1. introducciÓn -...
TRANSCRIPT

1. INTRODUCCIÓN
El cerezo Prunus avium L. es una especie que ha experimentado un gran aumento en
superficie, con 4.600 hectáreas (DECOFRUT, 2001) además se observa un
desplazamiento hacia zonas donde antes no se cultivaba.
El cerezo es una especie que se clasifica entre aquellas de alto requerimiento de frío,
dependiendo del cultivar, requiere entre 700 a 1400 horas con temperaturas bajo 7 ºC.
Por ello, entre los aspectos climáticos que se deben considerar, está la cantidad de
horas frío que caracteriza al sector elegido y que son necesarias para las plantas.
Las plantaciones de cerezo en zonas con inviernos benignos, presentan una brotación
desuniforme. Además, la floración de los cultivares tiende a desfasarse afectándose
con ello, fuertemente, los rendimientos finales.
Debido a esto, es que en las plantaciones en la IV Región, se requieren utilizar
cultivares de bajo requerimiento de frío invernal. Además, es preciso la utilización de
Cianamida Hidrogenada para suplir la falta de frío invernal, adelantar y homogenizar
la floración y posterior brotación.
Este trabajo se justifica en las ventajas esperadas del cultivo en esta área: cosecha
temprana, con lo que se logra mejores precios y ampliar el período de oferta del país a
los mercados externos. Por otra parte, aumento de la producción al mejorar la
floración de la especie.

Uno de los aspectos fundamentales para la producción de cerezas en la IV región, es
determinar el momento de la aplicación de Cianamida Hidrogenada, puesto que se
asocia con la cantidad de frío acumulado.
El presente trabajo pretende determinar el efecto que tienen diferentes acumulaciones
de frío, al momento de aplicar Cianamida Hidrogenada en la floración del cerezo.
Para esta investigación, se asume que el momento de aplicación de Cianamida
Hidrogenada, tiene un efecto sobre el adelanto y concentración de la floración de los
cultivares de cerezo, ‘Brooks’ y ‘Newstar’.
El objetivo general de esta investigación es:
Determinar el efecto de distintas acumulaciones de frío, al momento de la aplicación
de Cianamida Hidrogenada sobre los eventos florales del cerezo en la IV Región.
Como objetivos específicos se plantean:
Determinar la acumulación de horas frío, para la aplicación de Cianamida
Hidrogenada, en las condiciones de Mialqui, provincia del Limarí, IV Región, en
función de una mayor expresión floral, regular y abundante, para 2 cultivares de
cerezo.
Determinar el adelanto de la floración, con distintas acumulaciones de frío al
momento de aplicar Cianamida Hidrogenada sobre dos cultivares de cerezo dulce.
Salto de página
2. REVISIÓN BIBLIOGRÁFICA

2.1. Características botánicas
El Cerezo o guindo dulce (Prunus avium L.), pertenece a la familia de las Rosáceas.
Posee flores simples, de color blanco, con cinco pétalos y cinco sépalos, además de
un número indeterminado de estambres, estas se agrupan en racimos llamados
corimbos de uno a cinco unidades, el fruto corresponde a una drupa (SOTOMAYOR,
1995a).
Pese a que las flores son hermafroditas, casi todas las variedades de cerezo son auto
estériles, pues presentan incompatibilidad gametofítica, lo que hace necesario la
polinización cruzada para lograr producciones adecuadas (SOTOMAYOR, 1995a).
Dependiendo de los cultivares, se tiene una mayor o menor densidad de flores. La
floración depende, de varios factores, entre los cuales están, las condiciones
climáticas del invierno, y otros factores ambientales, como el largo del día,
temperatura y humedad, que inducen al estado de latencia o letargo que tendrá el
árbol durante el invierno (ADKINS y ROSS, 1981).
2.2. Estado de latencia o letargo
Es un estado en el cual un organismo vivo con aparente signo de inactividad, cuyo
crecimiento visible ha sido suspendido temporalmente por cualquier causa. Por la
definición anterior, y también etimológicamente, quietud o receso serian sinónimos
de latencia (GIL, 2000).
Atendiendo a la causa de la inactividad visible pueden distinguirse diversos casos de
latencia (GIL, 2000).

2.2.1. Inhibición o latencia correlativa
Es la falta de crecimiento de las yemas por acción inhibidora de otros órganos; por
ejemplo, las yemas laterales de un brote que no crecen en primavera o verano
(mientras crece el brote), por acción de la yema apical o de las hojas, pero tan pronto
estos órganos son eliminados (despunte, defoliación) la brotación se inicia
(ROMBERGER, 1963).
2.2.2. Quietud o latencia ambiental
Llamada también “eco-inactividad”, las yemas no brotan por ausencia de condiciones
ambientales tales como: baja temperatura, fotoperíodo, déficit hídrico, etc., pero si
ellas se normalizan, la brotación ocurre (GIL, 2000).
2.2.3. Letargo o latencia endógena
Llamada también “endo-inactividad”, el crecimiento visible de las yemas no se
manifiesta aunque, hayan condiciones ambientales propicias y no existan otros
órganos inhibidores, ya que se debe a factores propios del órgano, la propia yema
debe superar ese estado por cambios fisiológicos complejos, lo que ocurre
normalmente durante un período de exposición al frío (GIL, 2000).
2.2.4. Características y evolución del letargo
Las especies frutales de hoja caduca se han adaptado naturalmente a lugares con
estaciones climáticas bien marcadas. El letargo y la dureza de los tejidos, dos

condiciones que se desarrollan paralelamente, permiten la supervivencia de las
plantas en condiciones de frío adversas, hasta el advenimiento de mejores
condiciones. El estado de letargo, es la incapacidad endógena de una yema para
crecer y abrirse dando un brote o una flor, desaparece por una exposición a
temperaturas bajas por un tiempo determinado; estos dos factores caracterizan a las
especies, y más aun a los cultivares (TINKLIN y SCHWABE, 1970 y ABBOUT,
1969). Se desencadenan en el otoño, cuando el árbol se desprende de parte de su
follaje (un 70 %). Se caracteriza por una paralización de toda actividad metabólica, la
cual solo es reanudada a fines del invierno (RAZETO, 1999).
La latencia y el letargo de una yema evolucionan gradualmente. La primera
inactividad aparente de una yema se debe a la inhibición correlativa originada en el
ápice en activo crecimiento y también en las hojas adyacentes. La competencia entre
númerosos centros metabólicos puede causar la cesación del crecimiento apical en
cualquier momento de la temporada (NIGOND, 1967). Hacia el final, los cambios
climáticos, como el descenso de la temperatura o el acortamiento de los días, pueden
agregarse para mantener a las yemas en un estado de quietud, como se ha probado en
la vid (SCHNABEL Y WAMPLE, 1987).
La yema se va aletargando progresivamente en intensidad, al comienzo es suave y la
brotación puede ocurrir, aunque con dificultad, en respuesta al calor, la poda, heridas
y deshoje, es el letargo preliminar, el frío en ese momento contribuye a profundizar
su intensidad, luego, el estado de aletargamiento se va haciendo cada vez más intenso
hasta su condición máxima: el letargo profundo. El frío en esta etapa contribuye a su
disipación llegando a un letargo final, que es cada vez menos intenso, después de una
mayor exposición al frío, con mayor capacidad de responder a condiciones
estimulantes. El terminó del letargo (tiempo mínimo para iniciar brotación normal en
condiciones ambientales favorables), depende de la exposición a bajas temperaturas,

se llega a un post letargo, capaz de brotar si existen condiciones apropiadas, tanto
ambientales como correlativas de la planta, de no ser así, de acuerdo a AMEN (1968)
citado por DENNIS (1994), la fase de mantención del letargo continúa hasta que las
condiciones ambientales e internas de la planta cambian. Este mismo autor afirma
que la mantención es una condición de estado fijo, con factores responsables de
renovar el crecimiento, que podrían ser promotores de éste existiendo un balance con
aquellos que lo previenen (inhibidores de crecimiento). Hasta que las condiciones
cambien, las yemas permanecen despiertas, pero sin crecer, en un estado de quietud
(latencia) ambiental lo que es confirmado por GIL (2000), en donde el letargo
permanece estático.
En zonas tropicales es posible el cultivo de especies de hoja caduca, siguiendo la
terminólogía propuesta por LANG et al., la "eco-inactividad" es utilizada para
proporcionar el requisito previo de condiciones necesarias para la diferenciación
floral; esto ocurre durante la "ecto-inactividad"; induciendo a un nuevo ciclo de
crecimiento, quitando las fuentes de inhibición antes de que la "endo-inactividad" se
desarrolle. Así, el frío es innecesario: a) la exigencia de frío, de los cultivares y
especies tiene poca importancia en los trópicos. Cultivares con altas exigencias de
frío, potencialmente son bien satisfechos, b) El cultivo de especies caducas, en zonas
con temperaturas uniformes de los trópicos, depende de la inducción del siguiente
ciclo de crecimiento, después de que la diferenciación floral ha ocurrido, pero antes
de que el receso comience (GORDON, 1987).
2.3. Requerimiento de Horas – Frío
Existen varios métodos para calcular la cantidad de horas - frío que requiere el árbol
para iniciar el crecimiento. Sin embargo, el modelo de Utha de RICHARDSON,

SEELEY y WALKER (1974), introducido para durazneros, ha sido ampliamente
utilizado durante los últimos años. Este modelo asigna una unidad de frío a cualquier
hora con temperaturas entre 2,5 y 9,0 ºC. Temperaturas mayores o menores a este
rango restan o acumulan menos frío, ya que el modelo indica que no todas las
temperaturas tienen el mismo efecto fisiológico sobre las yemas (MELGAREJO,
1996).
WESTWOOD y BJORNSTAD (1978) indican que el requerimiento de frío de la
yema, es una característica intrínseca de la planta y depende de la zona de donde
provenga. Plantas originarias de climas templados, con largas estaciones libres de
heladas, tienen un requerimiento de frío relativamente bajo. Por otro lado, plantas
originarias de zonas con inviernos rigurosos, poseen un alto requerimiento de frío
para poder brotar. Dentro del mismo árbol existen diferencia entre los tipos de
yemas, normalmente las yemas florales presentan una menor demanda de frío
invernal para abrir que las vegetativas (RAZETO, 1999).
La respuesta al frío de las yemas de cerezo, varía de un año a otro, posiblemente
debido al desarrollo morfológico de la temporada anterior y a factores externos e
internos, así como la nutrición, la disponibilidad de agua (FELKER y ROBITAILLE,
1985).
Una vez que las unidades de frío (horas de frío) han sido acumuladas, las unidades de
calor aceleran la apertura de las yemas, poniendo paulatinamente fin al estado de
letargo en semillas o, en este caso, en las yemas (SEPULVEDA, 1989).
Especies y/o cultivares con un alto requerimiento de frío requieren de altas
temperaturas primaverales para iniciar la brotación. Por otro lado, especies y/o
cultivares que han satisfecho sus necesidades de frío a mediados de invierno,

requieren solo temperaturas moderadas para reiniciar la brotación (TIMMIS,
FUCHIGAMI y TIMMIS, 1981).
Si las necesidades de frío invernal de cada árbol no son satisfechas, se presentarán en
la siguiente época de crecimiento desórdenes fisiológicos más o menos importantes,
que serán en función del déficit de frío que la planta haya sufrido, dentro de los
fenómenos que ocasiona el déficit de frío están: El retraso en la apertura de las
yemas, brotación irregular y dispersa, la caída de yemas el cual es el efecto más
grave, fenómeno al cual el cerezo es poco susceptible y anormalidades de crecimiento
(MELGAREJO, 1996)
2.4. Cianamida hidrogenada
El interés para el control artificial de la brotación de especies de árboles frutales, de
hoja caduca, esta estrechamente unido con técnicas comerciales para cultivar estas
especies en zonas tropicales, donde las exigencias de frío no son satisfechas
naturalmente (EREZ, 1987).
Chile es un país pionero en el estudio de este regulador de crecimiento en frutales
para modificar la brotación, floración e, incluso, la fecha de madurez de la fruta. El
primer estudio reportado en el mundo acerca del uso de Cianamida Hidrogenada en
cerezo, data a fines de los ’80 (LEMUS, 1998).
La Cianamida Hidrogenada es un regulador de crecimiento que modifica el período
de receso invernal y estimula precozmente la brotación (LEMUS, 2002).
La cianamida cálcica, ha sido usada desde hace algún tiempo para romper el receso
de vid y otras especies caducas en Japón, Brasil, e Israel. Sin embargo, la forma de
pasta y la alta concentración, impidió su empleo como un agente comercial. No antes

del descubrimiento del ácido de cianamida, como un agente activo que rompe el
receso, esta sustancia considerada comercialmente factible. El empleo de ácido
cianamida como rocío fue encontrado valioso para la vid en Israel (EREZ, 1987).
La Cianamida cálcica (CaCn2) en solución, o Cianamida Hidrogenada (CH2N2) se
utiliza como promotor de la ruptura del receso de las yemas de frutales tales como:
vid, ciruelos, perales, duraznos, kiwis (AMBERGER, 1984). En cerezos constituye
una herramienta útil cuando la floración es irregular, por falta de frío invernal en
algunos años. También adelanta la floración y la maduración, que permite
alternativas de manejo en la polinización y en la cosecha. Su efecto en distintas
especies depende de la dosis y fecha de aplicación (LEMUS, 2002).
Otros compuestos tales como aceites minerales, Dinitro Orto-Cresol y Tiourea, han
sido usados con éxito para terminar el letargo en árboles de hoja caduca (SHULMAN
et al., 1983).
Las sustancias químicas, compensan la falta de frío en las zonas subtrópicales, que
han sido probados en los trópicos, sobre manzanos en las tierras bajas ubicadas en
Filipinas, H2CN2, el 2 % i.a. (Alzodef) era eficaz, mientras que el 2 % de thiourea era
el más eficaz en un sitio montañoso. Diferencias de temperaturas pueden ser
responsables, sobre durazneros, mezclas DNOC+ACEITE eran eficaces en el keniano
y el filipino, tierras altas. En el Este, el concentrado DNOC+ACEITE (el 2.2 % y el
10 %, respectivamente) era más eficaz que KNO3, thiourea, o H2CN2 (GORDON,
1987).
AMBERGER (1984), postula que al ser aplicada la Cianamida en dosis adecuadas, se
inhibe la acción de la enzima Catalasa, aumentando el contenido de arginina y
promoviendo la respiración mitocondrial, con lo que se aumentaría la liberación de

CO2. En cuanto a la inactivación de la catalasa, el grupo C=N de la Cianamida
reacciona con el fierro de la enzima peroxidasa para destruir el peroxido del
hidrógeno (H2O2), reduciéndolo en agua y oxígeno lo cual, tiene particular
importancia, ya que en muchas reacciones enzimáticas se produce como subproducto
del metabolismo, dado su efecto tóxico para la planta podría causar serios daños
(SKW TROTSBERG, 2002), el aumento de las cantidades de peróxido podrían
provocar una activación en la vía de las pentosa-fosfato y por consiguiente el término
del receso invernal.
Cianamida Hidrogenada estimula la reacción entre el peróxido de hidrógeno y
ascorbato, lo cual conduce a su vez, a un incremento en la actividad del ciclo pentosa-
fosfato (PPP), por lo que asegura por un lado la reducción del peróxido de hidrógeno
y producción de sustancias tales como lípidos, ARN, ADN, las cuales son
fundamentales para la brotación de las yemas. (SKW TROTSBERG, 2002).
La Cianamida es usada en frutales con distintos objetivos dependiendo de la especie y
de la zona climática en que se encuentre ubicada. Se aplica a las yemas cuando éstas,
se encuentran en el período de receso invernal (FOOT, 1987), usada para reemplazar
horas de frío en zonas donde no se cumplen los requerimientos, para adelantar la
cosecha obteniendo mejores precios, y hacer coincidir la floración de distintas
variedades logrando mejorar la polinización (SOTOMAYOR, 1995b).
La mayoría de los cultivares de cerezo, especialmente ‘Bing’, el más plantado en
Chile, requieren de polinización cruzada para lograr una buena cuaja. No sólo se
requiere un polinizante compatible, sino que es necesario que la floración sea
coincidente para que haya seguridad en la fecundación (LEMUS, 1998).
En Francia, sólo se usa para la variedad ‘Duronne 3’, y solucionar el problema de
polinización con la variedad ‘Noir’ de ‘Michel’ (LEMUS y RIVAS, 2002).

En vid cv. ‘Sultanina’ reemplaza las horas de frío y su resultado es el adelantamiento
y uniformidad de la brotación, obteniendo así, una mejor respuesta si se aplica
alrededor de los 45 días antes de la fecha estimada como brotación natural de las
plantas (SEPULVEDA, 1987).
Investigaciones realizadas por LYON, POLLER y RODRIGUEZ (1989), que
estudiaron los requerimientos de frío de vid del cv. ‘Sultanina’, indicaron que la
Cianamida Hidrogenada tuvo un efecto compensatorio cuando se obtuvo entre el 60 a
80% del requerimiento total de frío invernal.
En ensayos realizados en 1986, en vides cv. ‘Sultanina’, se evalúo el efecto de épocas
y dosis de Cianamida, concluyendo que aplicaciones 76 días antes de brotación
produjeron un adelanto en siete días, pero no se alteró el porcentaje de brotación
(VALENZUELA y LOBATO, 2000).
Esto concuerda con GARDIAZABAL (1988), que en ensayos efectuados en la
temporada 1986-1987 en la zona de Quillota y Los Andes, mostró que hay un
adelanto de entre cuatro y siete semanas con respecto a una brotación normal en kiwi,
cuando se aplica este producto,
Se observó que aplicaciones cercanas a la brotación en dosis altas, produjeron un
atraso de brotación, considerando estas respuesta como fitotoxicidad
(VALENZUELA y LOBATO, 2000).
2.5. Reguladores de crecimiento endógenos

Mucho conocimiento se ha acumulado sobre los efectos de los reguladores de
crecimiento en el letargo de yemas y sobre su ocurrencia durante su evolución, lo que
ha generado hipótesis acerca del rol que ellos juegan (GIL, 2000).
El ácido adscisico (ABA), es un regulador de crecimiento que normalmente aumenta
su concentración en el período invernal de la planta (TIMIS, FUCHIGAMI y TIMIS,
1981). El ABA aumenta en las yemas hacia el otoño y comienzos del invierno,
disminuyendo cerca de la brotación en diversas especies, específicamente en la región
meristemática de la yema y no de las escamas, en relativa concordancia con la
evolución del letargo (GIL, 2000).
Se piensa que el ABA juega un papel importante en la inducción del letargo y en su
profundización, cuando la capacidad de crecimiento es baja, pero no tanto en la
desaparición, la brotación puede ocurrir aún con alto nivel (GIL, 2000).
Indudable que el efecto del ABA esta estrechamente relacionado con el de otras
substancias promotoras del crecimiento, como las giberelinas (AG) las cuales tienen
un efecto opuesto al ABA en el letargo. La aplicación exógena de AG3 ha
reemplazado el requisito de frío para terminar el letargo en durazneros y avellanos,
pero se requiere de mayor concentración a medida que se profundiza el letargo y de
menor durante su disipación. El nivel de giberelinas aumenta después de un período
de enfriamiento de yemas y de semillas (GIL, 2000).
Hay resultados cuya interpretación indica que las giberelinas son el resultado, y no la
causa, del término del letargo y hacia el final del letargo, cuando se amplía la
respuesta a condiciones estimulantes de crecimiento, producen una respuesta muy
parecida al alza de temperatura, actuando como promotoras de brotación.

2.6. Características de los cultivares
2.6.1. ‘Brooks’
Su origen es de Estados Unidos, pose un buen vigor, con un hábito de crecimiento
semi erecto, posee como polinizantes a ‘Ruby’, ‘Garnet’, con una productividad muy
buena (CLAVERIE, 2002).
Buena adaptación a climas áridos. En Francia, la época de floración es temprana, dos
días antes que ‘Burlat’ (CLAVERIE, 2001), en Zaragoza, España, la época de
floración es media, del 29 de marzo al 2 de abril (GARCÍA y FAÑANAS, 2002), en
Quillota durante la temporada del 2001, la floración comenzó el 13 de septiembre
(CORTÉS, 2002), en la misma temporada, pero en la zona de Romeral, en donde no
se realizan aplicaciones de productos químicos para romper el letargo la, floración
comenzó el 14 de septiembre (JIL, 2002), en Quillota durante la temporada del 2002,
la floración comenzó el 23 de agosto al igual que ‘Burlat’ (ÁLVAREZ, 2003).
Características del fruto:
En Francia la fecha de cosecha es de 10 a 12 días después de ‘Early ‘Burlat’, con un
peso promedio de 8 a 10 gr, con un color púrpura, de forma reniforme, posee una
buena firmeza, de fruto muy sensible a partidura (CLAVERIE, 2002), en Zaragoza,
España, su época de de maduración es temprana 10 días después de ‘Burlat’
(GARCÍA Y FAÑANAS, 2002), durante la temporada del 2001, en Quillota la época
de maduración comienza el 15 de noviembre (CORTÉS, 2002), y en la temporada del
2002, comenzó el 5 de noviembre al igual que ‘Burlat’, comenzando 10 días antes
que la temporada anterior (ÁLVAREZ, 2003), en la zona de Romeral durante la
temporada del 2001, la maduración comienza el 19 de noviembre (JIL, 2002).

2.6.2. ‘Newstar’
Su origen es en Canada, posee un buen vigor, con un hábito de crecimiento semi
erecto, con un requerimiento de 520 horas frío aproximadamente (CORTÉS, 2002),
no requiere polinizantes, con una productivida muy buena (CLAVERIE, 2002).
En Francia la época de floración es de -1 a +2 días respecto a ‘Burlat’ (CLAVERIE,
2001). En Zaragoza, España, su época de floración es temprana entre el 27 y 30 de
marzo (GARCÍA y FAÑANAS, 2002). En Quillota durante la temporada 2001,
floreció el 15 de septiembre (CORTÉS, 2002), en la temporada del 2002 la floración
comenzó el 30 de agosto, 7 días después de ‘Burlat’ (ÁLVAREZ, 2003). En Curicó,
florece el 22 de septiembre (JIL, 2002).
En Francia la fecha de cosecha es 12 a14 días después de ‘Early Burlat’, con un peso
promedio de 7,5 a 9 gr, de un color púrpura, posee una forma reniforme, con una
firmeza y resistencia a partidura media (CLAVERIE, 2002).
En Zaragoza la época de maduración es media 11 a 14 días después de ‘Burlat’, entre
el 27 y 30 de mayo (GARCÍA y FAÑANAS, 2002). En Francia su época de
maduración es de 12 a 14 días después que ‘Burlat’ (CLAVERIE, 2002). En Quillota
durante la temporada 2001 la fecha de maduración fue el 20 de noviembre (CORTÉS,
2002), en la temporada anterior la maduración comenzó al iguala que ‘burlat’ el 5 de
noviembre, 15 días mas tarde (ÁLVAREZ, 2003). En Curicó el período de
maduración comienza el 29 de noviembre (JIL, 2002).
2.7. Características del porta injerto

2.7.1. Santa Lucía 64
Pertenece a la selección de P. mahaleb, es árbol vigoroso con 80-90% de ‘Merisier’,
con una entrada en producción más temprana que Prunus mahaleb. Proporciona un
buen calibre de fruto, prefiere suelos secos, cálidos y de textura ligera, dotados de
buen drenaje, tolerante a sequía y a suelos calcáreos es muy sensible a asfixia, y
Phytophtora spp., compatible con todos los cultivares (SAUNIER, 1997).
2.8. Características climáticas de la zona
Mialqui pertenece al agroclima Ovalle, el cual se encuentra desde Vicuña hasta San
Felipe, entre los paralelos 30º y 32°S, la variación estacional en la temperatura
disminuye con la latitud, que disminuye y se acercan a la uniformidad a lo largo del
año en el Ecuador, en los regímenes de temperaturas de la zona templada, que se
encuentran principalmente entre la latitud 50° y 30 °, las plantas poseen el frío
suficiente para romper el receso ocurrido durante los meses de frío.
En las áreas entre los paralelos 30° y 15° que incluye regiones subtropicales, así
como, muchos ambientes de desierto, son típicas en las cuales las temperaturas de
invierno son demasiado bajas para el crecimiento. El receso ocurre, pero el frío es
inadecuado, para la mayor parte de las especies de hoja caduca.
Las áreas entre las latitudes de 15 ° y 0 ° son por lo general de trópicos húmedos o
secos, de temperaturas relativamente uniformes durante todo el año, donde se
acumula poco a nada de frío, según el método actualmente usado de evaluación
(GORDON, 1987).
El régimen térmico de esta zona se caracteriza por una temperatura media anual de
16,6ºC, con una máxima media del mes más cálido (enero) de 28,5ºC y una mínima

media del mes más frío (julio) de 6,3ºC. El período libre de heladas aprovechable es
de 10 meses, de septiembre a junio inclusive. La suma anual de temperaturas, base
5ºC, es de 4.220 grados-días y base 10ºC, de 2.390 grados-días. La temperatura
media mensual se mantiene sobre los 10ºC, entre enero y diciembre.
El régimen hídrico se caracteriza por una precipitación anual de 125.7 mm siendo
junio el mes más lluvioso, con 35.7 mm. La evaporación de bandeja llega a 1.676
mm anuales, con un máximo mensual en febrero de 254 mm y un mínimo mensual en
julio de 52 mm. La estación seca es de 10 meses, agosto a mayo, con dos meses
húmedos, junio y julio.
Salto de página
3. MATERIALES Y MÉTODOS
3.1. Ubicación del experimento
El predio se encuentra ubicado en la localidad de Mialqui, comuna de Monte Patria,
Provincia del Limarí, IV Región. El predio cuenta con una superficie de 13
hectáreas, en las cuales se encuentran cultivadas en su totalidad con cerezos cultivares
‘Brooks’ y ‘Newtar’.
3.2. Diseño experimental
En la investigación se utilizaron 40 árboles de cerezo, cultivares ‘Brooks’ y
‘Newstar’, las que estaban sobre patrón Santa Lucía 64, de las 13 hectáreas 1
corresponde a ‘Newstar’ y 9,2 de ‘Brooks’, los cuales fueron plantados el año 1999,
con una densidad de 889 plantas por hectárea, con un marco de plantación de 4,5×2,5

m y con un 33,3 % del polinizante ‘Garnet’ y ‘Ruby’, el huerto posee un sistema de
riego por goteo.
A estas plantas se les aplicó Cianamida Hidrogenada mezclado con aceite mineral en
una dosis de 2,5% de producto comercial Cianamida Hidrogenada y 2% de aceite
mineral, la aplicación se realizó con una motobomba de 200 litros, en cuatro fechas
diferentes.
3.3. Acumulación de frío
El modelo que se utilizó para medir la acumulación de frío invernal, es el Modelo
Utah, éste es ideado por RICHARDSON, SEELEY y WALKER (1974) Cuadro 1.
Cuadro 1. Conversión de temperaturas según el modelo de Utha.
TEMPERATURA (ºC) UNIDADES DE FRÍO
< 1,4 0
1,5-2,4 0,5
2,5-9,1 1
9,2-12,4 0,5
12,5-15,9 0
16,0-18,0 -0,5
>18 -1
Fuente: Melgarejo, 1996.
Con el propósito de determinar la acumulación óptima de horas frío para la aplicación
de Cianamida Hidrogenada bajo las condiciones agro climáticas de la IV Región, se
evaluó el efecto de cuatro fechas de aplicación, más el testigo sin aplicación, cada
fecha con una determinada acumulación de frío, calculadas bajo el modelo de Utha, el
de bajo 7,2 °C y el modelo que va de 0 a 7,2 °C, desde el 11 de mayo hasta el 31 de

julio (Cuadro 2), ya que el instrumento pudo ser conseguido en la fecha indicada
anteriormente.
Cuadro 2. Tratamientos con Cianamida Hidrogenada, y la acumulación de frío al momento de la aplicación, bajo distintos modelos de acumulación de frío. La acumulación de frío para el tratamiento sin aplicación fue medida hasta el 31 de Julio.
Frío
acumulado Tratamiento
Fecha de aplicaciónUnidades
Utha 0–7°C <7°C T0 Sin aplicación 810.1 550.1 625.5 T1 05-Jul 532.7 404.7 425.7 T2 13-Jul 547 430.3 506.0 T3 22-Jul 618.8 489.0 564.6 T4 31-Jul 691.7 550.5 625.5
De acuerdo al umbral para la aplicación de Cianamida se satisface con un 50 a 70 %
de la normal acumulación de horas–frío. (GIL, 2000).
Se utilizaron 20 plantas de cada cultivar, conducidas en solaxe, las cuales fueron
seleccionadas por vigor y estado sanitario. Además, se localizó el ensayo en la zona
más homogénea del huerto.
Para analizar los datos de floración se utilizó el método de Stembridge and Gambrell
(1974), con un Diseño Completamente al Azar con Submuestreo y se utilizó el Test
de Tuckey, para realizar la separación de medias. Para el caso de la floración y
brotación se hizo un análisis descriptivo de los diferentes estados fenológicos, que se
indican en el Cuadro 3.
Cuadro 3. Estados fenológicos a medir de los cuales se utilizó el estado lado verde y flores abiertas:
Nº del estado Estado fenológico

1 Yema en reposo 2 Yema Hinchada 3 Lado verde 4 Punta verde 5 Yema abierta o reventada 6 Yemas en botón blanco 7 Flores en capullo 8 Flores abiertas 9 Flores en caída de pétalos
10 Fruto cuajado Fuente: Universidad Estatal de Michigan, 2002
La unidad experimental corresponde a la planta, se utilizaron dos ramas, de dos años
de edad, por árbol, las cuales son las submuestras, estas poseen crecimientos de una y
dos temporadas y se estableció un número de cuatro repeticiones por tratamiento, las
ramas fueron elegidas en función de los puntos cardinales.
La floración y la brotación, se midió dos veces por semana, en la cual se contó el
número de yemas florales y vegetativas en los diferentes estados fenológicos, para
esto se contó el número de yemas de cada rama, tanto vegetativas como florales, las
yemas vegetativas son más delgadas y puntiagudas a diferencia de las florales que son
más anchas y redondas. Las mediciones se realizaron desde julio hasta noviembre.
Además, se llevó un registro desde mayo hasta octubre, de las temperaturas del sector
donde está ubicado el ensayo, cada 10 minutos mediante el uso de un registrador
digital de temperatura y humedad, para determinar las horas frío del sector y la
cantidad de frío acumulado al momento de cada tratamiento, las temperaturas
máximas, mínimas y medias, al igual que la humedad relativa.
4. PRESENTACIÓN Y DISCUSIÓN DE RESULTADOS

4.1. Salida del receso
Corresponde a los primeros estados de desarrollo en yemas florales, en los cuales las
yemas pasan del estado de dormida al estado denominado lado verde, este estado se
extendió desde el 23 de julio al 21 de octubre, como primera respuesta observada
hubo un significativo adelanto del estado, lado verde, en ambos cultivares y en todos
los tratamientos con respecto al testigo sin aplicación, lo cual se puede comparar con
lo observado por LEMUS (1998), donde la primera respuesta observada fue un
adelanto del inicio de la floración en ‘Bing’ y ‘Van’, respecto al Testigo.
En Ovalle aplicaciones semanales de Cianamida Hidrogenada, entre el 12 de junio y
el 24 de julio adelantaron, entre 34 y 7 días el inicio de brotación, en vid cv.
‘Sultanina’ (SEPULVEDA, 1986). Esto se corrobora con los ensayos realizados por
MUÑOZ et al., (1988), donde aplicaciones en vid cv. ‘Sultanina’, con aplicaciones 90
días antes de la brotación normal, se produjo un adelanto de ella, el cual fluctúo entre
7 a 10 días.
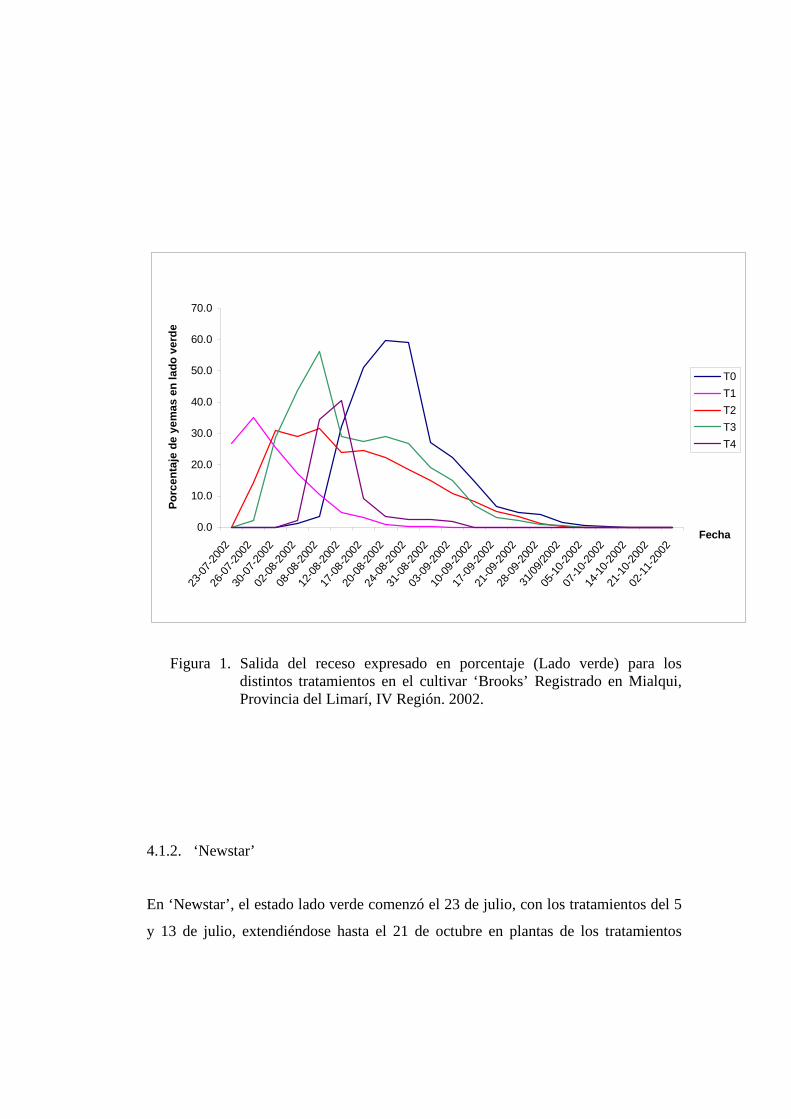
4.1.1. ‘Brooks’
En ‘Brooks’, el estado lado verde comenzó el día 23 de julio, con el tratamiento
aplicado el 05 de julio y terminó el 21 de octubre con plantas del testigo, sin
aplicación, mostrando el tratamiento del 5 de julio un adelanto de 11 días respecto al
inicio del testigo, el cual comenzó el 02 de agosto y se mantuvo hasta el 21 de
octubre (figura 1). En Israel aplicaciones de Cianamida Hidrogenada el 11 de marzo,
con una acumulación de aproximadamente de 350 unidades de frío según modelo de
Utha, produjo un adelanto en la floración, en cv. ‘Sam’, de 18 días comparado con el
testigo (SNIR Y EREZ, 1988). Lo que demuestra que en nuestras condiciones se
observa una tendencia similar.

En la figura 1, se observa que todos los tratamientos con Cianamida Hidrogenada
desplazaron el “pick” del estado lado verde, anticipándolo respecto al testigo.
El testigo presentó un proceso largo, que contrasta con el tratamiento del 5 de julio, el
cual fue concentrado. En vid cv. ‘Perlette’, estacas tratados con Cianamida
Hidrogenada tuvieron un adelanto y una mayor uniformidad en la brotación que las
estacas no tratados (DOKOOZLIAN Y WILLIAMS, 1995), en consecuencia, el
comportamiento en yemas de cerezo sigue un patrón similar a las yemas de vid.
0.0
10.0
20.0
30.0
40.0
50.0
60.0
70.0
23-0
7-20
0226
-07-
2002
30-0
7-20
0202
-08-
2002
08-0
8-20
0212
-08-
2002
17-0
8-20
0220
-08-
2002
24-0
8-20
0231
-08-
2002
03-0
9-20
0210
-09-
2002
17-0
9-20
0221
-09-
2002
28-0
9-20
0231
/09/20
0205
-10-
2002
07-1
0-20
0214
-10-
2002
21-1
0-20
0202
-11-
2002
Fecha
Porc
enta
je d
e ye
mas
en
lado
ver
de
T0T1T2T3T4
Figura 1. Salida del receso expresado en porcentaje (Lado verde) para los distintos tratamientos en el cultivar ‘Brooks’ Registrado en Mialqui, Provincia del Limarí, IV Región. 2002.
4.1.2. ‘Newstar’
En ‘Newstar’, el estado lado verde comenzó el 23 de julio, con los tratamientos del 5
y 13 de julio, extendiéndose hasta el 21 de octubre en plantas de los tratamientos

testigo y tratamiento del 22 de julio, mostrando los tratamientos anteriores un
adelanto de 21 días con respecto al testigo sin aplicación, que comenzó el 12 de
agosto y se mantuvo hasta el 21 de octubre (figura 2).
En arándanos, aplicaciones de Cianamida Hidrogenada a mediados de diciembre y a
principios de enero en Florida Estados Unidos, y a finales de enero en el estado de
Georgia aumentaron la brotación y acortaron el periodo de desarrollo del fruto
(WILLIAMSON, et al., 2002). Por lo tanto, se puede afirmar que las yemas de
cerezo, poseen un comportamiento similar a las yemas de arándanos, sometidas a la
aplicación de Cianamida Hidrogenada, en el sentido que tratamientos invernales
adelantan la cosecha.
La Figura 2, muestra que la respuesta en ‘Newstar’ no es tan marcada como lo que se
observa para ‘Brooks’ (Figura 1). En ‘Newstar’ también se observa un adelanto en el
estado de lado verde de los árboles tratados con Cianamida, pero hay un mayor
traslape de estos eventos, lo cual puede deberse a un mayor requerimiento de frío que
posee ‘Newstar’, respecto de ‘Brooks’ (Figura 2).
Para este cultivar el tratamiento 1 del 5 del julio indujo mayor adelanto, pero el
tratamiento 4 aplicado el 31 de julio, tuvo una mayor concentración del estado lado
verde, lo cual puede deberse a una mayor acumulación de frío al momento de la
aplicación de Cianamida Hidrogenada (cuadro 2), con lo cual se estima para este
cultivar, que aplicaciones de Cianamida Hidrogenada, con una mayor acumulación de
frío, tienen una mejor respuesta, ya que concentraría los diferentes estados
fenológicos, a niveles similares a los de áreas mas frías.

0.0
10.0
20.0
30.0
40.0
50.0
60.0
23-07
-2002
30-07
-2002
06-08
-2002
13-08
-2002
20-08
-2002
27-08
-2002
03-09
-2002
10-09
-2002
17-09
-2002
24-09
-2002
01-10
-2002
08-10
-2002
15-10
-2002
22-10
-2002
29-10
-2002
05-11
-2002
12-11
-2002
Fecha
Porc
enta
je d
e ye
mas
en
lado
ver
de
T0T1T2T3T4
Figura 2. Salida del receso expresado en Porcentaje (Lado verde) para los
distintos tratamientos en el cultivar ‘Newstar’ Registrado en Mialqui, Provincia del Limarí, IV Región. 2002
4.2. Floración
Esté periodo se extendió, para ambos cultivares desde el 08 de agosto hasta el 14 de
octubre, observándose una cierta cronología en la floración de los tratamientos, según

la fecha de aplicación de Cianamida. La floración se adelanta con respecto a la VIII
región, ya que con la aplicación en julio de Cianamida Hidrogenada al 2% en el cv.
‘Van’, el período de floración va desde el 27 de septiembre hasta el 06 de octubre
(JOUBLAN, 1997), existiendo un adelanto de 50 días, no obstante en esta zona la
floración es un poco más prolongada, como se observa en el cuadro 4.
Cuadro 4. Comparación de Fechas de floración y cosecha, entre las zonas de Romeral, provincia de Curico, VII región, 2001 y Mialqui, provincia del Limarí, IV región, 2002, de los cultivares ‘Brooks’ y ‘Newstar’.
Agosto Septiembre Octubre Noviembre
Cultivar 4 8 12 16 20 24 26 30 3 7 11 15 19 23 27 1 5 9 13 17 21 25 29 2 6 10 14 18 22 26 30 3 7 11Brooks Romeral(1)
F F F F F F F C C C C
Brooks IV Región (T1)
F F F F C C C C
Newstar Romeral(1)
F F F F F F F C C C
Newstar IV Región (T2)
F F F F F F F F F F
C C C C C
1) Fuente: JIL, 2002. F: Floración C: Cosecha
Como se observa en la figura 3, las condiciones climáticas desde julio a octubre
presentaron una temperatura con tendencia al alza, con una máxima de 34 °C el día
24 de septiembre y una mínima de -1,5 °C el día 3 de agosto, con una Humedad
Relativa (HR) del período de floración, el cual posee una tendencia a la baja, con una
HR máxima de 100 % el día 13 de agosto y una mínima de un 18 % el 31 de agosto.

-5
0
5
10
15
20
25
30
35
40
04-07
-2002
11-07
-2002
18-07
-2002
25-07
-2002
01-08
-2002
08-08
-2002
15-08
-2002
22-08
-2002
29-08
-2002
05-09
-2002
12-09
-2002
19-09
-2002
26-09
-2002
03-10
-2002
10-10
-2002
17-10
-2002
Fecha
Tem
pera
tura
(°C
)
T. max. T. min. T. media
a)
0
20
40
60
80
100
120
13-0
8-20
0220
-08-
2002
27-0
8-20
0203
-09-
2002
10-0
9-20
0217
-09-
2002
24-0
9-20
0201
-10-
2002
08-1
0-20
0215
-10-
2002
Fecha
Hum
edad
Rel
ativ
a (%
)
H.R. max. H.R. min. H.R. media
b)
Figura 3. a) Temperaturas y b) Humedad Relativa; máximas, mínimas y medias registradas en Mialqui, Provincia del Limarí, IV Región el año 2002.

4.2.1. ‘Brooks’
La floración comenzó el 08 de agosto, con el tratamiento del 5 de julio y
terminó el 28 de septiembre con el tratamiento testigo, como se observa en la
figura 4. El tratamiento aplicado el 05 de julio indujo a una floración más
anticipada y uniforme, la cual se inició el 08 de agosto, con una acumulación
de 278,85° día (base 4,5) (anexo 2), alcanzando el máximo número de flores
el 17 del mismo mes, en Quillota la floración comienza el 13 de septiembre,
con un largo de 19 días (CORTÉS, 2002), en la temporada siguiente se
presentó muy temprano y concentrada, comenzando el 21 de agosto,
alcanzando la plena flor el 27 de agosto, un día después de ‘Burlat’
(ÁLVAREZ, 2003), en la zona de Romeral (Curicó) comenzó el 14 de
septiembre y se extendió hasta el 12 de octubre (JIL, 2002), con una
acumulación de frío de 901 horas bajo 7°C, acumulación suficiente para este
cultivar, por lo que no se aplica Cianamida Hidrogenada. En la región de
Bordeaux, Francia, la floración es similar a la de Quillota (2002), cero a dos
días antes que ‘Burlat’ (CLAVERIE, 2002). En Zaragoza, España, la época
de floración va entre el 29 de Marzo y el 2 de Abril, siendo mas tarde y
concentrada que en Quillota (GARCÍA y FAÑANAS, 2002).
Las aplicaciones posteriores indujeron floración en el mismo orden que fueron
realizadas, alcanzando el máximo de flores abiertas entre el 17 de agosto y el 11 de
fueron similares en la fecha en que alcanzaron el máximo de flores abiertas.

0
50
100
150
200
250
300
23-07
-2002
26-07
-2002
30-07
-2002
02-08
-2002
08-08
-2002
12-08
-2002
17-08
-2002
20-08
-2002
24-08
-2002
31-08
-2002
03-09
-2002
10-09
-2002
17-09
-2002
21-09
-2002
28-09
-2002
31/09
/2002
05-10
-2002
07-10
-2002
14-10
-2002
21-10
-2002
02-11
-2002
Fecha
N° d
e flo
res
abie
rtas T0T1T2T3T4
Figura 4: Número de flores abiertas en el cv. ‘Brooks’, en la localidad de Mialqui,
Provincia del Limarí, IV Región, 2002.

En el cuadro 5 se presenta un resumen de la floración en ‘Brooks’.
Cuadro 5. Periodo de floración para los distintos tratamientos cv. ‘Brooks’, en la localidad de Mialqui, Provincia del Limarí, IV Región el año 2002.
Agosto Septiembre Tratamiento 2 8 12 17 20 24 28 1 3 7 11 15 19 21 25
Sin Aplicación PF
T1 PF
T2 PF
T3 PF
T4 PF
PF: indica la fecha en que se presentó el máximo número de flores abiertas.
Como se observa en el cuadro 5, la duración de la floración varió entre la aplicación
del 05 de julio y las posteriores, siendo de dos semanas la del primero y de más de
tres para los tratamientos posteriores. Uno de los objetivos del manejo del cerezo es
tener una floración concentrada y coincidente con los cultivares polinizantes, para
que la cuaja sea efectiva, al existir alta floración en poco tiempo y la cosecha resulte
más fácil, ya que la fruta presenta un similar estado de madurez, en un periodo más
corto.
En el cuadro 6, se muestra la diferencia estadística de la floración entre los
tratamientos.

Cuadro 6. Diferencia de la floración entre los tratamientos cv. ‘Brooks’,según método de Stembridge and Gambrell y Test de Tuckey con un 5% de significancia.
Tratamientos Días de adelanto
T0 0e T1 25.8a T2 19.2c T3 11.8d T4 21.8b
4.2.2. ‘Newstar’
La floración se extendió desde el 17 de agosto, con el tratamiento aplicado el 13 de
julio, hasta el 14 de octubre con el tratamiento testigo, como se observa en la figura 5,
la aplicación del 13 de julio, tuvo una floración más anticipada y de mayor magnitud
que las otras, a diferencia de lo ocurrido en ‘Brooks’, iniciándose la floración el 17 de
agosto, con una acumulación de 380,05° día (base 4,5) (anexo 3), la plena flor fue el
24 del mismo mes y se extendió hasta el 21 de septiembre. La cantidad de flores
también difiere respecto a ‘Brooks’, siendo mucho menor. En Quillota durante la
temporada del 2001, la floración de este cultivar comenzó el día 15 de septiembre,
dos días después que ‘Burlat’, y se extendió hasta el 2 de octubre, en la temporada
siguiente ‘Newstar’ comenzó la floración el 30 de agosto, seis días después de
‘Burlat’, y la plena flor se alcanzó el 11 de septiembre, 16 días después de ‘Burlat’
(ÁLVAREZ, 2003), en la zona de Romeral, floreció más tarde, el 22 de septiembre,
con una duración de 24 días, presentando un máximo número de flores abiertas el 6
de octubre (JIL, 2002). En la región de Bordeaux, Francia, es semitardía, de -1 a +2
días respecto a ‘Burlat’ (CLAVERIE, 2002). En Zaragoza, España, es considerada de
floración temprana, comienza el 27 de marzo y termina el 31 de marzo (GARCÍA y
FAÑANAS, 2002).

Con respecto a la salida del receso, la floración tuvo una marcada diferencia entre los
tratamientos, como se puede observar en la figura 5, sobre todo entre el tratamiento
aplicado el 13 de julio, siendo esta anticipada y uniforme, por el contrario el testigo
sin aplicación tuvo una floración tardía y prolongada, los tratamientos aplicados el
22 de julio y el 31 de julio fueron similares en cuanto ha adelanto y duración de la
floración.

0
5
10
15
20
25
30
23-07
-2002
30-07
-2002
06-08
-2002
13-08
-2002
20-08
-2002
27-08
-2002
03-09
-2002
10-09
-2002
17-09
-2002
24-09
-2002
01-10
-2002
08-10
-2002
15-10
-2002
22-10
-2002
29-10
-2002
05-11
-2002
12-11
-2002
Fecha
N° d
e flo
res
abie
rtas
T0T1T2T3T4
Figura 5. Número de flores abiertas en el cv. ‘Newstar’, en la localidad de
Mialqui, Provincia del Limarí, IV Región, 2002.

Las plantas con el tratamiento aplicado el 05 de julio iniciaron la floración el 20 de
agosto, siendo ésta, prolongada, incluso más que el tratamiento testigo que comenzó
su floración el 07 de septiembre, lo siguiente se resume en el Cuadro 7.
Cuadro 7. Periodo de floración para los distintos tratamientos cv. ‘Newstar’, en la localidad de Mialqui, Provincia del Limarí, IV Región el año 2002.
Agosto Septiembre Octubre Tratamiento 12 16 20 24 28 1 3 7 11 17 21 25 28 2 6 10 14 18
Sin Aplicación PF
T1 PF
T2 PF
T3 PF
T4 PF
PF: indica la fecha el la que se presentó el máximo número de flores abiertas.
En el cuadro 7, se observa un adelanto en la floración del tratamiento aplicado el 13
de julio frente a la aplicación del 5 de julio, a diferencia de lo afirmado por LYON,
POLLER, y RODRIGUEZ y OPAZO (1994), en la cual indican que el adelanto de la
floración es mayor en la medida que se realizan aplicaciones de Cianamida
Hidrogenada más temprano, pues a medida que se acumule mayor cantidad de frío el
tratamiento es menos efectivo.
Cuadro 8 muestra la diferencia estadística entre la floración de los tratamientos.

En este cultivo se observa una más larga floración para todos los tratamientos,
respecto al cv. ‘Brooks’.
Cuadro 8. Diferencia de la floración entre los tratamientos cv. ‘Newstar’,según método de Stembridge and Gambrell y Test de Tuckey con un 5% de significancia.
Tratamiento Días de adelanto
Sin aplicación 0e
T1 27.7b
T2 29.5a
T3 15.9d
T4 17.1c
Según LEMUS (1997), en Buin se produjo un adelanto de ocho días en ‘Bing’ y de
once días en ‘Van’, tratados con un 2% de Cianamida Hidrogenada, lo que permitió,
el 03 de octubre, un estado de abundante floración en ambos cultivares.
El momento de la aplicación es fundamental, ya que aplicaciones demasiado
tempranas no tienen efecto, mientras que las demasiado tardías pueden atrasar en vez
de adelantar la floración, por otra parte dañar la yema floral (LEMUS 1997), según
SNIR Y EREZ (1988) si la Cianamida es aplicada muy tarde, podría ser fitotoxico
para los brotes.
Al respecto VALENZUELA (1998), afirma que en cerezos, se desbebiese aplicar
entre mediados de julio y principios de agosto, para adelantar y uniformar la
floración.
Para vides, cianamida resulta superior a todas otras sustancias químicas que rompen
el receso. Con árboles frutales de hoja caduca, los efectos positivos se observaron en
la mayor parte de las especies, especialmente con cerezos dulces, pero este efecto, en

condiciones de inviernos calidos fueron obtenidos también con manzanos, ciruelos, y
perales (EREZ, 1987).
El empleo de sustancias químicas que rompen el receso, ha permitido el crecimiento
de árboles frutales de hoja caduca en áreas más tropicales donde nunca había antes
sido posible. El interés mundial en cultivar las especies caducas ha conducido a varias
formas de crecimiento de los árboles, sobre todo, la técnica de anulación de receso en
los trópicos. La perspectiva para una remota extensión en el crecimiento de árboles
frutales de hoja caduca es brillante (EREZ, 1987).
La deficiencia de frío, conduce a tres efectos que varían dependiendo del nivel de
deficiencia: a) Pobre brotación, pobre desarrollo del follaje, flor escasa y con
frecuencia flores anormales. b) Foliación retrasada y floración, y brotación desigual.
c) Pobre cantidad de frutas, el área de la hoja reducida, y el temprano cese de
crecimiento, debido a la inactividad secundaria. El desarrollo de fruta desigual,
producto de una floración desigual (EREZ, 1987).
Con respecto a la falta de fruta, puede atribuirse al déficit de frío invernal que puede
producir fenómenos como aborto del estilo, pistilos múltiples, lo que ocasionan frutos
múltiples, alteraciones en el desarrollo del polen y yemas que mueren antes de
desarrollarse el brote (MELGAREJO, 1996), o bien puede estar relacionado con un
problema de humedad relativa.
Salto de página
5. CONCLUSIONES
En el cv. ‘Brooks’ el tratamiento de Cianamida Hidrogenada, aplicado el 05 de julio
(532.7 unidades frío bajo el modelo de Utha), fue el que indujo un mayor adelanto y
concentración de la floración.

En el cv. ‘Newstar’ el tratamiento de Cianamida Hidrogenada, aplicado el 05 de julio
(532.7 unidades frío bajo el modelo de Utha), junto con el tratamiento del 13 de julio
(547.0 unidades frío bajo el modelo de Utha), alcanzaron una mayor precocidad de la
floración, que el resto de los tratamientos.
Cianamida adelanta la floración, en comparación a no aplicar, entre 16 y 20 días
respecto al testigo sin aplicación, así como también la concentra entre dos a tres
semanas, según el cultivar y la acumulación de frío al momento de la aplicación.
Salto de página
6. RESUMEN
Recientemente, se han realizado plantaciones de cerezo dulce en el norte de Chile, lo que se ha producido gracias a la incorporación de cultivares con una baja acumulación de frío y al empleo de Cianamida Hidrogenada, compensador del frío invernal. El objetivo de este trabajo fue determinar el momento óptimo de aplicación de Cianamida Hidrogenada en cerezos, bajo las condiciones agroclimáticas de la IV Región, para lo cual se evaluaron cuatro fechas de aplicaciones, en árboles de cerezo de tres años, en la localidad de Mialqui, comuna de Monte Patria, donde se registraron, hasta el 31 de Julio, 691,7 horas frío. La floración más precoz y concentrada en ‘Brooks’, se obtuvo con el tratamiento aplicado el 05 de julio, con una acumulación de 532,7 horas frío, que fue el que indujo una mayor precocidad y concentración tanto en la floración como en la brotación, la cual comenzó con una acumulación de 278,85 ° días (base 4,5). En ‘Newstar’ la floración más precoz se logró con el tratamiento aplicado 13 de julio, que tuvo una acumulación de 547,0 horas frío, la cual comenzó con una acumulación de 380,05° día (base 4,5). No obstante la floración más concentrada se logró con los tratamientos del 22 y 31 de julio, que no fueron precoces en su floración, pero presentaron una mayor concentración, cuyas floraciones comenzaron con una acumulación de 540,05 ° día (base 4.5) cada una, y con una duración de la floración de 14 días. Cianamida se muestra como una herramienta indispensable en el manejo del cerezo de la IV región.
Salto de página
7. ABSTRACT

Recently, Sweet Cherry (Prunus avium L.) plantings have been made in the North of Chile, which was made possible by the incorporation of cultivars with low chilling requirements and the use of hydrogenated cyanimide, a compensator for winter chilling-hour accumulation. The aim of this study was to determine the optimum time for the application of hydrogenated cyanimide to Sweet Cherries, under the climatic conditions of the IV Region, and for this, four applications were evaluated, in 3 years-old cherry trees, in the village of Mialqui, Monte Patria, where 691.7 chilling-hours were recorded by the 31st of July. More precocious and concentrated blooming for the cultivar ‘Brooks’ was obtained with the 5th of July treatment, with an accumulation of 523.7 chilling-hours, that induced a higher precocity and concentration as much on flowering as on and bud-break, which began with an accumulation of 278.85 growing º days (base 4,5). In the cultivar ‘Newstar’, precocious blooming was achieved with the july 13th treatment, at an accumulation of 547.0 chilling-hours, which began with the accumulation of 380.05 growing º days (base 4.5). Nevertheless, the most concentrated blooming was achieved with the July 22nd and 31st treatments, that did not show early blooming, but did show a higher concentration, with blooming beginning at an accumulation of 540.05 growing º days (base 4.5) for each one, and with flowering lasting for a period of 14 days. Hydrogenated cyanimide was shown to be an indispensable tool in the management of the Sweet Cherry in the IV region.
Salto de página
8. LITERATURA CITADA
ABBOTT, D. 1969. The role of budcales in the morfhogenesis and dormancy of the apple fruit bud. In: Luckwill, L. and Cutting, C. eds. Physiology of the Tree Crops. New york, Acadenic Press. 65-82 pp.
ADKINS, S. W. AND J. ROSS. 1981. Studies in wild oat seed dormancy: The
role of ethylene in dormancy breaking and germinat oat seed (Avena fatua). Plant Physiol. 67:358-362.
ÁLVAREZ, S. 2003.Caracterización fenológica y productiva de 14 cultivares de cerezo dulce en la zona de Quillota, V región. Taller de Licenciatura Ing. Agr. Quillota, Pontificia Universidad Católica de Valparaíso. Facultad de Agronomía. 118p.
AMBERGER, A. 1984. Uptake and metabolism of Hydrogen Cyanamide in plants, IN: R. J. Weaver eds., Proceeding of bud dormancy of grapevines. Potencial

and practical uses of hydrogen cyanamide on grapevines. Davis Univ. of California. 89 pp.
AMEN, R. 1968. A model of seed dormancy. Bot. Rev. 34: 1-31. CHANDLER, W. AND BROWN, D. 1953. Deciduos orchards in California winters.
Univ. calif. Agric. Ext. Ser. Circ. 179. EE.UU. CLAVERIE, J. 2001. Condiciones del cultivo del cerezo en Francia. Seminario
Internacional de Cerezas, Actualización commercial, variedad, portaingerto y sistemas de conducción de cerezas. Corporación Pomanova, Curicó, Chile, 9 y 10 de agosto del 2001. s/p.
-------------------. 2002. Mejoramiento genético varietal del cerezo en Francia
(Prunus avium). Seminario Cultivo del Cerezo en la Zona Centro Norte de Chile. Universidad Católica de Valparaíso. Facultad de Agronomía, Quillota, 27 y 28 de noviembre del 2002. 69-83 pp.
CORTÉS, A. 2002. Comportamiento fenológico y productivo del cerezo dulce
(Prunus avium. L.) en la localidad de la palma, Quillota, V región. Taller de Licenciatura Ing. Agr. Quillota, Universidad Católica de Valparaiso. Facultad de Agonomía. 97 pp.
DENNIS, F. 1994. Dormancy - What we know (and don't know). HortScience 29:
1249-1255. DESARROLLO COMERCIO FRUTICOLA. 2001. Proyección de la producción de
cerezas en Chile y análisis de mercado, (on line). http//:www.fruitonline.com DOKOOZLIAN, N. AND WILLIAMS, L. 1995. Chillikbng exposure and hydrogen
cyanamide interact in breaking dormancy of grape buds. HortScience 30(6): 1244-1247.

EREZ, A. 1987. Chemical Control of Budbreak. HortScience, December Vol.
22(6). 1240-1243. FELKER, K. C. AND H. A. ROBITAILLE. 1985. Chilling accumulation and rest
of sour cherry buds. Amer. Soc. Hort. Sci. 110: 227-232.
FOOT, J. 1987. Effect of Hydrogen Cyanamide on bud emergency in wine grapes. California Agriculture. 41 (3-4): 19.
GARCÍA, R. y FAÑANAS, G. 2002. Situación actual de las variedades de cerezo.
Fruticultura profesional 130: 5-14. GARDIAZABAL, F. 1988. Uso de dormex en kiwi. Producción de kiwi. Curso
breve. Santiago Universidad Católica de Chile. Facultad de Agronomía. p. 305-312.
GIL, G. 2000. El potencial productivo, Crecimiento vegetativo y diseño de huertos y viñedos. Santiago. Ed. Pontificia Universidad Católica de Chile. 583p.
GORDON, E. 1987. Producing Temperate-Zone Fruit At Low Latitudes: Avoiding
Rest and the Chilling Requirement. HortScience, Vol. 22(6): 1236-1240. JIL, M. 2002. Caracterización de nuevos cultivares de cerezo dulce (Prunus avium
L.) en la zona de Romeral, Provincia de Curicó, VII Región. Taller de licenciatura Ing. Agr. Universidad Católica de Valparaíso, Facultad de Agronomía. Quillota. 74 p.
JOUBLAN, J., LEMUS, G. 1997. Manejo del vigor y respuesta a reguladores de
crecimiento en la Octava región. Seminario Internacional actualidad mundial en el cultivo del cerezo. Universidad de Concepción, Chillan, 16 y 17 octubre del 1997. 92-118 p.

LEMUS, G. 1998. Manejo del cerezo. Efecto de algunos reguladores de crecimiento. 1. Huertos de alta densidad. Tierra Adentro. N° 18: 10-12.
LEMUS, G. 2002. Cianamida Hidrogenada como compensador de frío invernal.
Tierra adentro. Nº 44: 20-22. LEMUS, G. y RIVAS B. 2002. El cultivo del cerezo en zonas mediterráneas y
semiáridas: acercamiento a la experiencia y tecnología europea. Informe de Gira Tecnológica Centro Regional de Investigación INIA La Platina - FDI. Santiago Chile. 43 p.
LYON, M. POLLER, S. y RODRIGUEZ-CANO, M. 1989. Evolución del letargo de
yemas en seis especies frutales en relación al frío y tratamiento con tiourea o Cianamida. Tesis Ing. Agr. Santiago Universidad Católica de Chile, Facultad de Agronomía. 82 pp.
MELGAREJO, P. 1996. El frío invernal, factor limitante para el cultivo frutal, Modelos y métodos para determinar la acumulación de frío y de calor en frutales. Ed. A. Madrid Vicente.
MUÑOZ, I. VALENZUELA, J. GALVEZ, S. PEZOA, J. 1988. Uso de Cianamida
Hidrogenada en ‘Sultanina’. IPA La Platina N° 48: 26-31. NIGOND, J. 1967b. Recherches sur la dormance des bourgeons de la vigne. III.
Röle de la température dans I´évolution de la dormance. Ann Physiol. Vég. 9: 273-338.
OPAZO, I. 1994. Efecto de la aplicación de Cianamida Hidrogenada en mezclas con
aceite mineral, en cerezos (Prunus avium) cv. ‘Van’. Tesis Ing. Agr. Santiago Universidad de Chile, Facultad de Ciencias Agrarias y Forestales. 46 pp.
RAZETO, B. 1999. Para entender la fruticultura. 3a edición. Santiago, Vertigo. 373 p.

RICHARDSON, E.,SEELEY, S. AND WALKER, R. 1974. A Model for Estimating Completion of Rest for ‘Redhaven’ and ‘Elberta’ Peach Trees. HortScience 9: 331-332.
ROMBERGER, J.A. 1963.Meristem, growth, and development in woody plants.
U.S.D.A. For. Ser. Tech. Bull. 1293 EE.UU. SAUNIER, R. 1997. Portainjertos y variedades de cerezos, actualidad y tendencias en
Francia. Seminario Internacional Actualidad Mundial en el Cultivo del Cerezo. Universidad de Concepción, Chillan, 16 y17 de octubre del 1997. 9-34 p.
SCHNABEL, B. AND WAMPLE, R. 1987. Dormancy and cold hardiness in Vitis
vinifera L. Cv. White Riesling as influenced by photoperiod and temperature. Am. J. Enol. Vitic. 38 (4): 265-272.
SEPULVEDA, G. 1986. Uso de Cianamida Hidrogenada en vid cv. ‘Sultanina’, en
tres localidades del Norte Chico. Santiago Pontificia Universidad Católica de Chile. 160 p.
SEPULVEDA, G. 1987. Uso de Cianamida Hidrogenada en Vid, cvs ‘Sultanina’,
Perlette y Flame Seedless en tres localidades del Norte Chico. Informe Técnico centro Regional de Investigación La Platina. Santiago. 143 p.
SEPULVEDA, G. 1989. Efecto de dos reguladores de crecimiento sobre la
interrupción de la dormancia en cv. ‘Sultanina’. Informe Técnico Centro Regional de Investigación La Platina. Santiago Chile. 130 p.
SOTOMAYOR, C. 1995a. EL Cerezo. Chile agrícola.Vol. 21: 45-47. SOTOMAYOR, C. 1995b. El Cerezo. Chile agrícola. Santiago. Vol. 21: 90-92.

SHULMAN, Y., NIR, G., LAVEE, S., FANBERSTEIN, L. 1983. The effect of Cyanamide on the release from dormancy of grapevine buds. Scientia Horticultural. 19: 97 – 104.
SNIR I. AND EREZ A. 1988. Bloom advancemt in sweet cherry by hydrogen
cyanamide. Fruit varieties journal, 42(4): 120-122. STEMBRIDGE, G. E., GAMBRELL, C. E. 1974. Measuring peach thinning and
maturation responses. Hortscience, vol. 9(1): 29-30 SWK-Trostberg. 2002. (on line). http://www.dormex.com/cherries1.htm TIMMIS, K. A. L. H. FUCHIGAMI AND R. TIMMIS. 1981. Measuring
dormancy: The rise and fall of square waves. Hortscience. 16: 200-202 TINKLIN, I AND SCHWABE, W. 1970. Lateral bud dormancy in the black currant
Ribes nigrum (L.). Ann. Bot. 34: 691-706. University Michigan State. 2002. Fruit Growth Stage Reference - Tart Cherry, (on
line). www.msue.msu.edu/fruit/tchrygrw.htm. VALENZUELA, L. 1998. El cultivo del cerezo; aspectos técnicos. Revista Frutícola.
V. 19(2): 55-68.
VALENZUELA, J. y LOBATO, A. 2000. Uva de mesa en Chile. Santiago INIA. 338 p.
WESTWOOD, M. N. AND H. O. BJORNASTAD. 1978. Winter rainfall reduces period of apples and pears. J. Amer. Soc. Sci. 103: 142-144.
WILLIAMSON J., KREWER G., MAUST B. AND MILLER E. 2002. Hydrogen
cyanamide accelerates vegetative budbrake and shortens fruit development periodic of blueberry. HortSience 37(3): 539-542.


ANEXOS