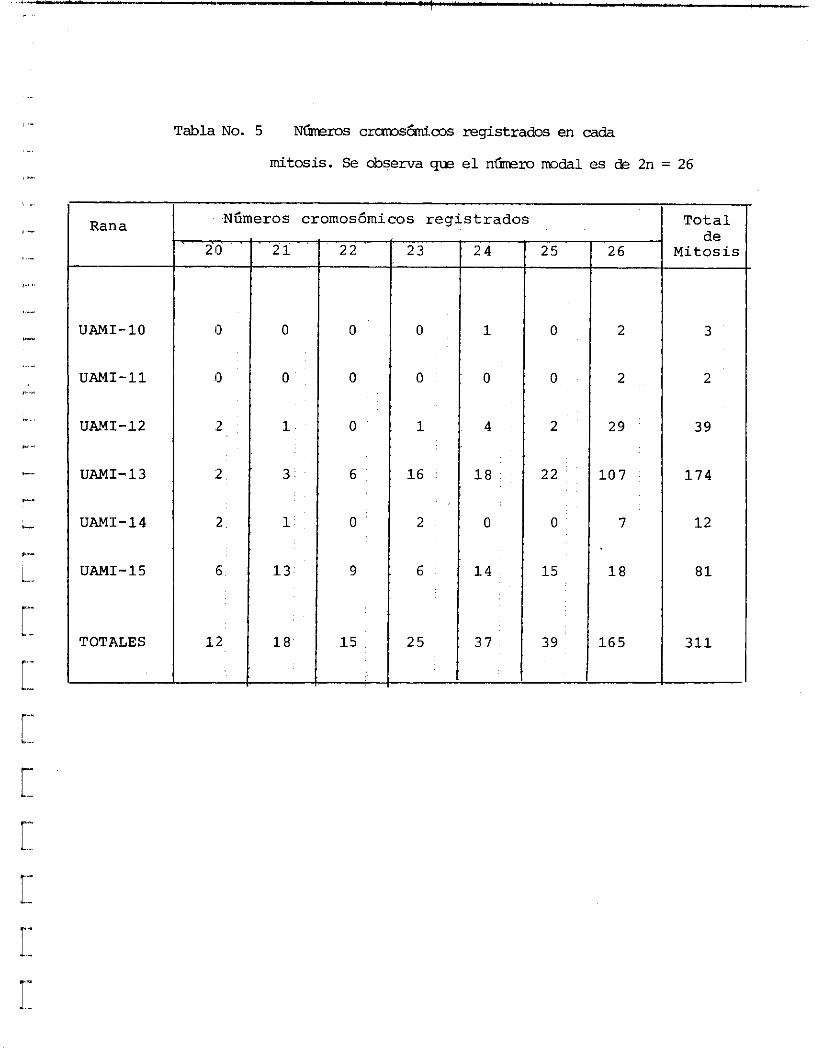
Tabla No. 5 N&ms c m s ~ m s reg i s t radoc en cada
mitosis. Se observa qw e l n h m modal es de 2n = 26
I Números cromosómicos r e g i s t r a d o s Ran a
UAMI-10
UAMI-11
UAMI - 1 2
UAMI-13
UAMI-14
UAMI-15
TOTALES
IO
o
2
2
2
6
1 2
2 1
O
O
1
3
1
13
18
22
1 5
23
O
O
1
1 6
2
6
25
2 4
1
O
4
18
O
1 4
37
25
O
O
2
22
O
15
39
26
2
2
29
1 0 7
7
18
1 6 5
T o t a l de
M i tos i s
3
2
39
1 7 4
1 2
81
3 1 1

.-
c
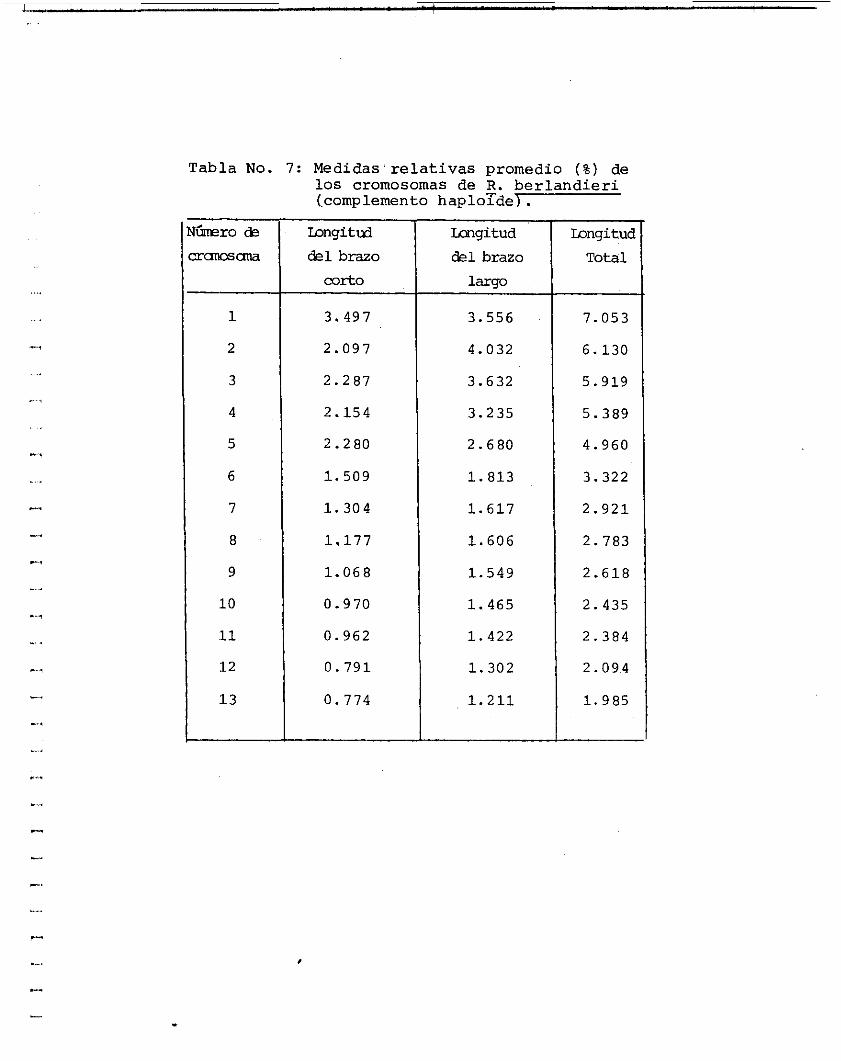
Tabla No. 7: Medidas'relativas uromedio í%) de
h r o de XalKXiaria
1
2
3
4
5
6
7
8
9
10
11
:L 2
1 3
~ .~ ..
los cromosomas de 8. berlandieri (complemento haploTde) .
Iongitui del brazo
Corto
3.497
2.097
2.287
2.154
2.280
1.509
1.304
1.177
1.068
0.970
0.962
0 .791
o. 774
limgitud del brazo
largo
3.556
4.032
3.632
3.235
2.680
1.813
1.617
1.606
1.549
1.465
1.422
1.302
1 . 2 1 1
bngitud Total
7.053
6.130
5.919
5.389
4.960
3.322
2.921
2.783
2.618
2.435
2.384
2.094
1.985

a, a
m m a ‘4 a 3
W
m m P o m m - ~ ~ m r n w P ~ m r n ~ ~ d w m ~ r n m o m o m w m N rl (u m m m tn P P
~ m m m o d m m % Z i Z Z ! ! 2 m w m - c o p - m P P r’ w m, m m m m N m N N 0 0 0 0 0 0 0 0 0 0 0 0 0
m m m w q m N m m m m m d m m m m m m o m o - P W W m q m - m d N m m r l d d r l 0 0 0 0 0 0 0 0 0 0 0 0 0
r - m m ~ - m m w ~ m w w N w w 4 w Z m m o m m m m m ~ N N 4 4 N 4 d r l d 4
0 0 0 0 0 0 0 0 0 0 0 0 0
4
991 x
"A
I-
*"
- I_-.
...
w..
- c O
rl v U
.- *-
...
vi
*-
I-

r-
. .
C'
F..
.. -
.^I P
L
F
... . c .
.*. P
..*.
C"
*,,_
P
le at
U

,
, , ..
I _.
I -
..,-
I-
&.*
..- ..*
I--
-..
c.
Y
c
L..
*-
..... r-.
..__ c< . -.<
c
, ...
e-
-- .* .- r- L.-
DI SCUSI CN
Muchas de l a s e s p e c i e s d e l género - Rana: R.pipiens ( D i Berardino, 1 9 6 2 ) , R. e s c u l e n t a , c a l c h i , 1 9 6 7 ) , e t c . , presentan un n&%o dip lo ide de 26 cromosomas foruados p o r 5 p a r e s de cromosomas l a r g o s , 8 pares de cromosomas pequeños y ausencia de a c r o c é n t r i c o s . Así mismo los c a r i o t i p o s reportados para R. b e r l a n d i e r i de Morelos y S i n d o a (Agui lar , 1983) y ahora los de T l a x c a l a , t i e n e n un a r r e g l o muy semejante.
R. r idibunda (Mores-
Comparando las medidas r e l a t i v a s de los cromosomas de g . b e r l a n d i e r i de la poblacidn de Tlaxcala con l a s medidas de los ejemplares co lec tados en los Estados de Norelos y S i n a l o a ( tablas 7 y 8 ) , s e puede a p r e c i a r l a s i m i l i t u d entre las longitudes t o t a l e s de los cromosomas 5 , 6 , 7 , 8 y 11 (4 .96 , 3.3, 2.9, 2.78 y 2.3 respect ivamente) ; t m b i k n hay parec idos e n t r e las longitudes de los brazos largos de los cromosomas 6 , 8 , 9 , 11 y 13 (1.8, 1.6, 1 . 5 , 1 .4 Y 1.2) Y por úl t imo e n t r e l e s longitudes de los brazos cortos de los cromosomas 8 , 11 y 1 2 (1.17, 0.9 y 0.7). En resumen los datos coinciden en una, dos y hasta en t r e s de las c a r a c t e - r í s t i c a s tomadas en cuenta para c o n s t r u i r e l idiograma.
Con excepcidn de l o s cromosomas 2 ( t a b l a 8 ) y 1 2 (tabla 6 ) que s e encuentran en los l í m i t e s para s e r consi - derados como submetecéntr icos , todos los demás t i e n e n una c a t e g o r í a b i e n d e f i n i d a se& e l c r i t e r i o propuesto p o r Levan ( 1 9 6 4 ) para c las i f icar cromosomas humanos.
La c o n s t r i c c i d n secunaar ia en l o s brazos l a r g o s d e l p a r n h e r o d i e z aparece t a n t o en los c a r i o t i p o s de las ranas de !risxcda como en l o s de Plíoreios y S i n a i o a , as í como en otras e s p e c i e s d e l género v. gr. R.ridibunda, R. lessonae, - B. e s c u l e n t a , e t c . (Horesca lch i , 1967; Koref- Sant ibañes , 1980 1.
Haste ahora e l análisis cromosdmico r e a l i z a d o entre les poblaciones de ranas leopardo de nilorelos, S ina l08 y Tlsx- c a l a solo hen revelado semejanzas entre si, s i n embergo aÚn quedan p o r h a c e r o t r o s es tudios que ayuden a a c l a r a r l a posi- c i ó n taxondmice de é s t a s r e n a s , p o r l o que nosotros también propusimos el bandeo cromosdmico como o t r o medio para

Tabla No. 8: Medidas r e l a t i v a s promedio de los cromosomas de R. b e r l a n d i e r i (complemento haplo ide ) de los E s t a d o s d e Morelos y Cinaloa. (Agu i la r , 1983)
’ ‘ No. de Longitud Longitud Longitud Proporción C l a s i f i c a c i 6n . . cronosoma t o t a l de l brazo d e l brazo de brazos
corto l a r g o 1 . -
. <. 1 7.27 3.34 3.93 1.1 m
... 2 6.21 2.42 3.79 1.7 m-sm
3
4
I ..
5
6
._ 7
8
9
I .
-*
._
. .. 10
,. I
11 r - I - 12
r- 13
5.51
5.44
4.96
3.30
2.96
2.78
2.55
2.59
2.33
2.17
1.87
2.17
2.04
2.05
1.42
1.22
1.17
0.97
1.07
0.90
0.77
0.61
3.40
3.40
2.91
1.88
1.74
1.61
1.58
1.52
1.43
1.40
1.26
1.6
1.6
1.4
1.3
1.5
1.3
1.6
1.4
1.6
1.8
2.0
m
m
m
m
m
m
m
m
m
sm
sm

I. i 1,
, .
...
. ,~
. ..
, .. . .. , ,.
" .- e-
..- * -
1 -
1.-
1- 1-
I -
.L
encontrar d i f e r enc ias ( o semejanzas) entre l o s car io t ipos .
Los mecanismos de bandeo cromosómico est& re la- cionados con una d i f e renc ia en e l contenido de proteínas histonas y no histonas en l a s d i f e r en t es regiones de l o s cromosomas.
Se emplean tratamien$os tan variados como son e l uso de urea, KVm0 t r ips ina , etc., en l a obtención de bandes cromosómicas; s in embargo se observd que estas sustancias no producían cambios en cromosomas de an f ib ios (Ychmid, 1978; Manago, 1980 y B i r s t e in , 1982).
4'
Bi rs t e in también reportó que era más frecuente l a l o c a l i z a c i ón de unas cuantas bandas en cromosomas de urodelos que en anuros, y real izando una comparación entre l os patro- nes de bandeo cromosbmico, t a l l a d e l genoma y organizacibn de cromatina en peces, u i f i b i o s , r ep t i l e s , aves y mamíferos, concluyó que l a estructura cromosómica en estos animales d i f i e r e en su contenido y organización de DNA, no pudiendose l o g r a r e l bandeo cromocómico en anuros debido a l a l t o grado de compacteción y e sp i ra i i z ac i ón de sus croinosoma~.
'La información que se ha tenido acerc i de l a t in- c ión d i f e r e n c i a l en croinosoinas de anuros es i 'ue éstos después d e l tratamiento con t r i ps ina adquieren una apmienc ie de e sp i ra i i eac i ón que desaparece conforme avanza i s técnicE y l o s cromosomes se deformnn s i n que se puedm observar bendas (B i rs te in , 1982) .
Fenómenos muy semejantes se presentaron cuzndo nosotros intentamos bandear cromosomas usando primero l a t é cn i ca descr i ta por dang (1972) y luego 18 de C e r r i l l o (1984) . Los e f e c t os de l a d i ~ e s t i ó n fueron menos drást icos al usar e s t e 6 l t imo método y la res2uesta de l o s croaosoinas ternbikn fue rn6s uniforme observandose unas regiones ints teiiidus que Otras.
Pinalmente siguiendo l e técnica descr i ta por C e r r i l l o pero variando le concentrzción de t r i ps ina (de 0.2 a 0.2% ), e l tiempo de exposición (de 3 a 10 minutos) y observandm l ~ s preparaciones, antes de t e ñ i r , a1 microscopio de contraste de fases , para asegurarnos de que l o s cromosomas estuvieran suficientamente hinchodos, se pudieron reconocer de 3 a 5

1 ...
1: < *
1-
+ -
I_
1 ”
bandas b i e n d e f i n i d a s en d i f e r e n t e s reg iones de l o s cro- mosomas siendo f á c i l de i d e n t i f i c a r a l o s cromosomas homologos.
Con e l f i n de obtener cromosomas con d i s t i n t o s grados de compactacibn inyectamos c o l c h i c i n a a l a s ranas y las s a c r i f i c a m o s después de 10, 6 , 4 y 3 horas. Con 6 y 10 horas de c o l c h i c i n a s e obtuvo un mayor nilimero de m i t o s i s y a l disminuir e l tiempo a 4 y 3 horas obteníamos un número menor, pero de mejor cei idad. Siendo é s t a s l a s m i t o s i s que usamos per@ cjbtener bandas, mientras que las primeras fueron usades para determinar e l número cromosómico.
Ahore que s e t i e n e una metodologís para obtener bandas cromosómic~s s e podran conocer los patrones de bandeo, esperendo que a e s t e n i v e l s e puedan poner de m a n i f i e s t o l o s p o s i b l e s cambios entre l es pobleciones de r m e s leopardo que habi ten en l e República. hlexicana.
Con e l f i n de c o n t a r con bases s u f i c i e n t e s para a c i e r a r l a p o s i c i ó n taxonómica de 16s renas leopardo, e l es tudio cromosómico s e r € complementado con e l a n á l i s i s de p r o t e í n e s por medios bioquímicos.
.,-
.I-

CONCLUSION
mea6 determinado un número dipioide de 26 cro- mosomas para R. berlandieri de Tlaxcala. -
La elaboración y análisis del cariotipo revel6 la presencia de 5 pares de cromosomas largos, 0 pares de cromosomas pequeños, una constricción secundaria en l o s brazos largos del par número diez a s í como ausencia de cromosomas acrocéntricos y par sexual.
Se realizó el idiograma expresando el tamaño de cada cromosoma como un porcentaje de la longitud total de l o s 13 pares de cromosomas.
Todos los cromosomas quedaron clasificados como metacéntricos con excepción de los pares 2, 12 y 13 que fueron designaaos como submetacéntricos.
Quedd establecida la técnica para obtener bandas G con tripsina.

, .. . I _
I .
. "
r ' 1.
I I [': I.:: IL I- 1: ,. )I-- .-
1 . _ 1" I:
.-
' I-
RESUMEN
Se r e a i i z d e l an4iisis cromosómico de ias ranas leopardo que h a b i t a n en Tecomalucan Esthdo de T laxca la .
Se obtuvo m a t e r i a l cromos6mico a partir de c é l u l a s de sangre , e p i t e l i o i n t e s t i n a l y méaulk ósea , con e l c u a l s e determind e l número cromosómico (2n= 26) ; 5e elabord e l c a r i o t i p o y e l idiograma resul tando s e r semejantes a los reportados para las poblaciones de Morelos y Sinaloa.
Ai quedar e s t a b l e c i d a l a t é c n i c a p a r a obtener bandas cromosómicas G s e puede cont inuar e l análisis cro- mosómico con e l f i n de obtener los patrones de bandeo para las d i s t i n t a s poblaciones de ranas leopardo y de é s t a manera d e t e c t a r sus p o s i b l e s d i f e r e n c i a s ya que hasta ahora son i n s u f i c i e n t e s los datos para d a r una conolusidn a c e r c a de su p o s i c i d n taxondmica.

r- A N E X O
.L
S

I-- %
...-
, ...

c I
r h.
r c
c
r: a:
J"
O Q B
:? s! I D -
" ? 8 0
3 a ! '
7! '? u1 .w
1 < 1 *
.n ni
1c ?? n .a

Aguilar, M.A.,
BIBLIGGRAFI A
B e j e r , E., h e r n h d e z , P. and B e t 1984. Chromosome analysis o f XV i n t e r n a t i o n a l Congres of Genetics , P 143,.
B e j a r , E. 1981. Determinacidn d e l ndmero cromosdmico y e la - borac ión d e l c a r i o t i p o de b e r l a n d i e r i f o r r e r i y comparacidn de e s t e con e l de -a b e r l a n d i e r i t r i l o b a t a . Reporte d e s e w i c i o s o c i a l . U.A.M.- I z t a p a l a p a , Bléxico.
B i r s t e i n , V. 1982. S t r u c t u r e 1 c h a r a c t e r i s t i c s o f genome organizat ion i n Amphibians : D i f f e r e n t i a l s t a i n i n g o f chromosomes and s t r u c t u r e . J.Mol. Evol. $: 73-91.
C e r r i l l o , M. 1984. Comunicacibn personal .
D i Berardino, M. 1962. The karyotype o f Rana p i p i e n s and i n v e s t i g a t i o n of i t s s t a b i l i t y during embryonic d i f f e r e n t i a t i o n . Dev. B i o l . 2 : 101-1026.
Frost, J, 1982. Funct ional g e n e t i c s i m i l a r i t y between geogra- phically separated populations o f mexican i e o p a r i f r o g s ( W R p i p i e n s complex). syst. 2001. 3 i ( i ) : 57-67.
Coin, C., Goin, O., Zug, E . , 1978. In t roduct ion .to herpe- to logy. Freeman and Company. U.S.A.
Koref-Santibañes, S. end Gunther, R. 1980. K a r i o l o g i c a l s t u d i e s i n - Rana l essonae , H. ridibunda and t h e i r hybrid g. "esculenta". Genetica 22/23: 195-207
Levan, A . , Fredga, K. end Sandberg, A, 1964. Nomenclature f o r centromeric p o s i t i o n i n chromosomes. Hereditas 52: 201-220.
L i t t l e j o h n , M.J. and Oldham, R.S. 1968. Rana pipiens complex: Mating cal1 s t r u c t u r e and taxonomy . Soience 162: 1003-1005.

I.. .
r* I
1- ..
P"
Manago, J . N . , Spine, F.L. and Becker t , W.H. 1980. D i f f e r e n t i a l e t a i n i n g of cul tured bone marrow c e l l chro- mosomes of Rana pipiens . C y t o l o g i a 4fi: 561-569.
Morescalchi , A. 1967. Le r e l a z i o n i tra il c a r i o t i p o d i Anuri d i p l a s i o c e l i . I. I1 corredo cromosomico de a l c u n i l3anide.e. Caryologia g : 6 5 - 8 5 .
-------------- 1973. Amphibia. C h i a r e l l i , A.B. and Capana, E. Cytotaxonomy and V e r t e b r a t e &Evolution. Academic P r e s s . Great B r i t a i n .
Rudich, H.J. 1980. S i n o p s i s b i o l ó g i c a y de explotac ión sobre i ~ l s p r i n c i p a l e s espec ies d e l género e México. T e s i s P r o f e s i o n a l Fac. C ienc ias UNAM, México.
en
Sanders , O. and Smith, H. 1371. Skin t a g s and v e n t r a l meia- nism i n the R i o Grande leopard frog. J. Herpebol. 2: 31-38.
Sanders , O. 1973. A new leopard f r o g (a b e r l c n d i e r i brovi- norum) from Southern Mexico. Herpetolo&ca I ( 2 ) : 87-92.
Schmid, Id. 1978. Chromosome banding i n Amphibia. I. Consti- t u t i v e heterochromatin and nucleolus organizer reg ions i n Bufo and H.yla. Chromosoma @:361-388.
W a n g , H . and F e d e r o f f , S. 1972. Fac tors a f f e c t i n g t h e q u a l i t y o f G bands produced by t r y p s i n i n human chro- mosomes. Cytobios 6: 47-54.


aterminactbn del- r;Om o +crwnomko, elaboración - - -
cariotipo, idicgrama y bandas cranoshicas de la poblaCi6n de Rana berlandieri del - Estado de Tlaxcala.
JUSTIFICA@ Y NATURALEZA DEL'pROVECTOi
- -A I 8 C = r a ' n a a - a - ~ x t c a f 3 a s - S ~ s ha -tder*iflcadoexckt
- __ - - - -
- - . -,-- sívamente con base a sus caracterfsticas morfólbgicas tales Corno forma y tamaño de las manchas dorsales, amplitud intefiorbttal forma de los pliegues dorsolarterales, etc.. tertsticas son insuficientes para determinar st el grupo está formado por una sola, varias espscies. o si son subspecies, ya que es muy amplia la variedad de colorss, hbitos, y en general todas las cara- terfstícas.morfol6gicas entre los individuos de los distintos climas y localidades del terrttorio mexicano.
Se ha visto que estas came-
- Es por esta razbn que se piensa recurrir al andlists cariol&
gico, esperando encontry de esta manera dilierencias más precisas p&a separar al complejo grupo de ranas leopardo que habitan en el país. ._ - _ - - . _--
- I - - INTRODUCCION;
- Ragerlandieri típicamente es una rana delgada verde con un
patrón dorsal de manchas obscuras ovaladas o redondas con bordas descoloridos, algunas veces las manchas son tenues o est& ausentes, existen unos pliegues dorsolaterales bien definidos, gereralmentt3 descokortdos, discontiwos y con una linea sobre la mandíbula supe- rior, En partes áridas de su distribucibn, las manchas son menos notorias, los machos tieren sacos vocales externosj la piel es a me- nudo verrugosa, es bastark comGn la pigmentactbn obscura srjbre -las superíicies ventraies. (WWch, 7- :- - .. . .~
,
-. Rana berlandieri comunmente también se le conocs corno reina c_
leopardo o rana tigre (Rudkh, 1980). .Su-t~.~om>&-gs: Reino Animalia" '
.. - Subreino Chordata Subphilum Vertebrata .
- . . . _ _ ,-
Superclase Tetrapoda- -. . -- Clase Amphtbta . , ;
. ~ I..
Orden Anura Suborden Diplastcoela Gknero Rana Espede.Rana berlandieri (Lfneus, 1968)
! I . .
i I

I
..
Dependiendo del territorio donde habitan las ranas leopardo de MtSxico, éstas se han dividido en subespecies, de tal mahera
se les co~3ce como subespecie 5. berlandieri bei-landieri,_las que se encuentran al oeste se le llama subespecies. bertaderi forreri y por Glttmo la subespecie & berlandieri brownorum que se extiende hacta el sur de México.
que las ranas localizadas al noroeste de la Redbtica MextcaM ._
(Sanders y Smith, 1973). - . . - .- - _ ,. Las interrelactones de las poblaciones del complejo-
.pipiens, dentro del cual se incluya a = berlandteri, sg,esai,, . _ diadas por dos tipos de biblogos los tax6nomos-y los evoi+ianistasL; quienes según Moore (1969), deberfan considerar al organismo .. - entero y no solamente caracteres fenotipicos unos y genotípicos 'otros; y, si se quiere conocer el significado evolutivo de los caracteres de diPerentes especies, deben estudiarse quellos que son conce- cuencia natural y, de aquf que sean adaptativos.
espectes {ntimamente emparentadas. SU territorio de citstritwcth se extiende desde el sur de Canadá hasta México y CostaRica. Las razas que habitan las regiaws m& septentdonales tienen lf- mites de temperatura mfnima y mhima para el desarrollo embrícr nario normal, en comparaci6n con las correspondientes a las risaas más mertdlonales. Las ranas del norte se desarrollan m6s rápida- mente que las del sur a temperaturas bjasj pero a temperaturas m& elevadas tal difeferancia disminuye y aún llega a invertirse. La variabilidad de los hbridos entre las razas de la especie R. pipfens parece estar en correlaci6n con su capacidad de adaptacibn a dtver- SEIS condtdones climátlcas.
- _ ~
_.
El nombre R. pipiens agrupa a un complejo de razas o de I
-
, I '
Se ha observado que entre las ranas y los sapos, los estfmu- los uditivas desempeñan un importante papel en la congregacióQle hembras y machos de la misma especie en los sitios donde s8
aparean. Durante el parfedo de reproducct6n, el canto de 10s machoc es es co para cada especie. E l papel de I stfmulos audltivos en eí tento reproducttvo & las especie sido e&u&ado en particular por W. F. Blair (1958 y 1964). El canto es estudiado a través <'.-
-. de espectrogramas de sonido (Dobzzhansky, 1970). -.
Este llamado se produce en5dos sacos membranosos alojados sobre l& tubos que sirven a la respiraci6n pulmonar;ellos vibran al recibir y arrojar alternativamente el aire de los pulmOnes, re- sonando & diversas manwas (Rubfn, 1981).
Littlejohn y Oldham (1968) han obtsnido con un asctlogratb las gráficas del canto de apareamiento. Enel complejo piptams

.f* se har.realizado estudios sobre la estructura del canto que han contri- bufdo ~. al conocimiento del apareamientp y a la taxonomía del grupo. .
~ ~ ~~ .. ~. ~ ~
.. La variabilidad geogr&ffca en la estructura del llamado indica que hay grandes poblaciones alopátrtcas de rana leopardo que estan presew tes en el centro de Estados Unidos. Estas formas parecen mantener sus distinciones en pequeñas zonas de simpatrfa y la mayoría de los machos adultos pueden ser separados morfOl6gicmnte. - - . - - .- <*
LOS intentos para clasificar a las ranas leopardo a través-& la morfolcgfa del adulto y los ensayos de hibridaci6n artificial, han MXT ducido a la aceptación de la idea de que es una sola especie nOrma1 de amplia distribucfóo con ntveles de incompatibilidad gedtica entre las poblaciones geográficame&.
Mecham (1969) ha hecho cruzas experimentales entre ranas del grupoR. pipiens: Lpalustr is , & areolata, R. meaapoda, R. m a - zumae y complejo R, pipiens (del cual us6 tres %emiespecies": berlan- dieri , sphenacephala y de plano sur). Las cruzas demostraron un alto nivel de compatibilidad gedtica para la mayor parte de las combinade res; la incanpattbilidad en cruzas dentro del grupo de especies& &- p2encpuede ser influido por diferencias en la adaptación de temperatura entre otros factores. El hecho de que exista baja compatibilidad deR. beriandieri en cruzas con- mcmtezumae, R. meaapoda, R. areolate y (en men* grado) R, pa3ustris, en comparaci6n con otra-krmas deR. plpfens, a d como laincompatibilidad con laE.2 . pipiens dsl nwte y la forma del plano sur apoya fbertemente la separacih de fi berlandieri como una especie distinta.
.-
. . . . . . . . . . .
, 1

1 C . , .L., < < ~ - . -~___- ' , I 8.I .
.. 3 .
. . .~~~ .~ ~~~ ~. ~~ ~. I-~ .~
ANTECEDENTE Sr
Se han bcho algunos estudios para demostrar laR dkfl.'mtas entre las cuatro pobiactones alopátricas de ranas leopan acuerdo con l a forma de pliegues dorsolaterales así c(xIV1 t~ly'spncta'
en 1971 Sanders y Smith separaron a las ranas leopardo del grupo Rana pipiens de los Estados Unidos citando Ins tiiiuron- cias morfbl6gicac entre los dos. grupm, espctalme&o el t~W't*'tnmo ventral. '
dieri. - en estudios posteriores (Eejar, isaij.
A las ranas leoparda mextcanas se les ha tderitiflCfltli' base en la distribuci6n de manchas y forma de los plk%W'S rk'rso- .5 ' laterales de tal manwa qua la especie Rana berlandfort pillA 'pmx- .
,,q+mdle - da ;GOT tres subespecies: R. berlandieri bgrlandferi , 5. .!...
,,>, .(ti'. , - a s
r4l*'I'r4 que O ausencia de oviductos vestigiales en machos,..--.- tal 5 % 1.-
___ .T
Al grupo mexicano se le di6 e l nombre de ~ ~ ~ ~ A I H ? ! - % e RJ n ya& Esta d iv ts ih como dos especies distintas ha si&
.~ ~
. .
. ~~~~~~~ .. . .. ~ ~ ,~ ... ~~~ . . ~~~~ ~~. ~~~ ._ ~ ~~~ ~~~ ~ ~ ~~~~~ ~ ~ .. .~ ~ ~
'
.. i
- . . > ri y - R. berlandieri forreri. CSanckrs, lW3). . . 1
dorsales a s C como otras marcas morfo16gglcas ayudan mUY I )<Woo para separar a las ranas, ya que se observa variabilidad aGn r t t d 1 W id( : ~
~
c
,b carat---. Para cada subespecie se ha determinado un p a t h terfStiCaS morfol6gicas, sin embargo se ha visto que lar> l ~ l f l l K ~ e : -:.I
~
vtduas & una misma localidad (Sanders, 1973). sari0 obtener otro medio de referencia para poder est&iryter Hi e' grupo de r a m leopardo en verdad está formado 'por trOb R ~ d n s ~ c ' e s
H a ~ d W k ) ~ " no--'-
,como se dtjo anteriormente. i ' O

- \ _. . . . ,’
.........
* , Determinar e l nGmero cromos6mic0, obtener los cariotipbs,
idicgramas y bandas crcinoshicas de l a poblaci6n de Rana - berlarr -del Estado de TEaxcala.
, . - -_ - .. - -
FRoGRAIvL4 Y NETODOLOGIA E L TRABAJO: - - _c . - -. -
1.- Obtencih de material biolbgico: Las ranas serán trafdas del Estado de Tlaxcala, la colecta
._ se hard durar&= la noche de preferencia en ausencia de luna, el %mino se alumbrará por medio de una l h p a r a con e l ffn de cegar a las ranas momer*aneamente, queden inmbviies por algún tiempo y puedan ser capturadas por medio de una talega.
.
-.- 2.- Obtencih de cromosomas: Las ranas ser& inyectadas intrapsritonealrnente con una
. -- _I
Solucitm ds colchicina al 0.30/., para ser sacrificadas desptks de aproximadamente 10 horas.
El material cromos6mico se obtendrá a partir de mddula h a , Sargre y epitelio intestinal.
El método para extraer sangre es el siguiente: tas rana5 con &ter et(lico y coiochdolas en una charola de diseccibn se iimpia y corta la región ventral ramoviendo e l pericardio para dejar expuesto e l corazón. Con una jerirga previamente t-epat-inizada se hace una pindh cardiaca con e l objeto de extraer la sangre la &al se& sembrada en frascos &pula con medio Mc Coy. Luego se incubar& a temperatura ambiente de 96 a 168 horas. 1’
E l método para extraer médula &a es e l siguiet-d: Se hacen las disecciones de fdmur, hGmero y tibiofhula, se cortan las epfPlcts
eL medio se colecta en tubos de centdíúga.
Extraer e l inbeetino, macsrarlo con navaja e incubarlo con trtpsina a 37 C durante 20 minutos, hacer pasar 1-ic &lulas por una malla y colectarlas en tubas de centrhga.
Una vez que se tenga la médula &ea, sangre o epitelio intestinal en tubos de centrffuga se procede a la obtencibn de cromo somas según l a técnica descrita por Bogart en 1970.
anestesiar a
. y se extrae la médula 6sea con medio Mc Coy. La ddu la con
E l método para extraer epitelio intestinal es e l siguiette:
- 3.- Determinacibn del &mero crcinosbmico: Se hacen preparaciones por e l método de secado a la flama,
con e l material obtenido a partir de las tres tkcnicas mencionedas. Las preparaciones se revisan ai microscopio de contraste ds fases
. . . . . . . . . . ... ~. . , . . . / L ’ I ~ . .

t ! I
t . ' Los números cromosómicos por mitosis se registrm, detierml
-. ~ . .I ir?. .:.tipos , &dose el danero modal.
~ ~. . .. .. ~~ 8.- Obtend6n del caFi&tpO: ' ~ ~ ... . _. h r l m
Se seleccionan las mejores mitosis de cada individuo -
(cai número cromochico canpkto: dmero modal) a dichas mitbts se les tomarán fotografías. y se ordenarán por pares de mayor a menor tamaño.
Se cortarán 10s c r o m o s o H ¿ S ~ ~ d é - T ~
5.- Oblxmcibn de bandas cromosbmicas: :: . ;i*cta La t$cnica que se realizará es Bandas G c m trtpsiw 31
pang, 1912). 37 C durante 6 segundos aproximadamente, lavar con epnol :Wlr:7$% ~ .~
y con alcohol absoluto, se deja secar la preparacibn al 'Mm, luego se tiñen los cromosanas cm Wright y Giemsa, por Gltímo lavar ' con metano1 y agua destflada.
ss introduce la preparacih en tripsim ai ü.O4%ear .. :~
,.,a . ~. ~ . - . . - , 6.- Obtención dsl idiograma: . ~.~ ~-
Se obtienen las medidas de los brazos de cada cranosma, sacando el promedio entre los cromosanas homblogos para obtcnt-,i, una medida relativa de su. tamaño. i . ~ = ,. L _- - - _. :.. . . - I . , _ _
.2 .. 1 .. _I
. . . - 3
~~ I . . . -. - - - > . .. , - ...
i I - LI-. ~ . , .._.:*.4 -

.,
I ' .
I . .i
BIBUOGRAFiA:
Befar, 0; E. 0981). Determinaci6n del nhem cromosbmico y ebb- racib del caridipo & - Rana berlandieri fwreri y canpara ci6n de éste con el & Rana berlandieri trllobata Servicio Social UAMl.
. . .
Bogart, J.P. (1970). Systematic prcblems in the b i l y .
lidae (Anura) as indicated by @ryotipic analysis. Cittoge-- netics, 9: 3 8 s 303.
Dcbzhansw, T. (LWOJ. Gen&aa del Proceso Evolutivo. Ed. Exbem - * ~~ Naw Yo* .
Littlejohn, M.J., Oldham, R.S. (1968) Rana @piens Canplex: mating call stwcture and taxonomy. Science, - 3857 : 1003- I-.
Mecham, S.J. 0969) New information from ExperlmerSal Crosses on Genetic Relationchips witMn the RaM pipiens species group. J. EXP. 20bl. 165: 1- 26.
Moore, A.J. (1969) Dfploid and Haploid ylbrioization of Difirent poe lationc of tty Rqna pipiens Canplex. Experiments wtth &ma les from tvkdeo. J. Exp. 2001. _. 170:169-180.
Morescalchi, A. 0973) AmphiMan "in" QtCtaxonomy and Vertebrate Evolution. Academic Press London, New 'fork, p 269-348.
MorescalcM, A. 0977) New .Developments in Vertebrate qotaxonany. 1.- QtotaxaiOmy of the amphibians. Gem~Ca, 1-e(9):174. 193.
Rubin, R. (1981) La rana y su explotactbn. E C S , bl&xfcO.
Sanders, O. Smith, H. (19il) Skin tags and ventral melanism ln the Rfo Grernde leopard frogs. J. Herpetol, - 5: 31- 38.
Sanders, O. (iW3) A new leopard frog -a berlaridieri brownorum) from Southern Mexico. J. Herpetol, 1. (2): 87-92.
. .. . , . . . .
. . .
, .
. . . .
. . . ,
. . . .. , .
