01 anatomia renal
DESCRIPTION
Capitulo 01 traducido del Comprehensive Clinical NephrologyTRANSCRIPT

La compleja estructura del riñón de mamífero se entiende mejor en la
forma unipapillary que es común a todas las especies pequeñas. La
figura 1-1 es una sección esquemática coronal a través de un riñón
unipapillary, con una corteza que encierra una médula en forma de
pirámide, la punta del que sobresale en la pelvis renal. La médula se
divide en una externa y una médula interna; la médula externa se
subdivide en una externa y una franja interior.
Los componentes específicos del riñón son las nefronas, los conductos
colectores, y un único microvasculature. El riñón multipapillary de los seres
humanos contiene aproximadamente 1 millón de nefronas, aunque este
número varía considerablemente. El número de nefronas ya se ha establecido
durante el desarrollo prenatal; después del nacimiento, las nuevas nefronas
no se pueden desarrollar, y una nefrona perdido no puede ser reemplazada.
A nefrona consta de un corpúsculo renal (glomérulo) conectado a un
túbulo complicado y retorcido que finalmente desemboca en un conducto
colector (Fig. 1-2 y Tabla 1-1). Tres tipos de nefrona se pueden distinguir
por la ubicación de corpúsculo s renales dentro de la corteza: superficial,
cortical media, y nefronas yuxtamedulares. La parte tubular de la nefrona
consta de un túbulo proximal y un túbulo distal conectado por un asa de
Henle (véase la discusión más adelante). Hay dos tipos de nefrona,
aquellos con largas asas de Henle y los que tienen bucles cortos. Bucles
cortos se convierten de nuevo en la médula externa o incluso en la
corteza (bucle cortical). Bucles largo s retroceden a niveles sucesivos de
la médula interna.
Un conducto colector se forma en la corteza renal cuando varias
nefronas se unen. Un túbulo de conexión está interpuesto entre una
nefrona y un conducto colector cortical . Conductos colectores
corticales descienden dentro de los radios medulares de la corteza.
Estos conductos atraviesan la médula externa como tubos
ramificados . Al entrar en la médula interna, los conductos colectores
corticales fusionan sucesivamente y abierto, finalmente, como
conductos papilares en la pelvis renal (ver Fig. 1-2 y la Tabla 1-1).
El patrón microvascular del riñón está organizado de manera similar en
species1,3 mamíferos (Fig 1-3;. Véase también la figura 01.01.). La
arteria renal, después de introducir el seno renal, finalmente divide en
las arterias interlobulares, que se extienden hacia la corteza en el
espacio entre la pared de la pelvis (o cáliz) y el tejido cortical adyacente.
En el cruce entre la corteza y la médula, las arterias interlobulares dividen y
pasan por encima en las arterias arqueadas, que también se ramifican. Las
arterias arqueadas dan lugar a las arterias radiales corticales (arterias
interlobulares), que ascienden radialmente a través de la corteza. No hay
arterias penetratethe médula. Arteriolas aferentes suministran los
mechones glomerulares y generalmente surgen de las arterias radiales
corticales. Afluentes Aglomerular a los plexos capilares rara vez se
encuentran. Como resultado, el suministro de sangre de los capilares
peritubulares de la corteza y la médula es exclusivamente postglomerular.
Los glomérulos son drenados por arteriolas eferentes. Dos tipos básicos de las
arteriolas eferentes se pueden distinguir, cortical y yuxtamedulares. Arteriolas
eferentes corticales, que derivan de glomérulos superficiales y midcortical,
suministran el plexo capilar de la corteza. Las arteriolas eferentes de los
glomérulos yuxtamedulares representan los vasos que irrigan de la médula renal.
Dentro de la franja exterior de la médula, estos vasos se dividen en la recta
descendente vasa y después penetran en el interior de la raya en los haces
vasculares en forma de cono. A intervalos, los vasos individuales dejan los
manojos para suministrar el plexo capilar a nivel medular adyacente.
Ascendente vasos rectos drenar la médula renal. En la médula interna, surgen
los vasos rectos en todos los niveles, ascendiendo vasos como no ramificados, y
atravesar la franja interior dentro de los haces vasculares. La recta ascendente
vasa que drenan la franja interior puede unirse a los haces vascula res o puede
ascender directamente a la franja exterior entre los haces. Todos los vasos
rectos ascendente atravesar la franja exterior como buques onduladas
individuales con amplia lumina intercalados entre los túbulos. Porque los
verdaderos capilares derivadas de ramas directas de las arteriolas eferentes son
relativamente escasos, forman los vasos rectos ascendente plexo capilar de la
franja exterior. El vasos rectos ascendentedesembocanenvenasarqueadas.
Los haces vasculares representan un intercambiador en contracorriente entre la
sangre que entra y que dejando la médula. Además, la organización de los haces
vasculares en los resultados de una separación del flujo de sangre a la tira interior
de aquél al los vasa recta medulla.Descending interior el suministro de la médula
interna atravesar la franja interior dentro de los haces vasculares. Por lo tanto la
sangre que fluye a la médula interna no ha sido expuesto previamente a los
túbulos de la franja interior o exterior. Todo ascendente vasos rectos procedentes
de la médula interna atravesar la franja interior dentro de los haces vasculares.
Por lo tanto la sangre que se ha perfundido túbulos de la médula interna no
posteriormente perfundir túbulos de la franja interior. Sin embargo, la sangre que
vuelve ya sea de la médula interna o la franja interior después hace perfundir los
túbulos de la franja exterior. Esta disposición en la banda exterior puede funcionar
como la trampa definitiva para evitar la pérdida de soluto a partir de la médula.
Las venas intrarrenales acompañan a las arterias. Central hasta el drenaje renal del
riñón son las venas arqueadas, las cuales, a diferencia de las arterias arqueadas, se
forman los arcos reales anastomosados en la frontera corticomedular. Las venas
arqueadas aceptan las venas de la corteza y la médula renal. Las venas arqueadas
unen para formar interlobar venas, que se ejecutan junto a las arterias
correspondientes.

1. Corpúsculo renal 2. túbulo contorneado prox imal
3. proximal túbulo recto 4. Descendente extremid ad delgad a
5. ascendente extremid ad delgad a 6. distal del túbulo rec to (gru esa rama
ascendente)7. Mácula dens a8. túbulo contorneado distal9. túbulo Conexión
10. cortical conducto colecto r11. Outer conducto colector medular
12. Interio r delconductocolectormedular
Figura 1-2 nefronas y el sistema de conductos colectores. Se muestra se
corta en bucle y nefronas-largos bucles, junto con un conducto colector
(no dibujadas a escala). Las flechasindicanconfluencia de másnefronas
Las arterias intrarrenales y de las arteriolas aferentes y eferentes
glomerulares están acompañados por fibras nerviosas simpáticas y
axones terminales que representan los nervios eferentes del riñón.
Túbulos tienen contacto directo con los axones terminales sólo
cuando los túbulos se encuentran al rededor de las arterias o las
arteriolas. Inervación tubular consiste en fibras ocasionales
adyacentes a túbulos perivasculares . La densidad de los contactos de
los nervios a los túbulos contorneados proximales es baja ; nunca se
han encontrado contactos a los túbulos proximales rectas, gruesa
ascendente extremidades de Henle bucles y conductos colectores
(ubicadas en rayos medulares y médula externa). La gran mayoría de
porciones tubulares no tienen relaciones di re ctas a las terminales
nerviosas. Nerviosaferentesdelriñón se cree que son escasos.
El glomérulo comprende un penacho de capilares especializados adjuntos
al mesangio, ambos de los cuales están encerrados en una extensión de
bolsa del túbulo, la cápsula glomerular, o cápsula de Bowman (Figs.1 -4 y 1 -
5). Los capilares y mesangio están cubiertos por las células epiteliales
(podocitos), formando el epitelio visceral de Bowman cápsula. En el polo
vascular, esta estructura se refleja para convertirse en el epitelio parietal
de la cápsula de Bowman. En la interfaz entre los capilares glomerulares y
el mesangio en un lado y la capa de podocitos en el otro lado, la
membrana basal glomerular (GBM) se desarrolla. El espacio entre las dos
capas de Bowman cápsula representa el espacio urinario, el cual en el polo
urinario continúa como la luz del túbulo.
Al entrar en el penacho, la arteriola glomerular aferente divide
inmediatamente en hasta cinco ramas capilares primarios, cada uno de lo
que da lugar a una red capilar anastomosis que representa un lóbulo
glomerular.
Glomérulo : término que se ut iliza con mayor frecu encia p ara
referirse a todo corpúsculo renal Bowman cápsula
parte enrevesada
Parte recta (pars recta) o rama d escend ente gru esa del asa d e Henle
Parte descendente, o rama descendente delgada del asa de Henle Parte ascendente o rama ascendente delgada del asa de Henle Parte recta, o gruesa rama ascendente del asa de Henle: subdividido en medular y partes corticales; la parte cortical contiene la mácula densa en su porción terminal Parte enrevesada
Túbulo colector cortical
Exterior del conducto colector medular: subdividido en una franja externa y una porción de banda interior Interior conducto colector medula r: subdividido en basal, media y porciones papilares
Figura 1-1 Sección coronal a través de un riñón unipapillary.

En contraste con la arteriola aferente, la arteriola eferente glomerular ya
está establecida dentro del mechón por confluencia de los capilares de
cada lobule.6 Así, la arteriola eferente tiene un segmento intraglomerular
significativa situado dentro del tallo glomerular.
Capilares glomerulares son un tipo único de vaso sanguíneo que consta de
sólo un tubo endotelial (Figs. 1-6, 1-7). Una pequeña raya del aspecto
exterior de este tubo hace tope directamente el mesangio; una parte
importante se abomba hacia el espacio urinario y está cubierto por la
MBG y la capa de podocitos. Esta porción periférica de la pared capilar es
el área de filtración. El mesangio glomerular representa el eje de un
lóbulo glomerular al que están unidos los capilares glomerulares.
Membrana basal glomerul ar
El GBM sirve como el esqueleto del penacho glomerular. Esta membrana
es un saco plegado complejamente con una abertura en el hilio glomerular
(véase Fig. 1-4). El aspecto exterior de este saco GBM está completamente
cubierta de podocitos. El interior de la bolsa se llena con los capilares y el
mesangio.
Como resultado, en su aspecto interior, el GBM está en contacto con
cualquiera de los capilares o el mesangio. En cualquier transición entre
estas dos localidades, los cambios GBM de un pericapilar convexa a un
curso perimesangial cóncava; los puntos de inflexión se llaman ángulos
mesangiales.
En las micrografías electrónicas de tejido tradicionalmente fija, la MBG
aparece como una estructura trilaminar, con una lámina densa delimitada
por dos capas menos densas, la externa lámina rara interna y la lámina rara
(véase Fig. 1-7). Los estudios con técnicas de congelación revelan sólo una
capa densa de espesor directamente unido a las bases del epitelio y
endothelium.
Los principales componentes de la MBG incluyen el colágeno de tipo IV,
laminina, y proteoglicanos heparán sulfato, como en las membranas
basales en otros sitios. Tipos V y VI de colágeno y nidogen (entactina)
también se han demostrado. Sin embargo, la MBG tiene varias
propiedades únicas, especialmente un espectro distinto de colágeno de
tipo IV y lamininaisoformas. El GBM madura consta de colágeno de tipo IV
hecha de α3, α4, y las cadenas a5 (en lugar de a1 y a2 cadenas de la
mayoría de las otras membranas basales) y laminina 11, hechas de α5, β2,
y el colágeno γ1 Tipo IV es la chains.8 diana antigénica en la enfermedad
de Goodpasture (véase el capítulo 24), y las mutaciones en los genes de la
α3, α4, y a5 cadenas de colágeno tipo IV son responsables de síndrome de
Alport (véase el Capítulo 48).
Figura 1-3 microvasculatura del riñón. Arteriolas aferentes suministran los
glomérulos y arteriolas eferentes salen los glomérulos y se dividen en los vasos
rectos descendente, que junto con la recta ascendente vasa forman los haces
vasculares de la médula renal. La vasa recta ascendente de la médula interna
toda atravesar la franja interior dentro de los haces vasculares, mientras que la
mayoría de los vasos rectos desde la línea interna del ascenso médula externa
fuera de los paquetes. Ambos tipos atraviesan la franja exterior como, canales
tortuosos de ancho.
Figura 1.4 corpúsculo renal y aparato yuxtaglomerular. (Modificado con
permiso de referencia 1.)
AAarteriola aferenteMDmácula densaEGMmesangioextraglomerular
EAarteriola eferente NTerminales nerviosas simpáticas
GCcélulas granularesSMCCélulas musculares lisas vasculares
PEepitelio parietalPOpodocitosMmesangioE endotelio
Fprocesos de piesGBMLa membrana basal glomerular US espacio urinario

Los modelos actuales representan la estructura básica de la MBG como
una red tridimensional de tipo IV collagen.7 El monómero de coláge no
tipo IV consta de una triple hélice que es 400 nm de longitud, con un
dominio grande, no colágena globular en su extremo C-terminal llamado
NC1. En el terminal N, la hélice posee un triple varilla helicoidal 60 nm de
largo: el dominio 7S. Las interacciones entre los dominios 7S de dos
hélices triples o los dominios NC1 de cuatro hélices triples permiten Tipo
monómeros de colágeno IV para formar dímeros y tetrámeros. Además,
el triple filamentos helicoidales de interconexión por asociaciones
laterales través de la unión de los dominios NC1 a los sitios a lo largo de la
región colagenosa. Esta red se complementa con una red interconectada
de laminina 11, lo que resulta en un conjunto de poligonal flexibles, no
fibrilar que proporciona resistencia mecánica a la membrana basal y sirve
como un andamio para la alineación de otros componentes de la matriz.
Figura 1.5 Sección longitudinal a t ravés de un glomérulo (rata) . En el polo vascular, la arteriola
aferente (AA), la arteriola ef erente (EA) , el mesangio extraglo merular (EGM), y d e la mácula
densa (MD) se ven ; PO, podocitos. En el polo urin ario, el epitelio parietal (PE) s e transforma
en el túbulo proximal (P). (Microscopía d e luz; aumento x 390).
Figura 1-6 porción periférica de un lóbulo glomerular. Esta parte muestra un
capilar, la posición axial del mesangio, y el epitelio visceral (podocitos). En la
interfaz de capilar mesangial, el endotelio capilar se apoya directamente el
mesangio.
Figura 1-7 capilar glomerular. A, La capa de interdigitación procesos de podocitos y la membrana basal glomerular (GBM) no rodean completamente el capilar. En los ángulos mesangiales (flechas), tanto desviarse de un supuesto pericapilar y cubren el mesangio. Procesos celulares mesangiales contienen paquetes densos de microfilamentos (MF) interconectan la MBG y salvar la distancia entre los dos ángulos mesangiales. B, la barrera de filtración. La parte periférica de la pared capilar glomerular comprende el endotelio con poros abiertos (puntas de flecha), el GBM, y los procesos de pie interdigitales. El GBM muestra una lámina densa delimitado por la lámina rara interna y
externa. Los procesos de pie están separadas por hendiduras de filtración puenteados por diafragmas delgadas (flechas). (Transmisiónmicroscopíaelectrónica [MET]; ampliación: A, × 8770; B, × 50 440).

La carga electronegativa de la MBG se debe principalmente a la presencia de
proteoglicanos polianiónicas. Los principales proteoglicanos de la MBG son
proteoglicanos heparán sulfato, incluyendo perlecan y agrina. Moléculas de
proteoglicanos se agregan para formar una malla que se mantiene bien hidratado
por moléculas de agua atrapadas en los intersticios de la matriz. Abrahamson9
proporciona un resumen actualizado de la organización molecular de la MBG.
Mesangio
Tres tipos de células principales se producen dentro del mechón glomerular,
todos los cuales están en estrecho contacto con la MBG: las células
mesangiales, las células endoteliales, y podocitos. La proporción de células
mesangiales / endotelial / podocitos es 2: 3: 1 en la rata. Las células
mesangiales y la matriz mesangial establecen el mesangio glomerular. Además, algunos estudios sugieren que los antígenos macrófagos cojinete
de HLA-DR / Ia-similares también pueden rara vez se encuentran en e l
mesangio normal.
Células me sangialesson de forma irregular, con muchos procesos que se
extienden desde el cuerpo celular hacia el GBM (ver Figs. 1-6, 1 -7). En estos
procesos, los conjuntos densos de microfilamentos se encuentran, que contiene actina, miosina, y α-actinin.10 Los procesos están unidos
directamente o a través de la interposición de microfibrillas a la GBM (véase
la discusión más adelante). El GBM representa la estructura efector de la
contractilidad mesangial. Conexiones celular GBM mesangiales son
especialmente prominentes junto a los capilares, la interconexión de los dos
ángulos mesangiales opuestas de la MBG. Las células mesangiales poseen una gran variedad de receptores, incluyendo
los de la angiotensina II (Ang II), vasopresina, factor natriurético atrial,
prostaglandinas, transformando β del factor de crecimiento (TGF-beta) y
otros factores de crecimiento (PDGF, EGF, CTGF) 11.
La matriz mesangial llena los espacios altamente irregulares entre las
células mesangiales y la perimesangial GBM, el anclaje de las células mesangiales a la GBM.6 La organización ultraestructural de esta matriz se
entiende de manera incompleta. En las muestras preparadas por una
técnica que evita tetróxido de osmio y utiliza ácido tánico para la tinción, se
ve una densa red de microfibrillas elásticas. Muchas proteínas de la matriz
extracelular comunes se han demostrado dentro de la matriz mesangial,
incluyendo los tipos de colágeno IV, componentes V y VI y proteínas microfibrilar tales como la fibrilina y el 31-kilodalton glicoproteína asociada
a microfibrillas. La matriz también contiene varias glicoproteínas, más abundante fibronectina, así como varios tipos de proteoglicanos.
Endotelio
Células endoteliales glomerulares consisten en cuerpos celulares y
periféricamente situados, atenuados y hojas citoplasmáticos altamente
Fenestrada (ver Figs. 1-6 y 1-7). Poros endoteliales glomerulares carecen de
los diafragmas, que se presentan sólo en el endotelio de los afluentes
finales a la eferente arteriole.6 El redondos u ovalados poros tienen un diámetro de 50 a 100 nm. La membrana luminal de las células endoteliales
está cargado negativamente debido a la capa de células de varias
licoproteínaspolianiónicos, incluyendo Podocalyxin. Además, los poros
endoteliales están llenos con tapones de tamiz hecho principalmente de sialoglicoproteínas.
Visceral epitelio (podocitos)
El epitelio visceral de Bowman cápsula comprende células altamente
diferenciadas, los podocitos (Figura 1.8;. Véase también la figura 1-6.). En
el glomérulo en desarrollo, podocitos tienen una forma poligonal simple.
En ratas, la actividad mitótica de estas células se completa poco después
del nacimiento, junto con el cese de la formación de nuevo esbozos nefrona (primordios).
En los seres humanos, este punto ya se alcanza durante la vida prenatal.
Podocitos diferenciados son incapaces de replicar; Por lo tanto
degenerado podocitos no pueden ser reemplazados en el adulto. Todos
los esfuerzos de la última década para encontrar células progenitoras que
pueden migrar hacia el penacho y reemplazar podocitos perdidos han fracasado. En respuesta a un estímulo de crecimiento extrema, como por
el factor de crecimiento de fibroblastos básico 2 (FGF-2), podocitos pueden someterse a la división nuclear mitótica.
Sin embargo, las células son incapaces de completar la división celular por la
citocinesis, resultando en células binucleadas o multinucleadas.
Podocitos tienen un cuerpo celular voluminosa que flota en el espacio
urinario, separado de la MBG por un espacio subpodocyte . Los cuerpos
celulares dan lugar a procesos primarios largas que se extienden hacia y
colocarán a los capilares por las porciones más distales y por una amplia
gama de procesos de pie. Esta situación precaria de ser fijado a la GBM sólo
por los procesos de los resultados en una vulnerabilidad única de los podocitos: se pierde en forma de células viables en la orina. Esto es
aparentemente el principal mecanismo de cómo se pierden los podocitos
durante la vida, con la apoptosis (muerte celular) jugando ningún papel
relevante.
La característica estructural más específico de los podocitos es el patrón de
procesos de pie que cubren el aspecto exterior de los capilares glomerulares. Los procesos de pie de podocitos vecinos interdigitan
regularmente entre sí, dejando ranuras serpenteantes (hendiduras de
filtración) entre las células que están puenteados por una estructura
extracelular, el diafragma de hendidura (Fig 1-9;. Ver también las figuras 1-6
para 1-. 8). Los podocitos son células epiteliales polarizadas con un luminal y
un dominio de membrana de células basales; el dominio de la membrana de células basales se corresponde con las únicas placas de los procesos de pie
que están incrustados en la MBG. La frontera entre basal y membrana
luminal es el diafragma de hendidura.
La membrana luminal y la ranura están cubiertos por una capa superficial de
espesor que es rica en sialoglicoproteínas, incluyendo Podocalyxin y
podoendin, y es responsable de la carga superficial negativa alta de los podocitos. En comparación, la membrana abluminal (es decir, las plantas de
procesos podocitos) contiene proteínas transmembrana específicos que
conectan el citoesqueleto a la MBG. Dos sistemas son conocidos: dímeros
(1) α3β1 integrina interconectar el citoplasmática proteínas de adhesión
focal vinculina, paxilina y Talin con el α3, α4, y las cadenas de a5 de
colágeno tipo IV y laminina 521; y (2) β-α-dystroglycans interconectar el adaptador de proteínas citoplasmática utrofina con agrina y a5 laminina cadenas en la MBG.
Figura 1-8 capilares glo merulares en la rata. Urin aria lado d el cap ilar está cubierta por los
podocitos altamente ramificados. El sistema d e interdigitación d e los proc esos primarios (PP) y
procesos de pie ( FP) líneas de tod a la superficie de los mechon es, que también se extiende por
debajo d e los cuerpos c elulares. Los proc esos de pie d e las células vecin as interdigitan pero
perdonó a las hendiduras de filt ración en el medio. (Microscopíaelectrónica de b arrido ;
magnific ación × 2200).

En contraste con el cuerpo de la célula, que alberga un sistema retículo
endoplasmático y aparato de Golgi prominente y ha bien desarrollado
máquinas autophagic, los procesos celula res contienen sólo unos orgánulos.
A citoesqueleto sofisticado representa la forma compleja de las células. En
el cuerpo celular y los procesos primarios, los microtúbulos y los filamentos
intermedios (vimentina, desmina) dominan. Dentro de los procesos de pie,
microfilamentos formar haces en forma de U prominentes dispuestos en el
eje longitudinal de dos procesos sucesivos de pie en un patrón de
superposición. En el centro, las curvas de estos paquetes están vinculados a
los microtúbulos de los procesos primarios; periféricamente, las curvas
están vinculados a la MBG por integrinas y dystroglycans. α-actinina-4 y
sinaptopodina establecer la agrupación-podocitos específica del
microfilamentos.
Podocitos contienen una gran variedad de receptores de la superficie,
incluyendo los de guanosinamonofosfato cíclico (cGMP) de señalización,
estimulada por péptidos natriuréticos (ANP, BNP, CNP y), así como por el
óxido nítrico (NO); monofosfato de adenosina cíclico (cAMP) de
señalización estimulada por la prostaglandina E2 (PGE2), dopamina,
isoproterenol, hormona paratiroidea (PTH) y el péptido relacionado con PTH-; y de iones de calcio (Ca2 +) de señalización estimulada por
numerosos ligandos, incluyendo Ang II, acetilcolina, PGF2, arginina
vasopresina (AVP), trifosfato de adenosina (ATP), la endotelina, e
histamina) 0,17 Entre los receptores de potencial transitori o (TRP) canales
de cationes, TRPC5 y TRPC6 han recibido recientemente mucha
attention.18-20 El objetivo principal de esta orquesta de señalización es el citoesqueleto, aunque los efectos concretos son poco conocidos. Otros
receptores, tales como por C3b, TGF-β, FGF-2, y otras citoquinas /
quimioquinas, han demostrado estar involucrados en el desarrollo de
podocitos diseases.17 Megalin es un receptor multiligandoendocitótica y el
principal antígeno de Heymann en la nefritis la rata, 21 pero no está
presente en los seres humanos.
Las hendiduras de filtración son los sitios de flujo de fluido convectivo a
través del epitelio visceral (véanse las Figs. 1-7 y 1-9). Hendiduras de
filtración tienen una anchura constante de aproximadamente 30 a 40 nm y
están puenteados por el diafragma de hendidura, una membrana proteínica
con una composición molecular incompletamente determinada.
Químicamente fija y el tejido tratado con ácido tánico revela una estructura similar a la cremallera con una fila de poros de aproximadamente 14 NM2 a
cada lado de una barra central. Actualmente, las proteínas conocidas para
establecer la membrana hendidura o mediar en su conexión con el
citoesqueleto de actina de los procesos de pie incluyen nefrina,
P-cadherina, FAT1, NEPH 1-3 y podocin.17 Sin embargo, cómo estas
moléculas interactúan entre sí para establecer una membrana porosa de tamaño selectivo aún no se conoce. Además de su función de barrera, la
membrana de hendidura es una plataforma para la señalización a la cytoskeleton.
El epitelio parietal
El epitelio parietal de la cápsula de Bowman está formado por células
epiteliales escamosas que descansan sobre una membrana basal (ver Figs.
1-4 y 1-5 ). Las células planas están llenos de haces de filamentos de actina
corriendo en todas direcciones. En contraste con la MBG, la membrana
basal parietal comprende varias capas de proteoglicano-densos que, además de tipo IV, contiene colágeno de tipo XIV. El proteoglicano
predominante de la membrana basal parietal es un sulfato de condroitina
proteoglycan.23 observaciones recientes sugieren que un nicho de células
madre epiteliales glomerulares reside dentro del epitelio parietal en la
transición hacia el túbulo proximal, 24 pero la evidencia inequívoca es
deficiente.
Filtración Barrera
La filtración a través de la pared capilar glomerular se produce a lo largo
de una vía extracelular, incluyendo los poros endoteliales, GBM, y se
cortó diafragma (véanse las Figs. 1-7 y 1-9). Todos estos componentes
son bastante permeable para el agua; la alta permeabilidad de agua,
pequeños solutos e iones resultados porque no hay membranas celulares se interponen.
Figura 1-9 barrera de filtración glomerular. Se muestran dos procesos de pie podocitos puenteados por la membrana de hendidura, la MBG, y el endotelio capilar poroso. Las superficies de los podocitos y del endotelio están cubiertos por una glycocalyx cargado negativamente que contiene el Podocalyxinsialoproteína (PC). El GBM se compone principalmente de colágeno de tipo IV (α3, α4 y α5), laminina (11 cadenas α5, ß2, y gamma 1), y el heparán
sulfato proteoglicano agrina. La membrana de hendidura representa una membrana porosa proteico compuesto por (en lo que se conoce) nefrina, NEPH 1-3 P-cadherina, y FAT1. El citoesqueleto de actina basada de los procesos de pie se conecta tanto a la GBM y la membrana de hendidura. En cuanto a las conexiones a la MBG, β1α3 integrina dímeros interconectar específicamente al complejo TPV (talina, paxilina, vinculina) a la laminina 11; la β- y α-dystroglycans interconectan utrofina a Agrin. Las proteínas de la membrana de hendidura se unen al citoesqueleto por varias proteínas adaptadoras, incluyendo podocina, zonulaoccludens proteína 1 (ZO-1; Z), proteína asociada a CD2 (CD), y cateninas (CAT). Entre los canales de cationes no selectivos (NSCC), asociados con TRPC6 podocina (y nefrina, no mostrados) en la membrana de hendidura. Sólo la angiotensina II (Ang II) de tipo 1 receptor (AT1) se muestra como un ejemplo de los muchos receptores de la superficie. Cas, p130Cas; Ez, ezrin; FAK, quinasa de adhesión focal; La ILK, integrina-quinasa vinculadas; M, la miosina; N, Na + -H + factor regulador intercambiador (NHERF2); S, sinaptopodina. (Modificado de referencia 41.)

La conductancia hidráulica de las capas individuales de la barrera de filtración
es difícil de estudiar. En un modelo matemático de la filtración glomerular, la
resistencia hidráulica del endotelio se predijo a ser pequeña, mientras que el
GBM y las hendiduras de filtración contribuyen cada uno alrededor de un
medio a la resistencia hidráulica total de la pared capilar.
La función de barrera de la pared capilar glomerular de macromoléculas es
selectivo para el tamaño, forma y carga. La selectividad de carga de la barrera
densa resulta de la acumulación de moléculas cargadas negativamente a lo
largo de toda la profundidad de la barrera de filtración, incluyendo la capa de
superficie de las células endoteliales, y desde el alto contenido de
proteoglicanos heparán sulfato cargados negativamente en el GBM.
Polianiónicos macromoléculas, tales como proteínas plasmáticas, son
repelidos por el escudo electronegativo procedente de estos densos
conjuntos de cargas negativas.
La estructura fundamental que representa la selectividad tamaño de la
barrera de filtración parece ser el diafragma de hendidura. Macromoléculas
no cargadas hasta un radio efectivo de 1,8 nm pase libremente a través del
filtro. Componentes más grandes son cada vez más restringida (indicados por
sus holguras fraccionarias, que disminuyen progresivamente) y están
totalmente restringidos en radios efectiva de más de 4 nm. Albúmina
plasmática tiene un radio efectivo de 3.6 nm; sin la repulsión de la carga
negativa, la albúmina plasmática pasaría a través del filtro en cantidades
considerables. Tan recientemente propuesto, un campo eléctrico (potencial
de flujo) se puede generar por filtración a través de la pared capilar
glomerular, que a su vez puede impedir el paso de las proteínas del plasma
cargados negativamente través de la barrera por electroforesis.
Estabilidad de Penacho glomerular
El principal desafío para los capilares glomerulares es combinar
permeabilidad selectiva con la estabilidad. Las paredes de los capilares
están constantemente expuestos a altos gradientes de presión transmural
de la alta presión de perfusión de los capilares glomerulares.
El sistema importante para el mantenimiento de la estructura compleja de los mechones consiste de la MBG y el mesangio. De hecho, los cilindros de
la MBG definen en gran medida la forma de los capilares glomerulares.
Estos cilindros no rodean completamente el tubo capilar, sin embargo, y
están abiertos hacia el mesangio. Mecánicamente, los cilindros se
completan con los procesos celulares mesangiales contráctiles que unen las
lagunas de la MBG entre dos opuestos ángulos mesangiales, permitiendo estas dos estructuras en conjunto para desarrollar tensión de la pared.
Tradicionalmente, los podocitos se han interpretado como un tipo de
pericitos que contribuye al desarrollo de la tensión de la pared mediante la
variación del tono de su sistema contráctil. En un desafío reciente a este
punto de vista, sin embargo, el sistema sólo queda capaz de crear tensión
en la pared se compone de los cilindros abiertos del GBM puenteado por las células mesangiales.
Además de la necesidad de desarrollar tensión de la pared para evitar la
dilatación de los capilares glomerulares, el patrón de plegamiento de la
MBG (es decir, la disposición de capilares glomerulares) de be ser
estabilizado contra los gradientes de presión centrífugas también. Esto
ocurre mediante la interconexión de los puntos de inflexión de la MBG por las células mesangiales de dentro y por podocitos de fuera.
túbulo renal
El túbulo renal se subdivide en varios segmentos distintos: un túbulo
proximal, un túbulo intermedia, un túbulo distal, un túbulo de conexión
(CNT), y el conducto colector (véase la figura 1-1 y la Tabla 1-1.). El asa de
Henle comprende la parte recta del túbulo proximal (que representa gruesa
rama descendente), el descendente delgada y las extremidades ascendente delgada (que representan el túbulo intermedio), y la rama gruesa ascendente
(que representa porción recta del túbulo distal), que incluye la mácula densa.
Los colectores segmentos de conducto de la CNT y varios forman el sistema de conductos colectores.
Los túbulos renales se describen por un epitelio de una sola capa anclado a
una membrana basal. El epitelio es un epitelio de transporte que consiste en
células epiteliales planas o cúbicas conectadas apical por un complejo de
unión que consiste en una unión estrecha (occludenszonula), una unión
adherens, y rara vez un desmosoma. Como resultado de esta organización,
existen dos vías diferentes a través del epitelio (Fig de 1-10.): Una vía
transcelular, incluido el transporte a través de las células luminales y
basolateral y las membranas a través del citoplasma, y una vía paracelular a
través de la unión compleja y lateral espacios intercelulares. Las
características funcionales de transporte paracelular se determinan por la
unión estrecha, que difiere en gran medida en su elaboración en los distintos
segmentos tubulares. Transporte transcelular se determina por los canales
específicos, transportistas y transportadores incluidos en las membranas de
la célula apical y basolateral. Los diversos segmentos de la nefrona difieren
en función de la distribución de las proteínas de transporte, y la capacidad de
respuesta a hormonas y medicamentos tales como diuréticos.
Tubulo Proximal
El túbulo proximal reabsorbe la mayor parte de agua y solutos filtrada (Fig.
1-11 A). El epitelio muestra numerosas adaptaciones estructurales para
este papel. El túbulo proximal tiene un borde prominente cepillo
(aumentando el área de superficie de la célula luminal) y extensa
interdigitación por procesos celulares basolateral (aumentando el área de superficie celular basolateral). Este interdigitación de células lateral se
extiende hasta la unión estrecha con fugas, aumentando así la correa de
unión apretada en longitud y proporcionar un gran aumento de paso para
el transporte pasivo de iones. Los túbulos proximales tienen grandes
mitocondrias, prominentes íntimamente asociados con las membranas
celulares basolateral, donde el sodio-potasio (Na +, K +) - se encuentra trifosfatasa de adenosina (ATPasa); esta maquinaria domina el transporte
transcelular. El transportador luminal para Na + entrada específica para el
túbulo proximal es el ion de sodio-hidrógeno (Na + -H +) intercambiador. La
alta permeabilidad hidráulica de agua tiene sus raíces en abundante
presencia de la proteína de canal de agua acuaporina 1 (AQP1).
Un sistema lisosomal prominente se conoce como el "aparato endocitótica vacuolar apical" y es responsable de la reabsorción de macromoléculas
(polipéptidos y proteínas tales como albúmina) que han pasado a través del
filtro glomerular. El túbulo proximal generalmente se subdivide en tres
segmentos (conocidos como S1, S2, S3, o P1, P2, P3) que difieren
considerablemente en la organización celular y por lo tanto también en la
función.
Figura 1-10 epitelios tubulares. Transporte a través del epitelio puede seguir dos
rutas: transcelular, a través de luminal y las membranas basolateral, y paracelular,
a través de la unión estrecha y los espacios intercelulares.

Asa de Henle
El asa de Henle se compone de la parte recta del túbulo proximal, rama descendente delgada y (en bucles largos) rama ascendente delgada y
gruesa rama ascendente (figura 1-12;. Véase también la figura 2.1.). La
rama descendente delgada, como en el túbulo proximal, es altamente
permeable para el agua (canales son de AQP1), mientras que a partir
exactamente en el punto de inflexión, la rama ascendente delgada es
impermeable para el agua. Las funciones de transporte específicas de las extremidades delgadas de Henle que contribuyen a la generación del gradiente osmótico medular están bajo debate.
Figura 1-11 Túbulos de la corteza renal. A, túbulo contorneado proximal está equipado con un borde en cepillo y un aparato de vacuolar prominente en el citoplasma apical. El resto del citoplasma está ocupado por un laberinto basal que consiste en grandes mitocondrias asociados con las membranas celulares basolateral. B, túbulo contorneado distal también ha interdigitados membranas celulares basolateral íntimamente asociados con grandes mitocondrias. En contraste con el túbulo proximal, sin embargo, la superficie apical se amplifica solamente por algunos microvellosidades rechoncha. (TEM; A, × 1,530; B, × 1,830).
Figura 01.12 Túbulos en la médula. A, Sección transversal a través de la franja interior de la médula exterior muestra una extremidad delgada descendente de un bucle largo Henle (DL), los medulares gruesa ascendente extremidades de Henle (AL), y un conducto (CD) recoger con células principales (P) y las células intercalares (ICC, capilares peritubulares;. F, fibroblastos B, En la sección transversal médula interna, descendente delgada y extremidades ascendente (TL), un conducto (CD recoger) y vasa recta (VR) se ven (TEM.; Una, × 990; B, × 1120).

La rama ascendente gruesa de Henle es a menudo llamado el "segmento
diluyendo". Es impermeable al agua pero reabsorbe cloruro de sodio
considerable (NaCl, sal), resultando en la separación de la sal del agua. La
sal está atrapado en la médula, mientras que el agua se lleva a la corteza,
donde se puede devolver a la circulación sistémica. El transportador
específico para la entrada de Na + en este segmento es el luminalNa + -K + -2Cl- cotransportador, que es el objetivo de los diuréticos como la
furosemida. Las uniones estrechas de la rama ascendente gruesa tienen una
permeabilidad relativamente baja. Las células fuertemente interdigitan por
procesos celulares basolateral, asociados con grandes mitocondrias que
suministran la energía para el transporte transepitelial. Las células sintetizan
una proteína específica, la proteína de Tamm-Horsfall, y lo liberan en la luz tubular. Proteína de Tamm-Horsfall se piensa que es más tarde importante
para prevenir la formación de cálculos renales. En contraste con el túbulo
proximal, la membrana luminal es de sólo escasamente amplificada por
microvellosidades. Justo antes de la transición al túbulo contorneado distal,
la rama gruesa ascendente de Henle contiene la mácula densa, que se
adhiere a la matriz glomérulo (véase aparato yuxtaglomerular).
Distal túbulo contorneado
El epitelio es bastante altamente diferenciada, exhibiendo las más
extensas interdigitacionesbasolateral de las células y la mayor densidad
de mitocondrias en todas partes de la nefrona (ver Fig. 1-11, B). Apical,
las células están equipadas con numerosas microvellosidades. El
transportador específico Na + del túbulo contorneado distal es el luminalNa + -CL- cotransportador, que es el objetivo de los diuréticos
tiazídicos.
SISTEMA DE DUCTOS COLECTORES
El sistema de conductos colectores incluye la CNT y los conductos
colectores corticales y medulares (ver Fig. 1-2). Dos nefronas pueden
unirse a nivel de la CNT, formando una arcada que citológico es un CNT.
Dos tipos de línea de células del túbulo de conexión: la célula CNT, que
es específico para los CNT, y la célula intercalada (IC), que también se
produce más tarde en el conducto colector. Las células de la CNT son similares a los conductos colectores (CD) células en la organización
celular. Ambos tipos de células comparten la sensibilidad a la
vasopresina (véase la sección siguiente); la célula CNT, sin embargo, carece de sensibilidad a los mineralocorticoides.
Conductos colectores
Los conductos colectores pueden subdividirse en conductos corticales y medulares, y de los conductos medulares en exterior e interior; las
transiciones son graduales (véase Fig. 1-12). Al igual que con la CNT,
los conductos colectores están revestidos por dos tipos de células:
células de CD (células principales y células IC). Las células IC
disminución en el número como el conducto colector desciende en la
médula y están ausentes de los conductos colectores papilares.
Las células CD son células poligonales simples, que aumentan en
tamaño hacia la punta de la papila (Fig. 1-13 A). La superficie basal de estas células se caracteriza por invaginaciones de la membrana celular
basal (repliegues basales). Las uniones estrechas tienen gran
profundidad apicobasal, y la superficie celular apical tiene un
glycocalyx prominente. A lo largo de todo el conducto colector, estas
células contienen un sistema de traslado luminal de acuaporina 2 bajo
el control de la hormona antidiurética (ADH, vasopresina), proporcionando el potencial para cambiar la permeabilidad al agua de
los túbulos colectores de cero (o al menos de bajo) a permeable. Un
canal luminalamilorida sensible Na + está implicada en la capacidad de respuesta de conductos colectores corticales a la aldosterona.
Las porciones terminales de los conductos colectores en la médula interna
expresan el UTB1 transportador de urea, que en ADH de manera
dependiente de cuentas para el reciclaje de urea, un proceso que es
crucial en el urineconcentrating mechanism.30 El segundo tipo de célula,
la célula IC, está presente tanto en la CNT y el conducto colector (Fig. 1 -13
B). Hay al menos dos tipos, designados A y B, células intercaladas que se distinguen por estructural, inmunocitoquímica, y las características
funcionales. Tipo A las células se han definido como la expresión de H + -
ATPasa en su membrana luminal; estas células secretan IC protones. Las
células de tipo B expresan la H + -ATPasa en la membrana basolateral;
estas células IC secretan iones bicarbonato (HCO3-) y reabsorben
protones. Con estos diferentes tipos de células, los conductos colectores son los reguladores finales de equilibrio de líquidos y electrolitos, que
juegan papeles importantes en el manejo de Na +, Cl -, y K +, así como el
ácido y la base. La capacidad de respuesta de los conductos colectores a la
vasopresina permite a un organismo para vivir en condiciones áridas, lo
que le permite producir una orina concentrada y, si es necesario, una
orina diluida.
El aparato yuxtaglomerular comprende la mácula densa, el
mesangioextraglomerular, la porción terminal de la arteriola aferente con sus
células granulares productoras de renina (también a menudo denominado
células yuxtaglomerulares), y las porciones iniciales de la arteriola eferente
(véase Fig. 1-4). La mácula densa es una placa de células especializadas en la
pared de la rama ascendente gruesa de Henle en el lugar donde el miembro se une a la mesangioextraglomerular del glomérulo matri z (Fig 1-14 A;. Véase también la figura 1-5. ).
Figura 1-13 Recogida d e c élulas d e los conductos. A, c élula P rincipal (c élulas CD) de un
conducto colector medular. La membrana c elular apic al tiene cierta microvellosidad es
rechoncho cubierto po r una glucocáliz prominente; la membran a de c élulas basales forma
invagin aciones. Tenga en cu enta la profunda unión estrech a. B , célu las intercaladas, t ipo A.
Nota del c itoplasma oscuro (celd as oscuras) con muchas mitocondrias y micropl iegues apic ales;
la membrana b asal forma in vaginaciones. (TEM; A, × 8720; B, × 6970).
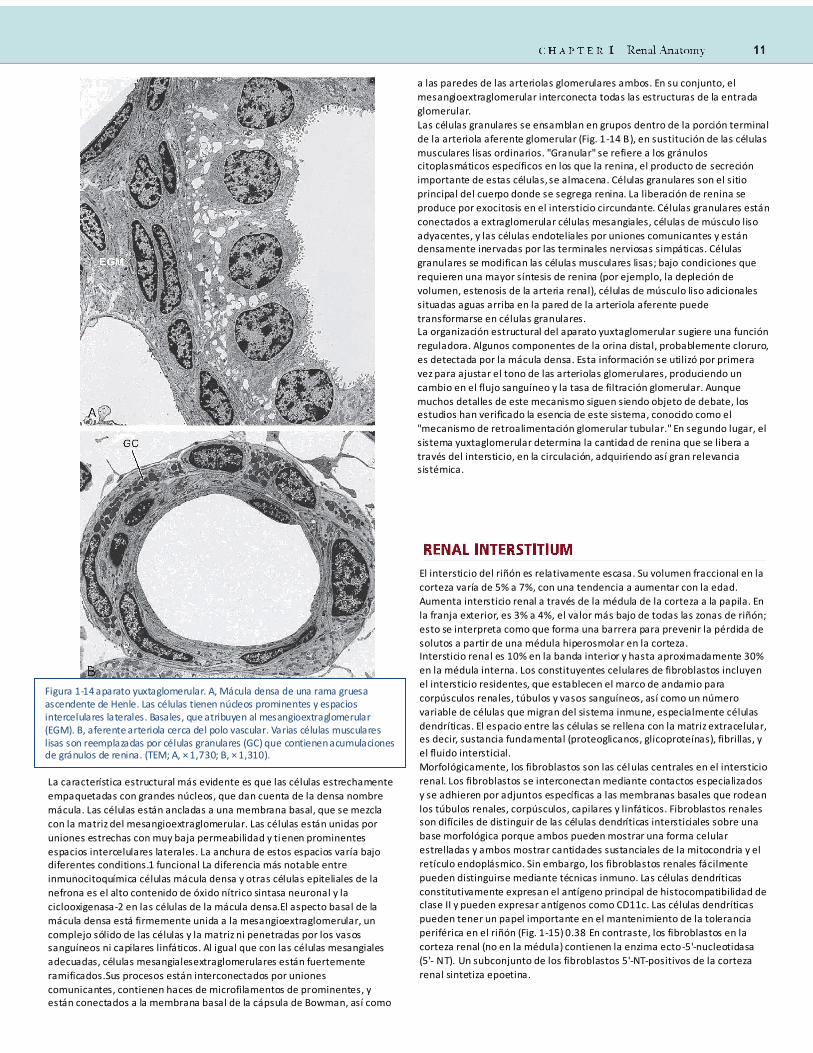
La característica estructural más evidente es que las células estrechamente
empaquetadas con grandes núcleos, que dan cuenta de la densa nombre
mácula. Las células están ancladas a una membrana basal, que se mezcla
con la matriz del mesangioextraglomerular. Las células están unidas por
uniones estrechas con muy baja permeabilidad y tienen prominentes
espacios intercelulares laterales. La anchura de estos espacios varía bajo diferentes conditions.1 funcional La diferencia más notable entre
inmunocitoquímica células mácula densa y otras células epiteliales de la
nefrona es el alto contenido de óxido nítrico sintasa neuronal y la
ciclooxigenasa-2 en las células de la mácula densa.El aspecto basal de la
mácula densa está firmemente unida a la mesangioextraglomerular, un
complejo sólido de las células y la matriz ni penetradas por los vasos sanguíneos ni capilares linfáticos. Al igual que con las células mesangiales
adecuadas, células mesangialesextraglomerulares están fuertemente
ramificados.Sus procesos están interconectados por uniones
comunicantes, contienen haces de microfilamentos de prominentes, y están conectados a la membrana basal de la cápsula de Bowman, así como
a las paredes de las arteriolas glomerulares ambos. En su conjunto, el
mesangioextraglomerular interconecta todas las estructuras de la entrada
glomerular.
Las células granulares se ensamblan en grupos dentro de la porción terminal
de la arteriola aferente glomerular (Fig. 1-14 B), en sustitución de las células
musculares lisas ordinarios. "Granular" se refiere a los gránulos citoplasmáticos específicos en los que la renina, el producto de secreción
importante de estas células, se almacena. Células granulares son el sitio
principal del cuerpo donde se segrega renina. La liberación de renina se
produce por exocitosis en el intersticio circundante. Células granulares están
conectados a extraglomerular células mesangiales, células de músculo liso
adyacentes, y las células endoteliales por uniones comunicantes y están densamente inervadas por las terminales nerviosas simpáticas. Células
granulares se modifican las células musculares lisas; bajo condiciones que
requieren una mayor síntesis de renina (por ejemplo, la depleción de
volumen, estenosis de la arteria renal), células de músculo liso adicionales
situadas aguas arriba en la pared de la arteriola aferente puede
transformarse en células granulares. La organización estructural del aparato yuxtaglomerular sugiere una función
reguladora. Algunos componentes de la orina distal, probablemente cloruro,
es detectada por la mácula densa. Esta información se utilizó por primera
vez para ajustar el tono de las arteriolas glomerulares, produciendo un
cambio en el flujo sanguíneo y la tasa de filtración glomerular. Aunque
muchos detalles de este mecanismo siguen siendo objeto de debate, los estudios han verificado la esencia de este sistema, conocido como el
"mecanismo de retroalimentación glomerular tubular." En segundo lugar, el
sistema yuxtaglomerular determina la cantidad de renina que se libera a
través del intersticio, en la circulación, adquiriendo así gran relevancia sistémica.
El intersticio del riñón es relativamente escasa. Su volumen fraccional en la
corteza varía de 5% a 7%, con una tendencia a aumentar con la edad.
Aumenta intersticio renal a través de la médula de la corteza a la papila. En
la franja exterior, es 3% a 4%, el valor más bajo de todas las zonas de riñón;
esto se interpreta como que forma una barrera para prevenir la pérdida de
solutos a partir de una médula hiperosmolar en la corteza. Intersticio renal es 10% en la banda interior y hasta aproximadamente 30%
en la médula interna. Los constituyentes celulares de fibroblastos incluyen
el intersticio residentes, que establecen el marco de andamio para
corpúsculos renales, túbulos y vasos sanguíneos, así como un número
variable de células que migran del sistema inmune, especialmente células
dendríticas. El espacio entre las células se rellena con la matriz extracelular, es decir, sustancia fundamental (proteoglicanos, glicoproteínas), fibrillas, y
el fluido intersticial.
Morfológicamente, los fibroblastos son las células centrales en el intersticio
renal. Los fibroblastos se interconectan mediante contactos especializados
y se adhieren por adjuntos específicas a las membranas basales que rodean
los túbulos renales, corpúsculos, capilares y linfáticos. Fibroblastos renales son difíciles de distinguir de las células dendríticas intersticiales sobre una
base morfológica porque ambos pueden mostrar una forma celular
estrelladas y ambos mostrar cantidades sustanciales de la mitocondria y el
retículo endoplásmico. Sin embargo, los fibroblastos renales fácilmente
pueden distinguirse mediante técnicas inmuno. Las células dendríticas
constitutivamente expresan el antígeno principal de histocompatibilidad de clase II y pueden expresar antígenos como CD11c. Las células dendríticas
pueden tener un papel importante en el mantenimiento de la tolerancia
periférica en el riñón (Fig. 1-15) 0.38 En contraste, los fibroblastos en la
corteza renal (no en la médula) contienen la enzima ecto-5'-nucleotidasa
(5'- NT). Un subconjunto de los fibroblastos 5'-NT-positivos de la corteza
renal sintetiza epoetina.
Figura 1-14 aparato yuxtaglomerular. A, Mácula densa de una rama gruesa ascendente de Henle. Las células tienen núcleos prominentes y espacios intercelulares laterales. Basales, que atribuyen al mesangioextraglomerular (EGM). B, aferente arteriola cerca del polo vascular. Varias células musculares lisas son reemplazadas por células granulares (GC) que contienen acumulaciones de gránulos de renina. (TEM; A, × 1,730; B, × 1,310).
En condiciones normales, es tos fibroblastos se encuentran
exclusivamente dentro de las porciones yuxtamedulares del laberinto
cortical . Cuando hay una demanda creciente de epoetina, las células
que sintetizan extienden a porciones más superficiales del laberinto
cortical y, en menor grado, a los rayos medulares. Los fibroblastos
dentro de la médula , en especial dentro de la médula interna, tienen
un fenotipo particular, conocidas como células intersticiales cargados
de l ípidos . Las células se orientan estrictamente perpendicularmente
hacia el eje longitudinal de los túbulos y vasos (funcionando todo en
paralelo) y contienen goti tas de l ípidos visibles. Estos fibroblastos de
la médula interna producen grandes cantidades de
glucosaminoglucanos y, posiblemente relacionados con las goti tas de l ípidos , lípidos vasoactivos, en particular PGE2.
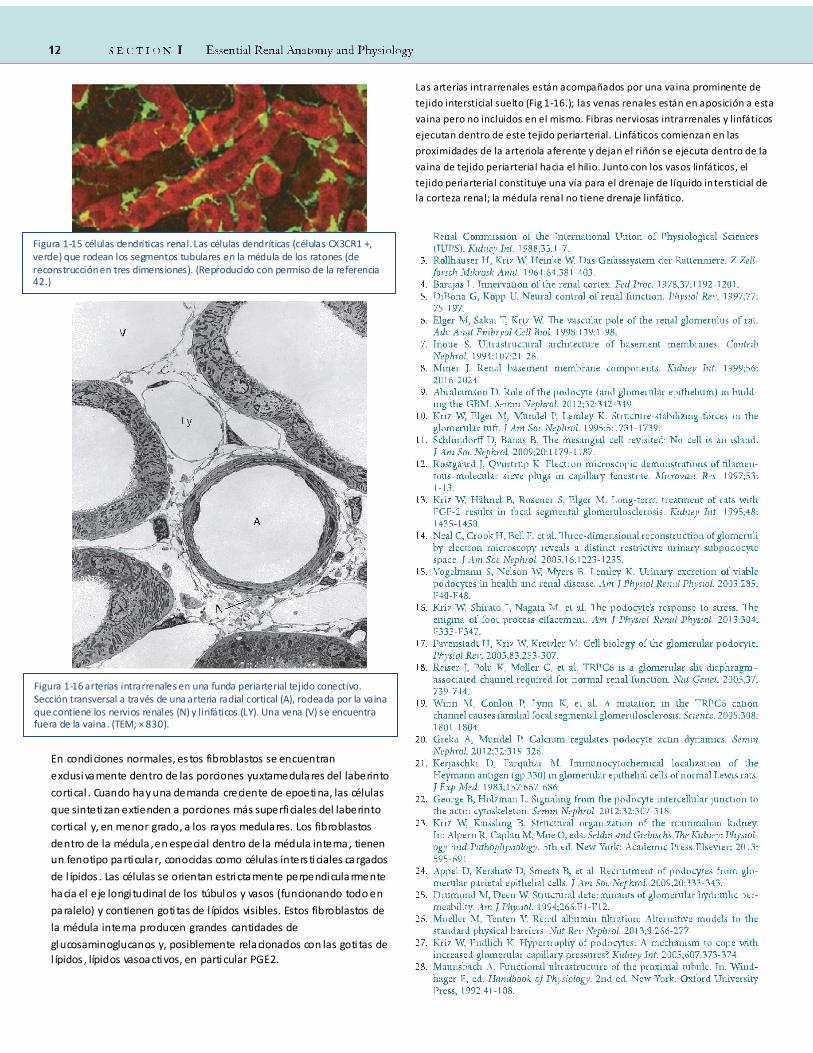
Las arterias intrarrenales están acompañados por una vaina prominente de
tejido intersticial suelto (Fig 1-16.); las venas renales están en aposición a esta
vaina pero no incluidos en el mismo. Fibras nerviosas intrarrenales y linfáticos
ejecutan dentro de este tejido periarterial. Linfáticos comienzan en las
proximidades de la arteriola aferente y dejan el riñón se ejecuta dentro de la
vaina de tejido periarterial hacia el hilio. Junto con los vasos linfáticos, el
tejido periarterial constituye una vía para el drenaje de líquido intersticial de
la corteza renal; la médula renal no tiene drenaje linfático.
Figura 1-15 células dendríticas renal. Las células dendríticas (células CX3CR1 +, verde) que rodean los segmentos tubulares en la médula de los ratones (de reconstrucción en tres dimensiones). (Reproducido con permiso de la referencia 42.)
Figura 1-16 arterias intrarrenales en una funda periarterial tejido conectivo. Sección transversal a través de una arteria radial cortical (A), rodeada por la vaina que contiene los nervios renales (N) y linfáticos (LY). Una vena (V) se encuentra fuera de la vaina. (TEM; × 830).